3.1. Gradient Design
The resulting morphology and relative concentration range and slopes of the two gradient designs were determined by optical microscopy.
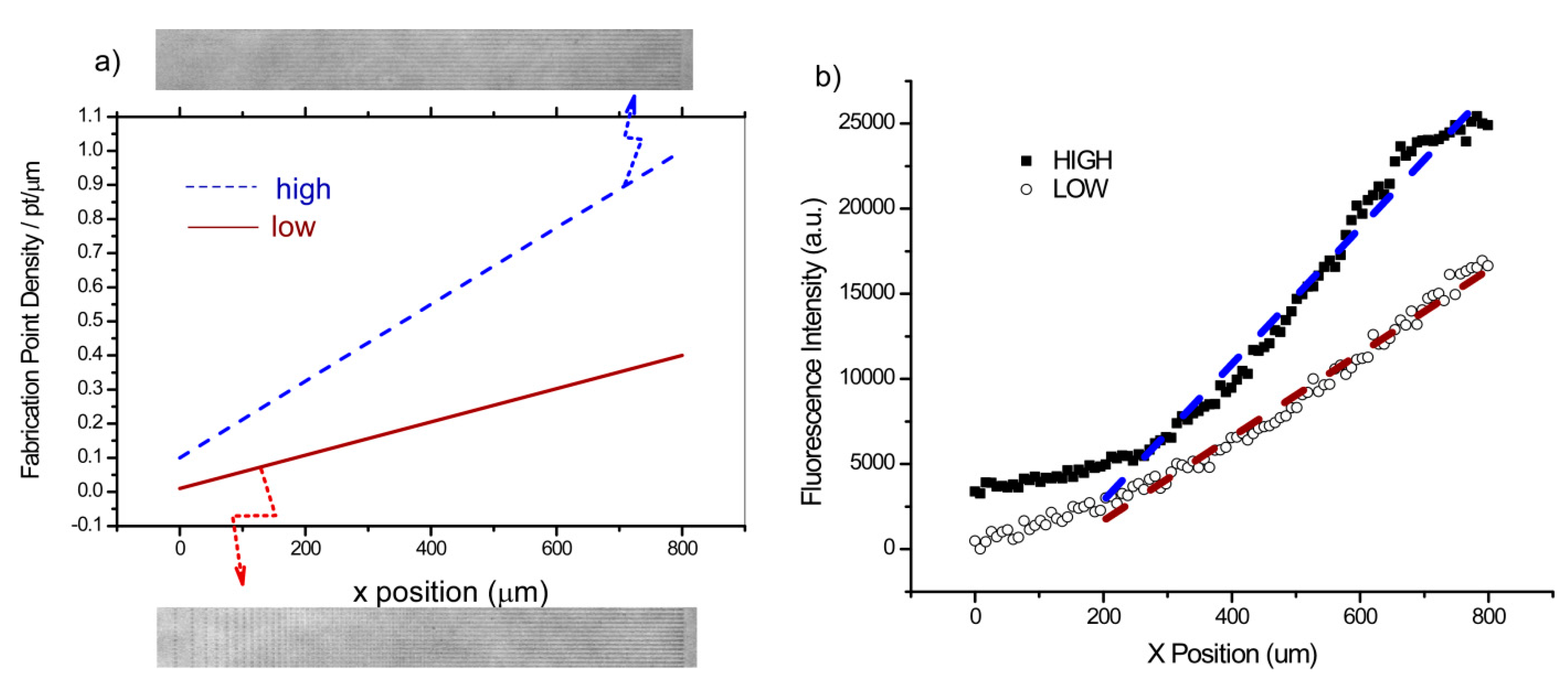
Figure 1a shows the design of the two gradients (low slope and high slope; see Methods for details) in terms of linear point density along the long axis. The representative respective phase contrast images of the gradients are show adjacent to the design. The relative concentration of the LN gradients was determined by immunofluorescence, where the structures were incubated with a primary LN antibody followed by a secondary fluorescent antibody. The fluorescence intensity and thus relative concentrations of the two designs are plotted in
Figure 1b with the best linear fit.
Figure 1.
Design of the low and high slope gradients: (a) Linear ramp showing the design as a function of point density, along with the resulting phase contrast image of the fabricated gradients. (b) The resulting LN immunofluorescence and corresponding linear fit (R2 > 0.95) representing the relative concentrations.
Figure 1.
Design of the low and high slope gradients: (a) Linear ramp showing the design as a function of point density, along with the resulting phase contrast image of the fabricated gradients. (b) The resulting LN immunofluorescence and corresponding linear fit (R2 > 0.95) representing the relative concentrations.
We ignored the first 200 microns of the gradient where the points are highly discrete and only used the 200–800 µm range. While there is scatter in the fluorescence intensities, the data in each case is well-fit by linear regression (
R2 > 0.95), and the change in slope between the low and high designs was approximately 1.5. The predicted ratio based on the design was 2.3, where the difference may have occurred through a variety of factors. However, it is likely that the upper end of the higher concentration design was in the limit which we have previously defined as the terminal crosslink density [
40], where more laser exposure does not result in higher concentration, as all the available reactive sites on the protein have already been utilized. The difference in concentration at the end of each gradient was also approximately a factor of 1.5.
