Next Generation Winemakers: Genetic Engineering in Saccharomyces cerevisiae for Trendy Challenges


{kind=link}
{kind=link}
{kind=link}
Abstract
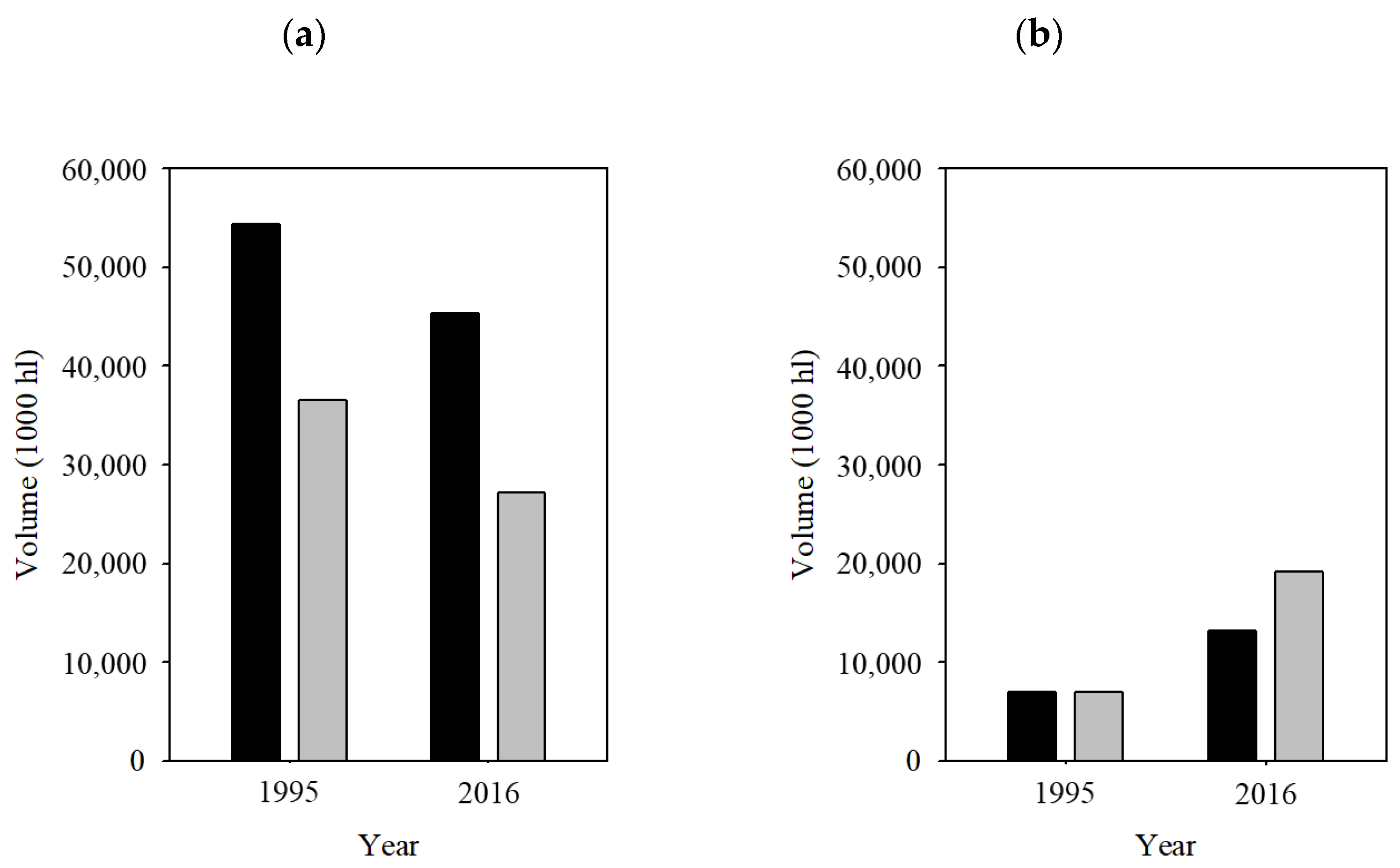
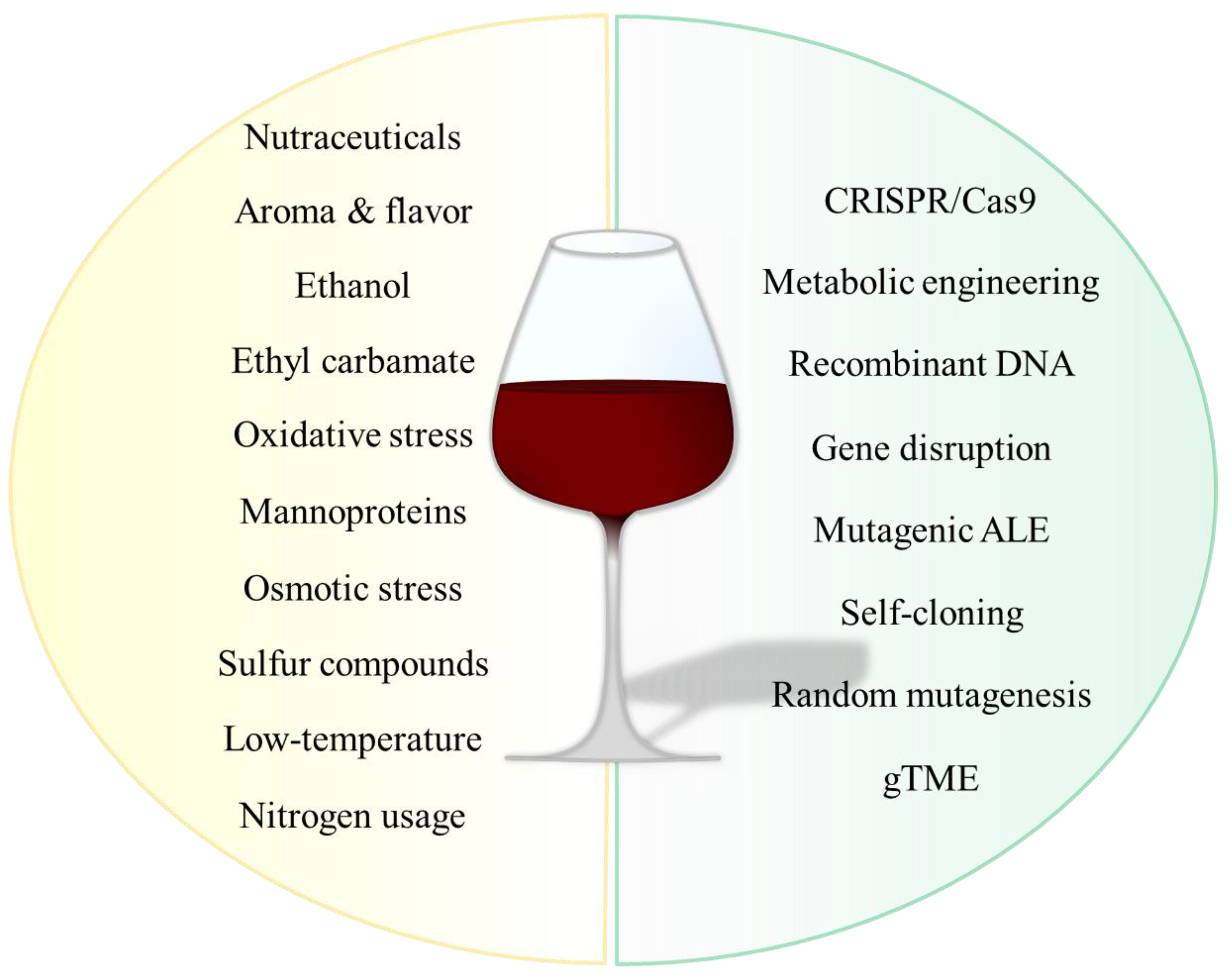
:1. Introduction
2. Nutraceuticals
2.1. Resveratrol
2.2. Hydroxytyrosol
3. Aroma and Flavor
4. Ethanol
4.1. Lower Ethanol Content
4.2. Higher Ethanol Content
5. Ethyl Carbamate
6. Other Oenological Traits
6.1. Oxidative Stress
6.2. Mannoproteins
6.3. Osmotic Stress
6.4. Sulfur Compounds
6.5. Low Temperature Fermentation
6.6. Nitrogen Usage
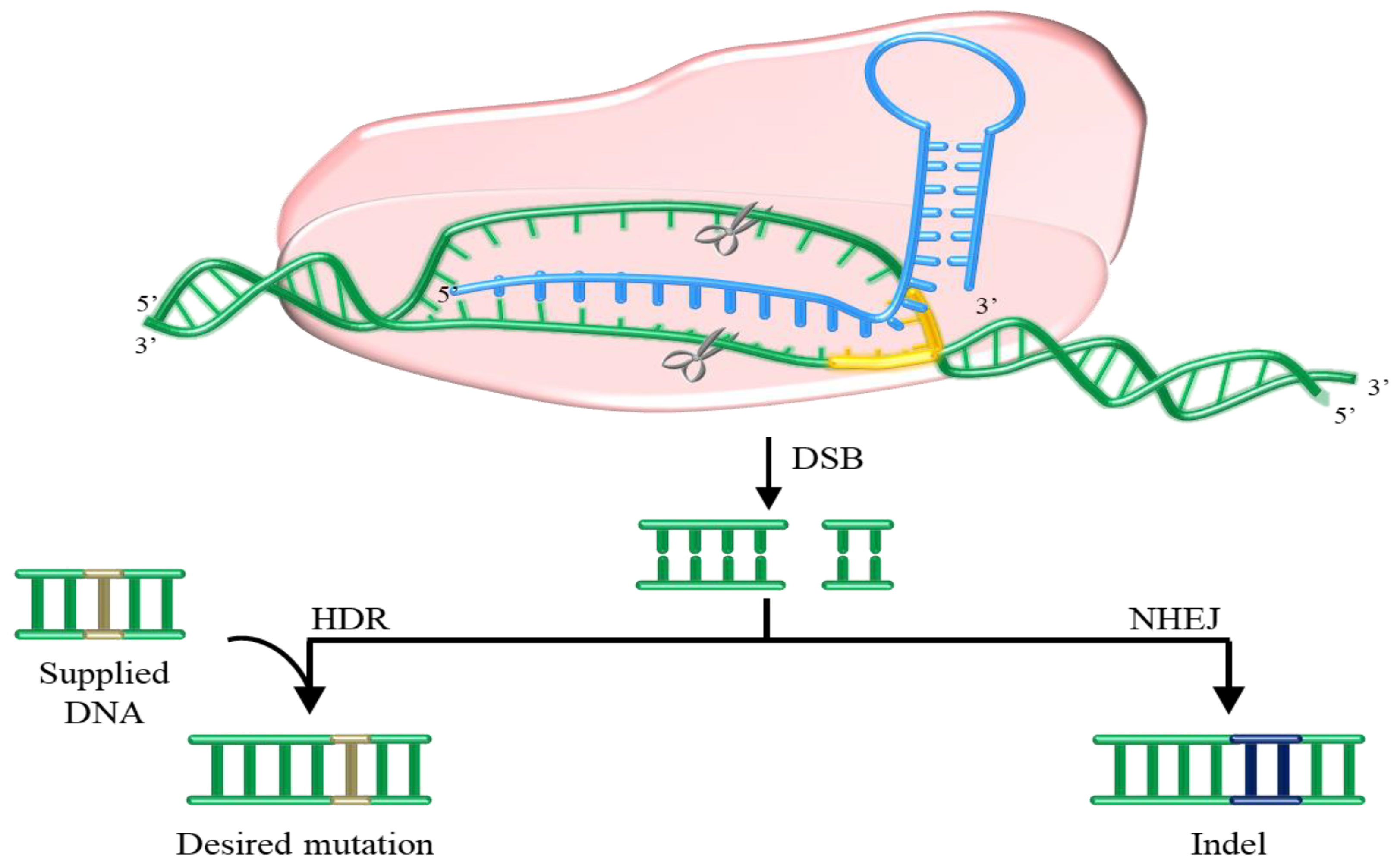
7. GMOs and Non-GMOs
8. Conclusions
Funding
Conflicts of Interest
References
- McGovern, P.; Jalabadze, M.; Batiuk, S.; Callahan, M.P.; Smith, K.E.; Hall, G.R.; Kvavadze, E.; Maghradze, D.; Rusishvili, N.; Bouby, L.; et al. Early Neolithic wine of Georgia in the South Caucasus. Proc. Nalt. Acad. Sci. USA 2017, 114, E10309–E10318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortimer, R.K. Evolution and variation of the yeast (Saccharomyces) genome. Genome Res. 2000, 10, 403–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martini, A. Origin and domestication of the wine yeast Saccharomyces cerevisiae. J. Wine Res. 1993, 4, 165–176. [Google Scholar] [CrossRef]
- Duan, S.F.; Han, P.J.; Wang, Q.M.; Liu, W.Q.; Shi, J.Y.; Li, K.; Zhang, X.L.; Bai, F.Y. The origin and adaptive evolution of domesticated populations of yeast from Far East Asia. Nat. Commun. 2018, 9, 2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, R.; Tronchoni, J.; Quirós, M.; Morales, P. Genetic improvement and genetically modified microorganisms. In Wine Safety, Consumer Preference, and Human Health; Moreno-Arribas, M.V., Bartolomé Sualdea, B., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 71–96. ISBN1 978-3-319-24512-6. ISBN2 (eBook) 978-3-319-24514-0. [Google Scholar] [CrossRef]
- Wortmann, J. Anwendung und Wirkung Reiner Hefen; Verlagsbuchhandlung Paul Parey: Berlin, Germany, 1895. [Google Scholar]
- Goffeau, A.; Barrell, B.G.; Bussey, H.; Davis, R.W.; Dujon, B.; Feldmann, H.; Galibert, F.; Hoheisel, J.D.; Jacq, C.; Johnston, M.; et al. Life with 6000 genes. Science 1996, 274, 546–567. [Google Scholar] [CrossRef] [Green Version]
- Liti, G.; Carter, D.M.; Moses, A.M.; Warringer, J.; Parts, L.; James, S.A.; Davey, R.P.; Roberts, I.N.; Burt, A.; Koufopanou, V.; et al. Population genomics of domestic and wild yeasts. Nature 2009, 458, 337–341. [Google Scholar] [CrossRef] [Green Version]
- Peter, J.; De Chiara, M.; Friedrich, A.; Yue, J.X.; Pflieger, D.; Bergström, A.; Sigwalt, A.; Barre, B.; Freel, K.; Llored, A.; et al. Genome evolution across 1011 Saccharomyces cerevisiae isolates. Nature 2018, 556, 339–344. [Google Scholar] [CrossRef] [Green Version]
- Baeshen, N.A.; Baeshen, M.N.; Sheikh, A.; Bora, R.S.; Ahmed, M.M.M.; Ramadan, H.A.I.; Saini, K.S.; Redwan, E.M. Cell factories for insulin production. Microb. Cell Fact. 2014, 13, 141. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Huang, M.; Nielsen, J. Exploring the potential of Saccharomyces cerevisiae for biopharmaceutical protein production. Curr. Opin. Biotechnol. 2017, 48, 77–84. [Google Scholar] [CrossRef]
- Hitzeman, R.A.; Chen, C.Y.; Hagie, F.E.; Lugovoy, J.M.; Singh, A. Yeast: An alternative organism for foreign protein production. In Recombinant DNA Products: Insulin, Interferon and Growth Hormone; Bollon, A.P., Ed.; CRC Press: Boca Raton, FL, USA, 2018; pp. 47–65. ISBN1 978-1-315-89715-8. ISBN2 978-1-351-07625-8. [Google Scholar] [CrossRef]
- Ko, J.K.; Lee, S.M. Advances in cellulosic conversion to fuels: Engineering yeasts for cellulosic bioethanol and biodiesel production. Curr. Opin. Biotechnol. 2018, 50, 72–80. [Google Scholar] [CrossRef]
- Favaro, L.; Jansen, T.; van Zyl, W.H. Exploring industrial and natural Saccharomyces cerevisiae strains for the bio-based economy from biomass: The case of bioethanol. Crit. Rev. Biotechnol. 2019, 39, 800–816. [Google Scholar] [CrossRef] [PubMed]
- Morawski, B.; Quan, S.; Arnold, F.H. Functional expression and stabilization of horseradish peroxidase by directed evolution in Saccharomyces cerevisiae. Biotechnol. Bioeng. 2001, 76, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Perez, D.; Garcia-Ruiz, E.; Alcalde, M. Saccharomyces cerevisiae in directed evolution: An efficient tool to improve enzymes. Bioengineered 2012, 3, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viña-Gonzalez, J.; Gonzalez-Perez, D.; Alcalde, M. Directed evolution method in Saccharomyces cerevisiae: Mutant Library creation and screening. J. Vis. Exp. 2016, 110, e53761. [Google Scholar] [CrossRef] [Green Version]
- Fraschini, R. Divide precisely and proliferate safely: Lessons from budding yeast. Front. Genet. 2019, 9, 738. [Google Scholar] [CrossRef]
- Kim, S.; Park, J.; Kim, T.; Lee, J.S. The functional study of human proteins using humanized yeast. J. Microbiol. 2020, 58, 343–349. [Google Scholar] [CrossRef]
- Pretorius, I.S.; Boeke, J.D. Yeast 2.0-connecting the dots in the construction of the world’s first functional synthetic eukaryotic genome. FEMS Yeast Res. 2018, 18, foy032. [Google Scholar] [CrossRef] [Green Version]
- Novo, M.; Bigey, F.; Beyne, E.; Galeote, V.; Gavory, F.; Mallet, S.; Cambon, B.; Legras, J.L.; Wincker, P.; Casaregola, S.; et al. Eukaryote-to-eukaryote gene transfer events revealed by the genome sequence of the wine yeast Saccharomyces cerevisiae EC1118. Proc. Natl. Acad. Sci. USA 2009, 106, 16333–16338. [Google Scholar] [CrossRef] [Green Version]
- Vilela, A. The importance of yeasts on fermentation quality and human health-promoting compounds. Fermentation 2019, 5, 46. [Google Scholar] [CrossRef] [Green Version]
- Stervbo, U.; Vang, O.; Bonnesen, C. A review of the content of the putative chemopreventive phytoalexin resveratrol in red wine. Food Chem. 2007, 101, 449–457. [Google Scholar] [CrossRef]
- Mukherjee, S.; Dudley, J.I.; Das, D.K. Dose-dependency of resveratrol in providing health benefits. Dose Response 2010, 8, 478–500. [Google Scholar] [CrossRef]
- Weiskirchen, S.; Weiskirchen, R. Resveratrol: How much wine do you have to drink to stay healthy? Adv. Nutr. 2016, 7, 706–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirola, L.; Frojdo, S. Resveratrol: One molecule, many targets. IUBMB Life 2008, 60, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Kuršvietienė, L.; Stanevičienė, I.; Mongirdienė, A.; Bernatonienė, J. Multiplicity of effects and health benefits of resveratrol. Medicina 2016, 52, 148–155. [Google Scholar] [CrossRef]