3.2. Migration Speed and Directionality
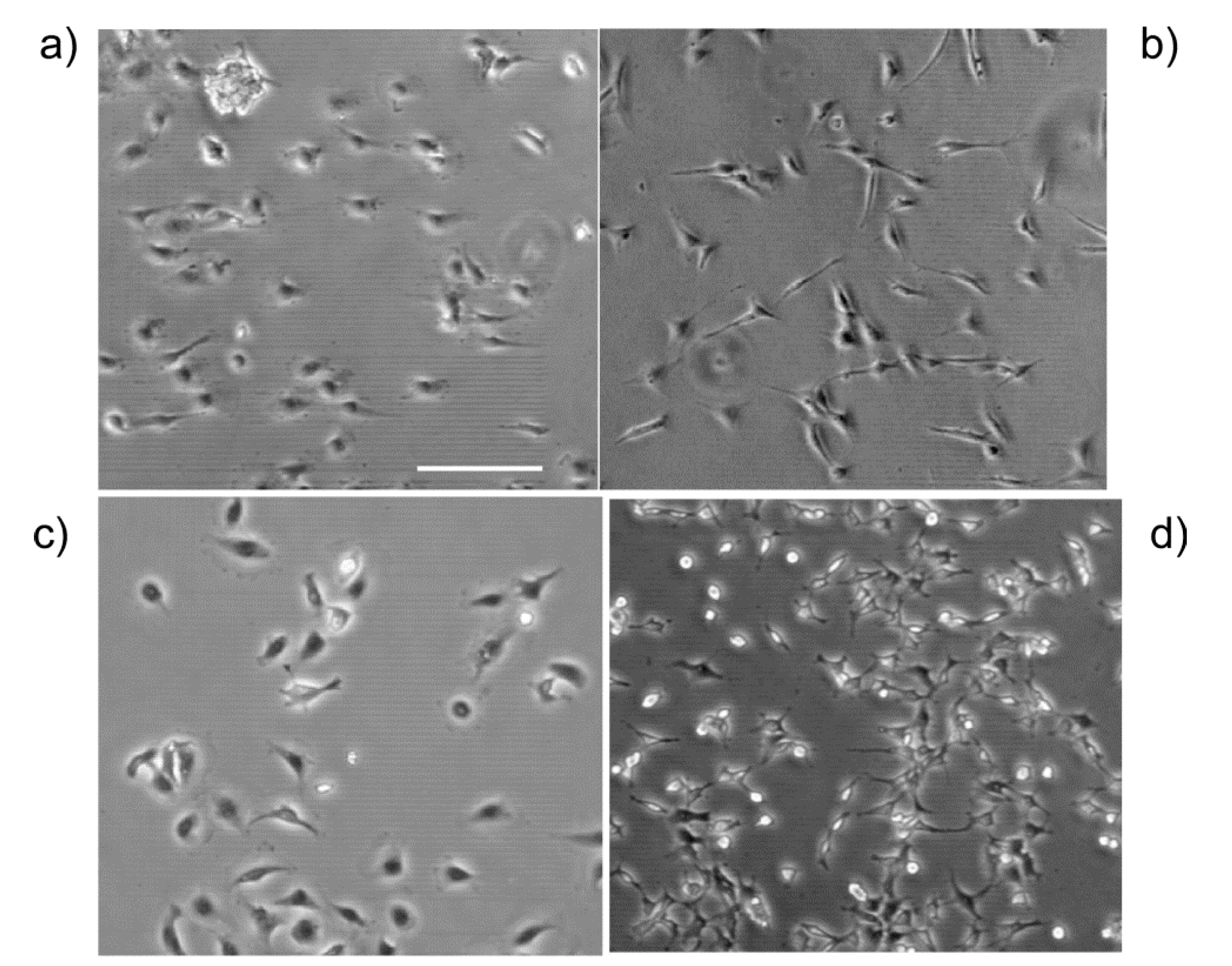
A representative phase contrast image of the IOSE, OVCA433, SKOV-3.ip1 and HEY-1 cells on LN gradients is shown in
Figure 2.
Figure 2.
Representative phase contrast images of each cell line on the high slope gradient, where (a) IOSE; (b) OVCA433; (c) SKOV-3.ip1; and (d) HEY-1. The gradient increases in concentration from left to right. Scale bar = 200 microns.
Figure 2.
Representative phase contrast images of each cell line on the high slope gradient, where (a) IOSE; (b) OVCA433; (c) SKOV-3.ip1; and (d) HEY-1. The gradient increases in concentration from left to right. Scale bar = 200 microns.
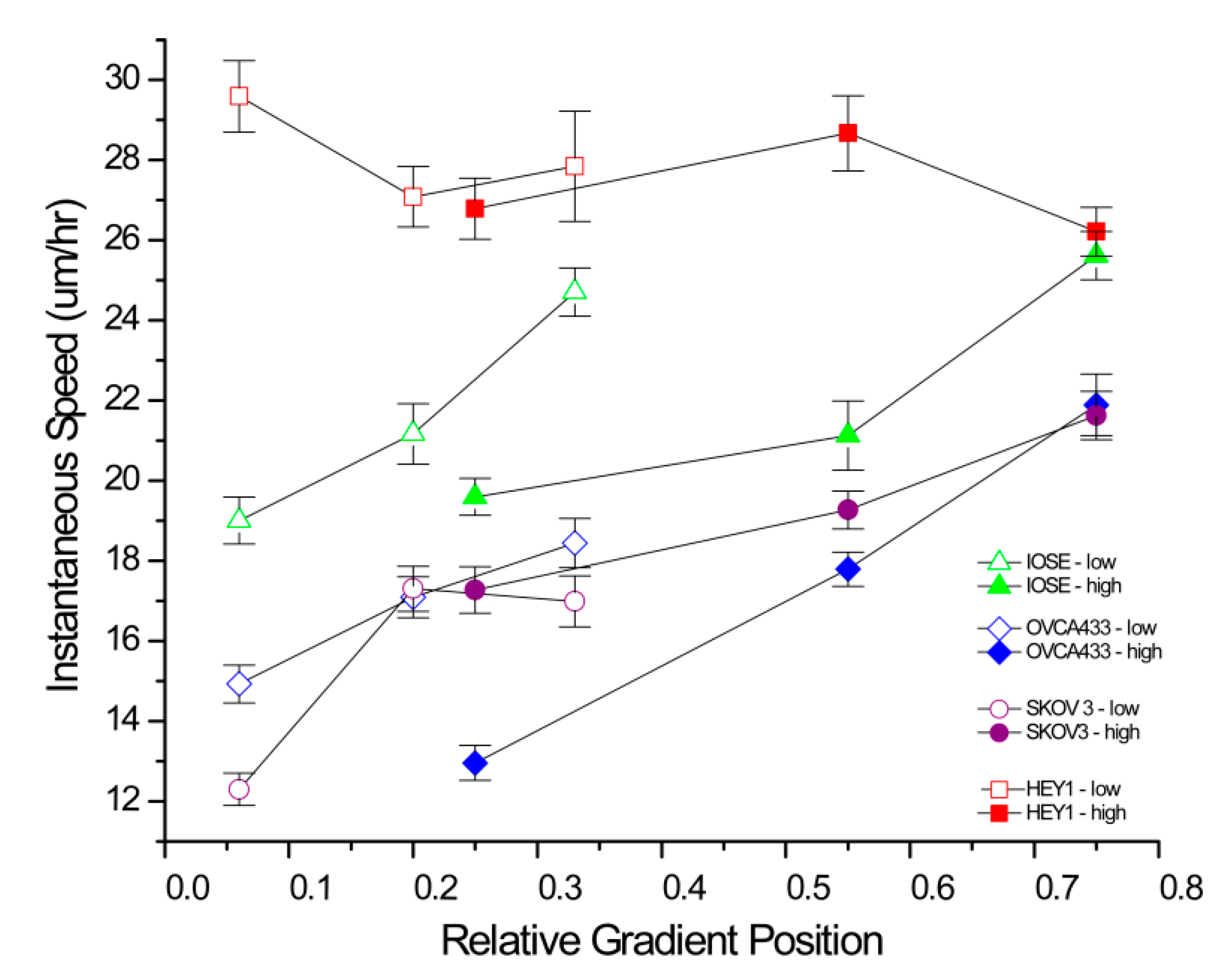
The averaged instantaneous speeds with the mean error are summarized in
Figure 3 for each cell type on the low and high gradient designs, where we break down each into three regions, which we delineate as low, medium and high concentration. We plotted the instantaneous speeds based on the relative position of the gradient where we defined the spread from 0 to 1. Specifically, cell measurements done on “mid” and “high” regions of the LOW slope gradients overlap in crosslinked laminin concentration (see
Figure 1) with “low” and “mid” regions of the HIGH slope gradients. For the three cancer cell lines (red = HEY-1, blue = OVCA433, and violet = SKOV-3.ip1), we found a smooth increase in speed with increasing metastatic potential for both the low and high gradient designs. This is similar as to what we found in our previous results for these cell types using constant concentration fibers of LN [
36]. We also find that the speed increases for the SKOV-3.ip1 and OVCA433 cells with increasing gradient concentration, demonstrating a strong haptotactic response to the changing LN concentration.
Most of the comparisons for each cell response at each concentration range were statistically different for these two cell types (
Table 1 and
Table 2).
Figure 3.
Merged migration speed data for the four cell lines on the low and high slope gradients. The statistical analysis for each cell line is given in
Table 1,
Table 2,
Table 3 and
Table 4. The data shows the relative roles of local concentration and slope on the haptotaxis response of the cell lines. Error bars represent standard error.
Figure 3.
Merged migration speed data for the four cell lines on the low and high slope gradients. The statistical analysis for each cell line is given in
Table 1,
Table 2,
Table 3 and
Table 4. The data shows the relative roles of local concentration and slope on the haptotaxis response of the cell lines. Error bars represent standard error.
Table 1.
p values from pair-wise t-tests of migration speeds of OVCA433 cells at low, medium, and high concentration regions of the low and high slope LN gradients. n = number of cells used in the analysis. In lieu of explicit p values, we denote “Y” (yes) as statistical differences corresponding to p < 0.05 to better visualize trends.
Table 1.
p values from pair-wise t-tests of migration speeds of OVCA433 cells at low, medium, and high concentration regions of the low and high slope LN gradients. n = number of cells used in the analysis. In lieu of explicit p values, we denote “Y” (yes) as statistical differences corresponding to p < 0.05 to better visualize trends.
| OVCA433 | Low Slope Low n = 24 | Low slope Medium n = 31 | Low slope High n = 24 | High slope Low n = 29 | High slope Medium n = 40 | High slope High n = 23 |
|---|
| Low slope Low | | Y | Y | Y | Y | Y |
| Low slope Medium | | | 0.09 | Y | 0.29 | Y |
| Low slope High | | | | Y | 0.38 | Y |
| High slope Low | | | | | Y | Y |
| Highslope Medium | | | | | | Y |
Table 2.
p values from pair-wise t-tests of migration speeds of SKOV-3.ip1 cells at low, medium, and high concentration regions of the low and high slope LN gradients. n = number of cells used in the analysis. “Y” corresponds to p < 0.05.