- Becker, J.V.W.; Armstrong, G.O.; van der Merwe, M.J.; Lambrechts, M.G.; Vivier, M.A.; Pretorius, I.S. Metabolic engineering of Saccharomyces cerevisiae for the synthesis of the wine-related antioxidant resveratrol. FEMS Yeast Res. 2003, 4, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.; Liang, J.L.; Kang, L.Z.; Huang, X.Y.; Huang, J.J.; Ye, Z.W.; Guo, L.Q.; Lin, J.F. Increased resveratrol production in wines using engineered wine strains Saccharomyces cerevisiae EC1118 and relaxed antibiotic or auxotrophic selection. Biotechnol. Prog. 2015, 31, 650–655. [Google Scholar] [CrossRef]
- Shin, S.Y.; Jung, S.M.; Kim, M.D.; Han, N.S.; Seo, J.H. Production of resveratrol from tyrosine metabolically engineered Saccharomyces cerevisiae. Enzyme Microb. Technol. 2012, 51, 211–216. [Google Scholar] [CrossRef]
- Rebollo-Romero, I.; Fernández-Cruz, E.; Carrasco-Galán, F.; Valero, E.; Cantos-Villar, E.; Cerezo, A.B.; Troncoso, A.M.; Carmen Garcia-Parrilla, M. Factors influencing the production of the antioxidant hydroxytyrosol during alcoholic fermentation: Yeast strain, initial tyrosine concentration and initial must. LWT-Food Sci. Technol. 2020, 130, 109631. [Google Scholar] [CrossRef]
- Muñiz-Calvo, S.; Bisquert, R.; Puig, S.; Guillamón, J.M. Overproduction of hydroxytyrosol in Saccharomyces cerevisiae by heterologous overexpression of the Escherichia coli 4-hydroxyphenylacetate 3-monooxygenase. Food Chem. 2020, 308, 125646. [Google Scholar] [CrossRef]
- Romboli, Y.; Mangani, S.; Buscioni, G.; Granchi, L.; Vincenzini, M. Effect of Saccharomyces cerevisiae and Candida zemplinina on quercetin, vitisin A and hydroxytyrosol contents in Sangiovese wines. World J. Microbiol. Biotechnol. 2015, 31, 1137–1145. [Google Scholar] [CrossRef]
- Karković Marković, A.; Torić, J.; Barbarić, M.; Jakobušić Brala, C. Hydroxytyrosol, Tyrosol and derivatives and their potential effects on human health. Molecules 2019, 24, 2001. [Google Scholar] [CrossRef] [Green Version]
- Proestos, C.; Bakogiannis, A.; Psarianos, C.; Koutinas, A.A.; Kanellaki, M.; Komaitis, M. High performance liquid chromatography analysis of phenolic substances in Greek wines. Food Control. 2005, 16, 319–323. [Google Scholar] [CrossRef]
- Minuti, L.; Pellegrino, R.M.; Tesei, I. Simple extraction method and gas chromatography-Mass spectrometry in the selective ion monitoring mode for the determination of phenols in wine. J. Chromatogr. A 2006, 1114, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Mar, M.I.; Mateos, R.; García-Parrilla, M.C.; Puertas, B.; Cantos-Pinar, E. Bioactive compounds in wine: Resveratrol, hydroxytyrosol and melatonine: A review. Food Chem. 2012, 130, 797–813. [Google Scholar] [CrossRef]
- Li, X.; Chen, Z.; Wu, Y.; Yan, Y.; Sun, X.; Yuan, Q. Establishing an artificial pathway for efficient biosynthesis of hydroxytyrosol. ACS Synth. Biol. 2018, 7, 647–654. [Google Scholar] [CrossRef]
- Brouk, M.; Fishman, A. Protein engineering of toluene monooxygenases for synthesis of hydroxytyrosol. Food Chem. 2009, 116, 114–121. [Google Scholar] [CrossRef]
- Molina-Espeja, P.; Cañellas, M.; Plou, F.J.; Hofrichter, M.; Lucas, F.; Guallar, V.; Alcalde, M. Synthesis of 1-naphthol by a natural peroxygenase engineered by directed evolution. ChemBioChem 2016, 17, 341–349. [Google Scholar] [CrossRef] [Green Version]