Table 2.
p values from pair-wise t-tests of migration speeds of SKOV-3.ip1 cells at low, medium, and high concentration regions of the low and high slope LN gradients. n = number of cells used in the analysis. “Y” corresponds to p < 0.05.
| SKOV-3.ip1 | Low Slope Low n = 12 | Low Slope Medium n = 19 | Low Slope High n = 15 | High Slope Low n = 11 | High Slope Medium n = 17 | High Slope High n = 17 |
|---|
| Low slope Low | | Y | Y | Y | Y | Y |
| Low slope Medium | | | 0.71 | 0.96 | Y | Y |
| Low slope High | | | | 0.74 | Y | Y |
| High slope Low | | | | | Y | Y |
| High slope Medium | | | | | | Y |
The IOSE cells (green) also displayed a strong migration speed response to the gradients, where the speed increased with increasing concentration, and also showed a strong dependence of the slope (statistics in
Table 3).
Table 3.
p values from pair-wise t-tests of migration speeds of IOSE cells at low, medium, and high concentration regions of the low and high slope LN gradients. n = number of cells used in the analysis. “Y” corresponds to p < 0.05.
Table 3.
p values from pair-wise t-tests of migration speeds of IOSE cells at low, medium, and high concentration regions of the low and high slope LN gradients. n = number of cells used in the analysis. “Y” corresponds to p < 0.05.
| IOSE | Low Slope Low n = 24 | Low Slope Medium n = 31 | Low Slope High n = 24 | High Slope Low n = 29 | High Slope Medium n = 40 | High Slope High n = 23 |
|---|
| Low slope Low | | Y | Y | 0.42 | 0.04 | Y |
| Low slope Medium | | | Y | 0.06 | 0.97 | Y |
| Low slope High | | | | Y | Y | 0.29 |
| High slope Low | | | | | 0.10 | Y |
| High slope Medium | | | | | | Y |
For example, the speed at the high concentration region of the low gradient was higher than the speeds at the low and medium regions of the high slope gradient. In our previous study of constant concentration LN fibers, the IOSE had slower migration speeds than the cancer cells [
36]. However, here these cells were faster than the OVCA433 and SKOV-3.ip1 cells. A possible explanation lies in the relative size of these cells. They have much larger spread area than the OVCA43 and SKOV-3.ip1 cells (~3500
vs. ~2000 µm
2) [
36] and experience a larger range of concentration, and the slope of the gradients may play a larger role in the observed behavior.
Similar to our previous results, the highly metastatic HEY-1 cells had the highest migration speed [
36]. In contrast to the other cells, the highly metastatic HEY-1 cells showed similar speeds at all concentration regions (see
Table 4).
Table 4.
p values from pair-wise t-tests of migration speeds of HEY-1 cells at low, medium, and high concentration regions of the low and high slope LN gradients. n = number of cells used in the analysis. “Y” corresponds to p < 0.05.
Table 4.
p values from pair-wise t-tests of migration speeds of HEY-1 cells at low, medium, and high concentration regions of the low and high slope LN gradients. n = number of cells used in the analysis. “Y” corresponds to p < 0.05.
| HEY-1 | Low Slope Low n = 21 | Low Slope Medium n = 25 | Low Slope High n = 30 | High Slope Low n = 25 | High Slope Medium n = 21 | High Slope High n = 26 |
|---|
| Low slope Low | | Y | 0.32 | Y | 0.48 | Y |
| Low slope Medium | | | 0.64 | 0.78 | 0.19 | 0.36 |
| Low slope High | | | | 0.52 | 0.66 | 0.26 |
| High slope Low | | | | | 0.12 | 0.5 |
| High slope Medium | | | | | | Y |
This independence of migration speed on LN concentration could suggest the migration for these cells is governed by contact guidance, rather than ECM binding cues. To investigate this idea, we performed analogous speed measurements on the same cell lines migrating on the analogous gradients created from BSA, which only provides contact guidance in the absence of ECM cues.
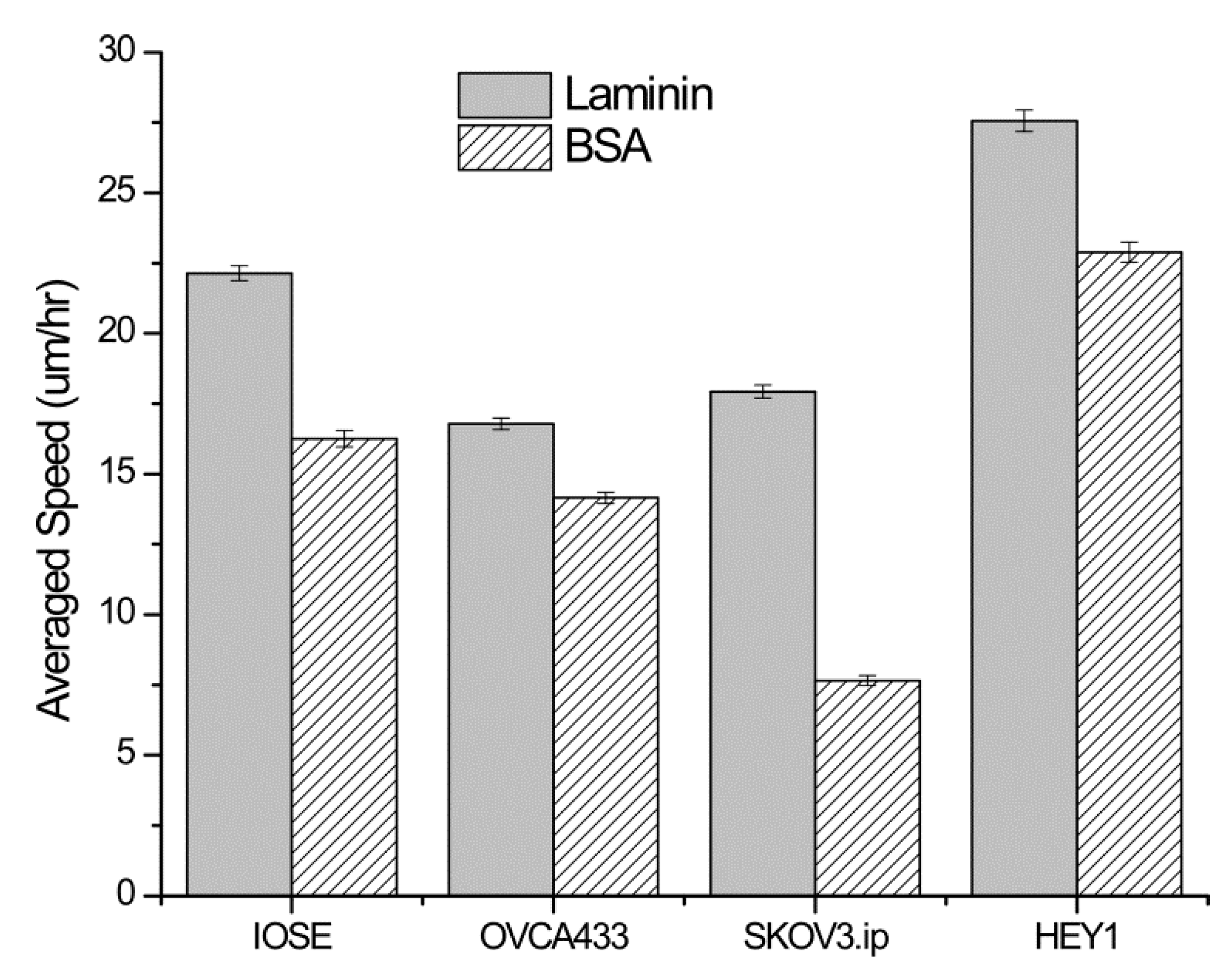
Figure 4 shows the comparison of the migration speeds from each cell line on the high slope LN and BSA gradients (high concentration region). In this analysis the speeds were not parsed by concentration regions but determined over the entire length of the laminin and BSA gradients. All the cell types migrated faster on the LN than the BSA gradient, where the differences were all significant (
p < 0.05). This is strong evidence that LN ECM cues (presumably through integrin binding) in conjunction with contact guidance contribute to the observed migration speed. We previously measured the migration of these cells on self-assembled monolayers and found random, slower migration than that observed on either LN or BSA constant concentration fibers, showing the ECM cues in conjunction with topographic cues result in faster migration than either alone [
36].
Figure 4.
Average migration speed of the four cell lines on LN and BSA gradients made from the same parameters for the high slope design. All cells migrated with statistically faster speeds on the LN, demonstrating the importance of both ECM and contact cues. Error bars represent standard error. Data was analyzed from 40–100 cells in each case.
Figure 4.
Average migration speed of the four cell lines on LN and BSA gradients made from the same parameters for the high slope design. All cells migrated with statistically faster speeds on the LN, demonstrating the importance of both ECM and contact cues. Error bars represent standard error. Data was analyzed from 40–100 cells in each case.
To further investigate the migration dynamics, we examined the directed migration by measuring the trajectories of each cell type relative to the gradient. The data for each line is shown as a polar plot in
Figure 5. The IOSE, OVCA433, and SKOV-3.ip1 display similar behavior, where the migration vector is predominantly along the gradient axis. The polar distribution in each case passes the Rayleigh test, showing a statistical directionality. The Watson
U2 tests between these cell types were not different. Together, this shows that the cells respond in a well-defined similar way to the gradient, and is consistent with the haptotactic response of each in
Figure 3. The data for the HEY-1 cells was markedly different. We observed more of a random distribution, which did not pass the Rayleigh test. Moreover, the Watson
U2 test was different relative to the other cells.
3.3. Cell Alignment on LN and BSA Gradients
We measured the resulting alignment of the four cell lines on LN gradients of the same low and high slope designs used for migration speeds. This was performed over the entire gradient as well as for three regions, which we again denote low, medium and high concentration, where these correspond to regions between 200–400 microns, 400–600 microns, and 600–800 microns, respectively. Images were selected at regular intervals over the 48 h observation time for this analysis. Cells were modeled as ellipses and the alignment was plotted as the angle between the long axis of the cell and the axis of the gradient [
35]. The numbers of cells analyzed for each range were between 50 and 100.
Figure 5.
Directed migration of the four cells lines along the fiber axis of the high slope gradient shown as polar plots. The IOSE, OVCA433, and SKOV-3.ip1 cells had similar responses of directed migration based on the Rayleigh test and the pairwise U2 Watson test. The HEY-1 cells did not pass the Rayleigh test, indicating a random distribution and were different from the other cells based the pairwise U2 Watson test. The slight asymmetries are a computational artifact.
Figure 5.
Directed migration of the four cells lines along the fiber axis of the high slope gradient shown as polar plots. The IOSE, OVCA433, and SKOV-3.ip1 cells had similar responses of directed migration based on the Rayleigh test and the pairwise U2 Watson test. The HEY-1 cells did not pass the Rayleigh test, indicating a random distribution and were different from the other cells based the pairwise U2 Watson test. The slight asymmetries are a computational artifact.