- Boronat, A.; Mateus, J.; Soldevila-Domenech, N.; Guerra, M.; Rodríguez-Morató, J.; Varon, C.; Muñoz, D.; Barbosa, F.; Morales, J.C.; Gaedigk, A.; et al. Cardiovascular benefits of tyrosol and its endogenous conversion into hydroxytyrosol in humans. A randomized, controlled trial. Free Radic. Biol. Med. 2019, 143, 471–481. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, Y.; Peter Guengerich, F.; Ma, L.; Li, S.; Zhang, W. Engineering cytochrome P450 enzyme systems for biomedical and biotechnological applications. J. Biol. Chem. 2020, 295, 833–849. [Google Scholar] [CrossRef] [Green Version]
- Georgiev, V.; Ananga, A.; Tsolova, V. Dietary supplements/nutraceuticals made from grapes and wines. In Wine Safety, Consumer Preference, and Human Health; Moreno-Arribas, M.V., Bartolomé Sualdea, B., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 201–227. ISBN1 978-3-319-24512-6. ISBN2 (eBook) 978-3-319-24514-0. [Google Scholar] [CrossRef]
- Avbelj, M.; Zupan, J.; Kranjc, L.; Raspor, P. Quorum-Sensing Kinetics in Saccharomyces cerevisiae: A Symphony of ARO Genes and Aromatic Alcohols. J. Agric. Food Chem. 2015, 63, 8544–8550. [Google Scholar] [CrossRef]
- Avbelj, M.; Zupan, J.; Raspor, P. Quorum-sensing in yeast and its potential in wine making. Appl. Microbiol. Biotechnol. 2016, 100, 7841–7852. [Google Scholar] [CrossRef] [PubMed]
- Herrero, O.; Ramón, D.; Orejas, M. Engineering the Saccharomyces cerevisiae isoprenoid pathway for de novo production of aromatic monoterpenes in wine. Metab. Eng. 2008, 10, 78–86. [Google Scholar] [CrossRef] [PubMed]
- van Wyk, N.; Grossmann, M.; Wendland, J.; von Wallbrunn, C.; Pretorius, I.S. The whiff of wine yeast innovation: Strategies for enhancing aroma production by yeast during wine fermentation. J. Agric. Food Chem. 2019, 67, 13496–13505. [Google Scholar] [CrossRef] [PubMed]
- Swiegers, J.H.; Capone, D.L.; Pardon, K.H.; Elsey, G.M.; Sefton, M.A.; Francis, I.L.; Pretorius, I.S. Engineering volatile thiol release in Saccharomyces cerevisiae for improved wine aroma. Yeast 2007, 24, 561–574. [Google Scholar] [CrossRef]
- Holt, S.; Cordente, A.G.; Williams, S.J.; Capone, D.L.; Jitjaroen, W.; Menz, I.R.; Curtin, C.; Anderson, P.A. Engineering Saccharomyces cerevisiae to release 3-mercaptohexan-1-ol during fermentation through overexpression of an S. cerevisiae gene, STR3, for improvement of wine aroma. Appl. Environ. Microbiol. 2011, 77, 3626–3632. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Lloyd, N.D.R.; Pretorius, I.S.; Borneman, A.R. Heterologous production of raspberry ketone in the wine yeast Saccharomyces cerevisiae via pathway engineering and synthetic enzyme fusion. Microb. Cell Fact. 2016, 15, 49:2–49:7. [Google Scholar] [CrossRef] [Green Version]
- Pardo, E.; Rico, J.; Gil, J.V.; Orejas, M. De novo production of six key grape aroma monoterpenes by a geraniol synthase-engineered Saccharomyces cerevisiae wine strain. Microb. Cell Fact. 2015, 14, 136:1–136:8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, X.F.; Li, H.P.; Wang, L.Y.; Zhang, C.; Xing, X.H.; Bao, C.Y. Atmospheric and room temperature plasma (ARTP) as a new powerful mutagenesis tool. Appl. Microbiol. Biotechnol. 2014, 98, 5387–5396. [Google Scholar] [CrossRef]
- Dragosits, M.; Mattanovich, D. Adaptive laboratory evolution—Principles and applications for biotechnology. Microb. Cell Fact. 2013, 12, 64. [Google Scholar] [CrossRef] [Green Version]
- Tian, T.; Wu, D.; Ng, C.T.; Yang, H.; Sun, J.; Liu, J.; Lu, J. A multiple-step strategy for screening Saccharomyces cerevisiae strains with improved acid tolerance and aroma profiles. Appl. Microbiol. Biotechnol. 2020, 104, 3097–3107. [Google Scholar] [CrossRef]