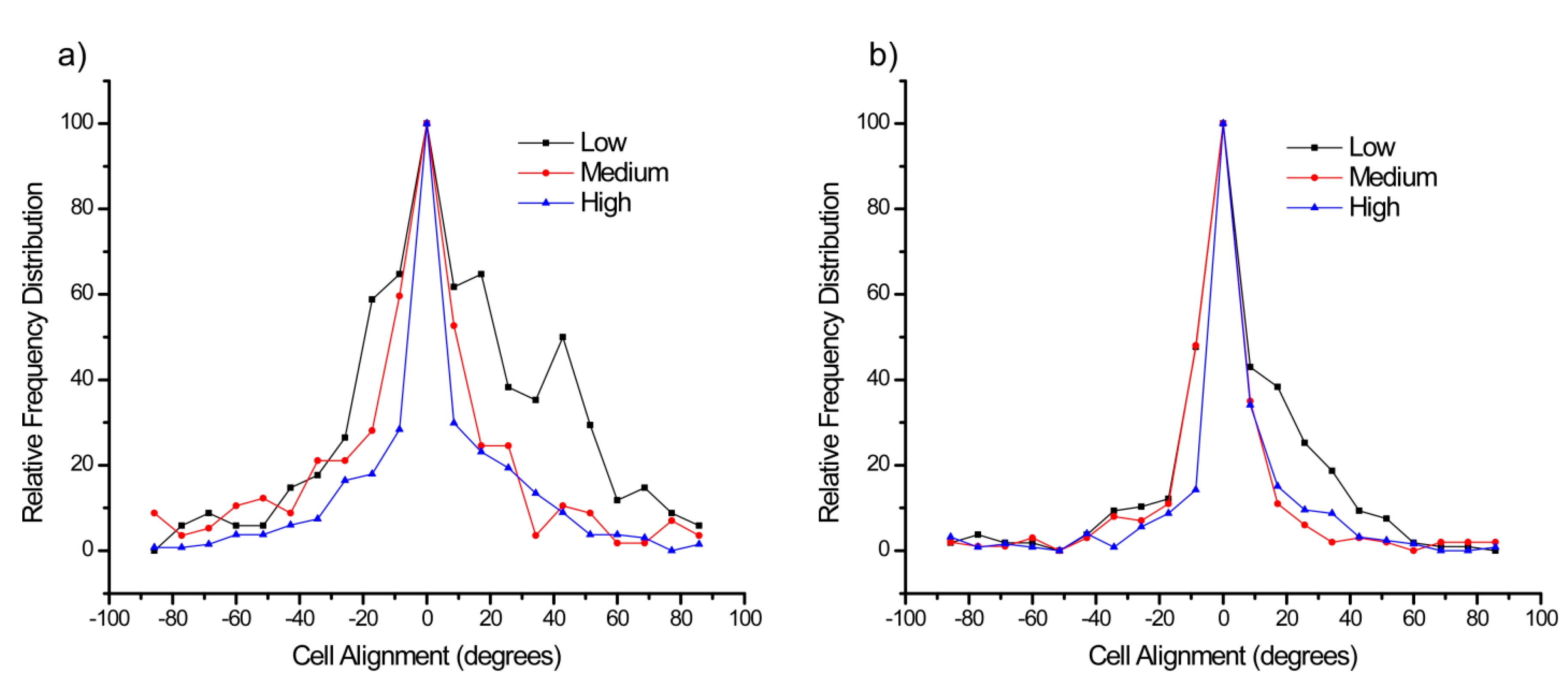
Figure 6a,b shows the distribution of angles of the OVCA433 cells for the lower and higher slope gradients over the three respective concentration regions. Here the data for each concentration was normalized to its own maximum to better observe the distribution. We found that for the lower gradient, each response was centered about 0 degrees,
i.e., parallel to the gradient direction, where cells on higher concentration regions had a tighter distribution in the orientation. This may arise due to the increased number of binding sites. In the higher slope gradient, the distribution continued to narrow, where the medium and high concentration regions had similar distributions. This is indicative of a saturated response, where increased binding sites did not affect the alignment. The distributions for the other cell types were also centered about 0 degrees, and in general decreased with increasing LN concentration. The respective widths of the distribution in terms of circular standard deviation are shown in the
Table 5. For all cell lines, using the Watson
U2 multisample test, the alignment for the regions on the low slope gradient was statistically broader than the medium and high concentration regions on the high slope gradient. This indicates that small changes in concentration have a fairly large effect on the alignment.
Figure 6.
Histograms of the alignment of OVCA433 cells on the low (a); and high (b) LN gradients relative to the fiber axis. The alignment is further broken down into low, medium, and high concentration regions of the respective gradient. The alignments were all peaked at 0°, i.e., parallel to the gradient but increased LN concentration resulted in narrower distributions until a saturated response was observed.
Figure 6.
Histograms of the alignment of OVCA433 cells on the low (a); and high (b) LN gradients relative to the fiber axis. The alignment is further broken down into low, medium, and high concentration regions of the respective gradient. The alignments were all peaked at 0°, i.e., parallel to the gradient but increased LN concentration resulted in narrower distributions until a saturated response was observed.
Table 5.
Circular standard deviations of the four cell lines on the low, medium, and high concentration regions of the low and high slope gradients. The distributions narrowed at higher concentrations for all cell lines.
Table 5.
Circular standard deviations of the four cell lines on the low, medium, and high concentration regions of the low and high slope gradients. The distributions narrowed at higher concentrations for all cell lines.
| LOW–Low | LOW–Med | LOW–High | HIGH–Low | HIGH–Med | HIGH–High |
|---|
| IOSE | 38.3° | 32.2° | 23.7° | 30.5° | 27.4° | 27.3° |
| OVCA433 | 32.2° | 30.0° | 24.3° | 23.1° | 21.2° | 20.9° |
| SKOV-3.ip1 | 35.0° | 32.6° | 22.6° | 30.5° | 27.8° | 25.9° |
| HEY-1 | 38.0° | 32.0° | 29.6° | 35.7° | 28.8° | 28.4° |
To compare the effect of contact guidance
vs. ECM cues, the alignment response on a BSA high slope gradient from the same design parameters as in LN was also measured. Representative phase contrast images of IOSE cells on LN and BSA gradients at the same time (12 h post-seeding) are shown in
Figure 7a. We found that the OVCA433 cells on LN were more spread and aligned at the same 12 h time point. This was also observed for the other cell types. The distribution of orientations for OVCA433 cells on the low, medium and high regions of the BSA gradient are shown in
Figure 7b. We observed a more random distribution of alignment angles for this cell type as well as the others on the BSA gradient, where, in all cases, the responses did not pass the Rayleigh test and had a random distribution of orientations. Thus we concluded that the LN gradient provides operative ECM cues resulting in cell alignment along the axis of the gradient.