- Samoticha, J.; Wojdyło, A.; Chmielewska, J.; Nofer, J. Effect of different yeast strains and temperature of fermentation on basic enological parameters, polyphenols and volatile compounds of aurore white wine. Foods 2019, 8, 599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Malo, M.; García-Ríos, E.; Melgar, B.; Sanchez, M.R.; Dunham, M.J.; Guillamón, J.M. Evolutionary engineering of a wine yeast strain revealed a key role of inositol and mannoprotein metabolism during low temperature fermentation. BMC Genom. 2015, 16, 537. [Google Scholar] [CrossRef] [Green Version]
- Wiedenheft, B.; Sternberg, S.H.; Doudna, J.A. RNA-guided genetic silencing systems in bacteria and archaea. Nature 2012, 482, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Mitsui, R.; Yamada, R.; Ogino, H. CRISPR system in the yeast Saccharomyces cerevisiae and its application in the bioproduction of useful chemicals. World J. Microbiol. Biotechnol. 2019, 35, 111. [Google Scholar] [CrossRef] [PubMed]
- Adames, N.R.; Gallegos, J.E.; Peccoud, J. Yeast genetic interaction screens in the age of CRISPR/Cas. Curr. Genet. 2019, 65, 307–327. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Fang, H.; Zhang, D. Expanding application of CRISPR-Cas9 system in microorganisms. Synth. Syst. Biotechnol. 2020, 5, 269–276. [Google Scholar] [CrossRef]
- van Wyk, N.; Kroukamp, H.; Espinosa, M.I.; von Wallbrunn, C.; Wendland, J.; Pretorius, I.S. Blending wine yeast phenotypes with the aid of CRISPR DNA editing technologies. Int. J. Food Microbiol. 2020, 324, 108615:1–108615:7. [Google Scholar] [CrossRef]
- Dzialo, M.C.; Park, R.; Steensels, J.; Lievens, B.; Verstrepen, K.J. Physiology, ecology and industrial applications of aroma formation in yeast. FEMS Microbiol. Rev. 2017, 41, S95–S128. [Google Scholar] [CrossRef] [Green Version]
- Luong, J.H. Kinetics of ethanol inhibition in alcohol fermentation. Biotechnol. Bioeng. 1985, 27, 280–285. [Google Scholar] [CrossRef]
- Athes, V.; Pena y Lillo, M.; Bernard, C.; Perez-Correa, R.; Souchon, I. Comparison of experimental methods for measuring infinite dilution volatilities of aroma compounds in water/ethanol mixtures. J. Agric. Food Chem. 2004, 52, 2021–2027. [Google Scholar] [CrossRef]
- Kutyna, D.R.; Varela, C.; Henschke, P.A.; Chambers, P.J.; Stanley, G.A. Microbiological approaches to lowering ethanol concentration in wine. Trends Food Sci. Technol. 2010, 21, 293–302. [Google Scholar] [CrossRef]
- Rodrigues, A.J.; Raimbourg, T.; González, R.; Morales, P. Environmental factors influencing the efficacy of different yeast strains for alcohol level reduction in wine by respiration. LWT-Food Sci. Technol. 2016, 65, 1038–1043. [Google Scholar] [CrossRef]
- Dequin, S.; Baptista, E.; Barre, P. Acidification of grape must by Saccharomyces cerevisiae wine yeast strains genetically engineered to produce lactic acid. Am. J. Enol. Vitic. 1999, 50, 45–50. [Google Scholar]
- Goold, H.D.; Kroukamp, H.; Williams, T.C.; Paulsen, I.T.; Varela, C.; Pretorius, I.S. Yeast’s balancing act between ethanol and glycerol production in low-alcohol wines. Microb. Biotechnol. 2017, 10, 264–278. [Google Scholar] [CrossRef] [PubMed]