Figure 7.
(a) Representative phase contrast images of OVCA433 cells on LN (left) and BSA (right) gradients 12 h after seeding. The cells are better spread on the LN gradient. Scale bar = 200 microns. (b) Polar plots of the orientation of the OVCA433 cells on the low, medium, and high slope ranges of the BSA gradient, showing a random distribution in each case. Polar plots are used for better visualization of the random distribution.
Figure 7.
(a) Representative phase contrast images of OVCA433 cells on LN (left) and BSA (right) gradients 12 h after seeding. The cells are better spread on the LN gradient. Scale bar = 200 microns. (b) Polar plots of the orientation of the OVCA433 cells on the low, medium, and high slope ranges of the BSA gradient, showing a random distribution in each case. Polar plots are used for better visualization of the random distribution.
3.4. Cytoskeletal Alignment
We measured the response of the cytoskeleton of the cell lines on the low slope gradient. Cells were fixed and stained for actin stress fibers and focal adhesion as described in the Methods. HEY-1 cells, as reported previously do not display distinct stress fibers or focal adhesions [
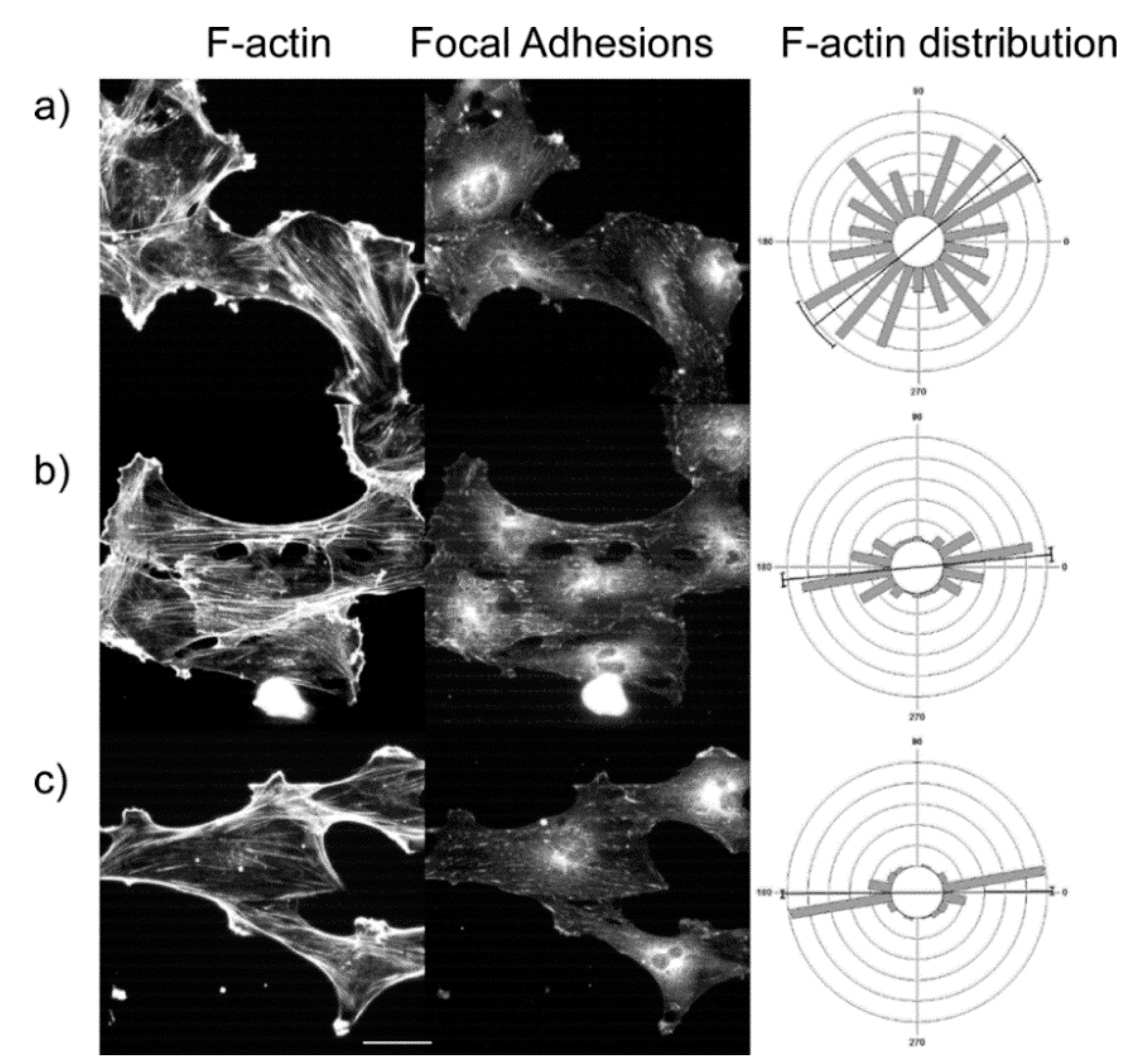
36] and are not included in this analysis. We determined the angular distribution of stress fibers relative to the gradient axis over low, medium, and high concentration ranges as was done in the previous section. Representative fluorescence images of SKOV-3.ip cells and the resulting angular distributions over multiple cells (300–500 measured stress fibers in each case) are shown in the left and middle columns, respectively, in
Figure 8. Here we grouped the actin orientation per region of the gradient rather than comparing the stress fibers within single cells and the statistical comparison was then performed between these groups.