- Cambon, B.; Monteil, V.; Remize, F.; Camarasa, C.; Dequin, S. Effects of GPD1 overexpression in Saccharomyces cerevisiae commercial wine yeast strains lacking ALD6 genes. Appl. Environ. Microbiol. 2006, 72, 4688–4694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehsani, M.; Fernández, M.R.; Biosca, J.A.; Julien, A.; Dequin, S. Engineering of 2,3-butanediol dehydrogenase to reduce acetoin formation by glycerol-overproducing, low-alcohol Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2009, 75, 3196–3205. [Google Scholar] [CrossRef] [Green Version]
- Varela, C.; Kutyn, D.R.; Solomon, M.R.; Black, C.A.; Borneman, A.; Henschke, P.A.; Pretorius, I.S.; Chambers, P.J. Evaluation of gene modification strategies for the development of low-alcohol-wine yeasts. Appl. Environ. Microbiol. 2012, 78, 6068–6077. [Google Scholar] [CrossRef]
- Varela, C.; Schmidt, S.A.; Borneman, A.R.; Pang, C.N.I.; Krömerx, J.O.; Khan, A.; Song, X.; Hodson, M.P.; Solomon, M.; Mayr, C.M.; et al. Systems-based approaches enable identification of gene targets which improve the flavour profile of low-ethanol wine yeast strains. Metab. Eng. 2018, 49, 178–191. [Google Scholar] [CrossRef]
- Heux, S.; Cachon, R.; Dequin, S. Cofactor engineering in Saccharomyces cerevisiae: Expression of a H2O-forming NADH oxidase and impact on redox metabolism. Metab. Eng. 2006, 8, 303–314. [Google Scholar] [CrossRef]
- Heux, S.; Sablayrolles, J.M.; Cachon, R.; Dequin, S. Engineering a Saccharomyces cerevisiae wine yeast that exhibits reduced ethanol production during fermentation under controlled microoxygenation conditions. Appl. Environ. Microbiol. 2006, 72, 5822–5828. [Google Scholar] [CrossRef] [Green Version]
- Cuello, R.A.; Flores Montero, K.J.; Mercado, L.A.; Combina, M.; Ciklic, I.F. Construction of low-ethanol-wine yeasts through partial deletion of the Saccharomyces cerevisiae PDC2 gene. AMB Express 2017, 7, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malherbe, D.F.; du Toit, M.; Cordero Otero, R.R.; van Rensburg, P.; Pretorius, I.S. Expression of the Aspergillus niger glucose oxidase gene in Saccharomyces cerevisiae and its potential applications in wine production. Appl. Microbiol. Biotechnol. 2003, 61, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Rossouw, D.; Heyns, E.H.; Setati, M.E.; Bosch, S.; Bauer, F.F. Adjustment of trehalose metabolism in wine Saccharomyces cerevisiae strains to modify ethanol yields. Appl. Environ. Microbiol. 2013, 79, 5197–5207. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xu, Y. Adaptive evolution of Saccharomyces cerevisiae with enhanced ethanol tolerance for Chinese rice wine fermentation. Appl. Biochem. Biotechnol. 2014, 173, 1940–1954. [Google Scholar] [CrossRef]
- Alper, H.; Stephanopoulos, G. Global transcription machinery engineering: A new approach for improving cellular phenotype. Metab. Eng. 2007, 9, 258–267. [Google Scholar] [CrossRef]
- Xue, T.; Chen, D.; Su, Q.; Yuan, X.; Liu, K.; Huang, L.; Fang, J.; Chen, J.; He, W.; Chen, Y. Improved ethanol tolerance and production of Saccharomyces cerevisiae by global transcription machinery engineering via directed evolution of the SPT8 gene. Food Biotechnol. 2019, 33, 155–173. [Google Scholar] [CrossRef]
- Srivastava, R.; Rai, K.M.; Pandey, B.; Singh, S.P.; Sawant, S.V. Spt-ada-gcn5acetyltransferase (SAGA) complex in plants: Genome wide identification evolutionary conservation and functional determination. PLoS ONE 2015, 8, e0134709. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.W.; Li, Y.Z.; Guo, J.; Wang, Q.; Huang, S.Y.; Chen, Y.F.; Du, L.P.; Xiao, D.G. Reduced production of ethyl carbamate for wine fermentation by deleting CAR1 in Saccharomyces cerevisiae. J. Ind. Microbiol. Biotechnol. 2016, 43, 671–679. [Google Scholar] [CrossRef]
- Genbauffe, F.S.; Cooper, T.G. The urea amidolyase (DUR1,2) gene of Saccharomyces cerevisiae. DNA Seq. 1991, 2, 19–32. [Google Scholar] [CrossRef]
- Coulon, J.; Husnik, J.I.; Inglis, D.L.; van der Merwe, G.K.; Lonvaud, A.; Erasmus, D.J.; van Vuuren, H.J.J. Metabolic engineering of Saccharomyces cerevisiae to minimize the production of ethyl carbamate in wine. Am. J. Enol. Vitic. 2006, 57, 113–124. [Google Scholar]
- Wu, D.; Li, X.; Sun, J.; Cai, G.; Xie, G.; Lu, J. Effect of citrulline metabolism in Saccharomyces cerevisiae on the formation of ethyl carbamate during Chinese rice wine fermentation. J. Inst. Brew. 2018, 124, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Vigentini, I.; Gebbia, M.; Belotti, A.; Foschino, R.; Roth, F.P. CRISPR/Cas9 system as a valuable genome editing tool for wine yeasts with application to decrease urea production. Front. Microbiol. 2017, 8, 2194:1–2194:11. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Xie, W.; Li, X.; Cai, G.; Lu, J.; Xi, G. Metabolic engineering of Saccharomyces cerevisiae using the CRISPR/Cas9 system to minimize ethyl carbamate accumulation during Chinese rice wine fermentation. Appl. Microbiol. Biotechnol. 2020, 104, 4435–4444. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Cheng, Y.; Li, Y.; Du, G.; Xie, G.; Zou, H.; Zhou, J.; Chen, J. Adaptive Evolution relieves nitrogen catabolite repression and decreases urea accumulation in cultures of the Chinese rice wine yeast strain Saccharomyces cerevisiae XZ-11. J. Agric. Food Chem. 2018, 66, 9061–9069. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Hu, X. Metabolic engineering of arginine permeases to reduce the formation of urea in Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2018, 34, 47:1–47:8. [Google Scholar] [CrossRef] [PubMed]
- Feller, A.; Georis, I.; Tate, J.J.; Cooper, T.G.; Dubois, E. Alterations in the Ure2 αCap domain elicit different GATA factor responses to rapamycin treatment and nitrogen limitation. J. Biol. Chem. 2013, 288, 1841–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torrellas, M.; Rozès, N.; Aranda, A.; Matallana, E. Basal catalase activity and high glutathione levels influence the performance of non-Saccharomyces active dry wine yeasts. Food Microbiol. 2020, 92, 103589. [Google Scholar] [CrossRef]
- Bonciani, T.; De Vero, L.; Mezzetti, F.; Fay, J.C.; Giudici, P. A multi-phase approach to select new wine yeast strains with enhanced fermentative fitness and glutathione production. Appl. Microbiol. Biotechnol. 2018, 102, 2269–2278. [Google Scholar] [CrossRef]
- Mezzetti, F.; de Vero, L.; Giudici, P. Evolved Saccharomyces cerevisiae wine strains with enhanced glutathione production obtained by an evolution-based strategy. FEMS Yeast Res. 2014, 14, 977–987. [Google Scholar] [CrossRef] [Green Version]
- Fierro-Risco, J.; Rincón, A.M.; Benítez, T.; Codón, A.C. Overexpression of stress-related genes enhances cell viability and velum formation in Sherry wine yeasts. Appl. Microbiol. Biotechnol. 2013, 97, 6867–6881. [Google Scholar] [CrossRef]