The results for the three cell lines and three concentration ranges in terms of mean vector and circular standard deviation are summarized in
Table 6. We found that at low concentration, the distribution was essentially random and became highly aligned (
i.e., towards 0°) at medium and high concentration, although there is a distribution of angles around the mean. For each cell, the Pair-wise Watson
U2 tests showed that the distribution at each concentration was statistically different (
p < 0.001). These alignment results are also consistent with the ECM cues guiding the alignment of the f-actin cytoskeleton. This is further consistent with these cues guiding the cell orientation (
Section 3.3).
Figure 8.
Representative cytoskeleton images (left = phalloidin; middle = anti-vinculin) for SKOV-3.ip1 cells on the LN high slope gradient for (
a) low concentration; (
b) medium concentration; and (
c) high concentration regions. Scale bar = 40 microns. The right column is the f-actin angular distribution relative to the fiber axis, showing an increase in stress fiber alignment at higher LN concentration. The focal adhesion localization on the LN similarly increased. The statistical analysis for stress fibers and focal adhesions is given in
Table 6 and
Table 7, respectively.
Figure 8.
Representative cytoskeleton images (left = phalloidin; middle = anti-vinculin) for SKOV-3.ip1 cells on the LN high slope gradient for (
a) low concentration; (
b) medium concentration; and (
c) high concentration regions. Scale bar = 40 microns. The right column is the f-actin angular distribution relative to the fiber axis, showing an increase in stress fiber alignment at higher LN concentration. The focal adhesion localization on the LN similarly increased. The statistical analysis for stress fibers and focal adhesions is given in
Table 6 and
Table 7, respectively.
Table 6.
F-actin mean angle and distribution relative to the long axis for the cells lines on the low slope gradient. The average angle for three concentration regions of each cell type was statistically different. n = 300 to 500 stress fibers per region and the errors are standard deviations.
Table 6.
F-actin mean angle and distribution relative to the long axis for the cells lines on the low slope gradient. The average angle for three concentration regions of each cell type was statistically different. n = 300 to 500 stress fibers per region and the errors are standard deviations.
| Low | Medium | High |
|---|
| IOSE | 70.6 ± 67.6 | 6.0 ± 35.2 | 1.4 ± 26.7 |
| OVCA433 | 54.1 ± 55.8 | 3.9 ± 38.0 | 1.0 ± 23.9 |
| SKOV-3.ip1 | 55.7 ± 55.7 | 5.4 ± 25.6 | 0.6 ± 15.07 |
We also investigated the distribution of focal adhesions as a function of LN concentration. Representative anti-vinculin immunofluorescence images of the focal adhesions for SKOV-3.ip1 cells are shown in the right column of
Figure 8 next to the corresponding images of the actin stress fibers and angular distribution. For all concentrations, some punctuate focal adhesions are associated with the LN features. In analogy with the stress fiber analysis, these data points were grouped and analyzed based on regions along the gradient rather than by within individual cells. We measured the probability that focal adhesions were located on the LN gradients as opposed to spaces between the patterns. The data for IOSE, OVCA433, and SKOV-3 cells are tabulated in
Table 7, where a value of 1 corresponds to equal probability of the adhesions being on the LN and the BSA background between in linear gradient, and higher values are the increased probability of adhering to the gradient. The theoretical limits are then 0 and infinity for having none and all of the adhesions on the gradients, respectively. For the IOSE and SKOV-3.ip1 cells, the probability increased with concentration (
p < 0.01). The OVCA433 values were statistically similar but all statistically greater than unit probability.
Table 7.
Ratio of focal adhesion overlap on LN gradients vs. BSA background for the cells lines on the low slope gradient. The ratios for the IOSE and SKOV-3 cells were different at the three concentration regions. The OVCA433 data were not different from each other were all greater than unity. Focal adhesions from 16–20 cells are analyzed per region and the errors are mean error.
Table 7.
Ratio of focal adhesion overlap on LN gradients vs. BSA background for the cells lines on the low slope gradient. The ratios for the IOSE and SKOV-3 cells were different at the three concentration regions. The OVCA433 data were not different from each other were all greater than unity. Focal adhesions from 16–20 cells are analyzed per region and the errors are mean error.
| Low | Medium | High |
|---|
| IOSE | 0.94 ± 0.05 | 1.33 ± 0.08 | 1.54 ± 0.07 |
| OVCA433 | 1.10 ± 0.11 | 1.12 ± 0.09 | 1.25 ± 0.09 |
| SKOV-3.ip1 | 1.06 ± 0.06 | 1.40 ± 0.05 | 1.57 ± 0.06 |
While these preferred factors may seem low, the relative area covered by the LN gradient
vs. the BSA background is less than 10%, thus, based on area, the preference can be considered to be more pronounced. Moreover, it was not expected that the focal adhesions would be solely located on the LN as the BSA monolayer is not a repulsive background and the cells will eventually form new adhesions, especially as new matrix is formed. This is a different scenario than sometimes reported in the literature where islands or stripes of attractive regions were placed against a cell-repulsive background, and the focal adhesions are expressed solely on the fabricated regions by default [
41,
42].






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