- Marangon, M.; Vegro, M.; Vincenzi, S.; Lomolino, G.; de Iseppi, A.; Curioni, A. A novel method for the quantification of white wine mannoproteins by a Competitive Indirect Enzyme-Linked Lectin Sorbent Assay (CI-ELLSA). Molecules 2018, 23, 3070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Ramos, D.; Gonzalez, R. Genetic determinants of the release of mannoproteins of enological interest by Saccharomyces cerevisiae. J. Agric. Food Chem. 2006, 54, 9411–9416. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Ramos, D.; Cebollero, E.; Gonzalez, R. A recombinant Saccharomyces cerevisiae strain overproducing mannoproteins stabilizes wine against protein haze. Appl. Environ. Microbiol. 2008, 74, 5533–5540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penacho, V.; Blondin, B.; Valero, E.; González, R. Flocculation and transcriptional adaptation to fermentation conditions in a recombinant wine yeast strain defective for KNR4/SMI1. Biotechnol. Prog. 2012, 28, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Inglis, D.L.; Pickering, G.P. Vintning on thin ice—The making of Canada’s iconic dessert wine. In The World of Niagara Wine; Ripmeester, M., Mackintosh, P., Fullerton, C., Eds.; Wilfrid Laurier University Press: Waterloo, ON, Canada, 2013; pp. 229–248. ISBN 9781554584055. [Google Scholar]
- Ferreira, C.; van Voorst, F.; Martins, A.; Neves, L.; Oliveira, R.; Kielland-Brandt, M.C.; Lucas, C.; Brandt, A. A member of the sugar transporter family, Stl1p is the glycerol/H+ symporter in Saccharomyces cerevisiae. Mol. Biol. Cell 2005, 16, 2068–2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muysson, J.; Miller, L.; Allie, R.; Inglis, D.L. The use of CRISPR-Cas9 genome editing to determine the importance of glycerol uptake in wine yeast during Icewine fermentation. Fermentation 2019, 5, 93. [Google Scholar] [CrossRef]
- Capece, A.; Pietrafesa, R.; Siesto, G.; Romano, P. Biotechnological approach based on selected Saccharomyces cerevisiae starters for reducing the use of sulfur dioxide in wine. Microorganisms 2020, 8, 738. [Google Scholar] [CrossRef] [PubMed]
- Corte, L.; Roscini, L.; Zadra, C.; Antonielli, L.; Tancini, B.; Magini, A.; Emiliani, C.; Cardinali, G. Effect of pH on potassium metabisulphite biocidic activity against yeast and human cell cultures. Food Chem. 2012, 134, 1327–1336. [Google Scholar] [CrossRef]
- Hinze, H.; Holzer, H. Analysis of the energy metabolism after incubation of Saccharomyces cerevisiae with sulfite or nitrite. Arch. Microbiol. 1986, 145, 27–31. [Google Scholar] [CrossRef]
- Romano, P.; Suzzi, G. Sulphur dioxide and wine microorganisms. In Wine Microbiology and Biotechnology, 1st ed.; Fleet, G., Ed.; CRC Press: Boca Raton, FL, USA, 1993; pp. 373–393. ISBN1 3718651327. ISBN2 9783718651320. [Google Scholar]
- Fugelsang, K.; Edwards, C. Wine Microbiology; Springer: Boston, MA, USA, 2007; ISBN1 978-0-387-33341-0 (print). ISBN2 978-0-387-33349-6 (online). [Google Scholar] [CrossRef]
- Lage, P.; Sampaio-Marques, B.; Ludovico, P.; Mira, N.P.; Mendes-Ferreira, A. Transcriptomic and chemogenomic analyses unveil the essential role of Com2-regulon in response and tolerance of Saccharomyces cerevisiae to stress induced by sulfur dioxide. Microb. Cell 2019, 6, 509–523. [Google Scholar] [CrossRef]
- Park, H.; Bakalinsky, A.T. SSU1 mediates sulphite efflux in Saccharomyces cerevisiae. Yeast 2000, 16, 881–888. [Google Scholar] [CrossRef]
- García-Ríos, E.; Nuévalos, M.; Barrio, E.; Puig, S.; Guillamón, J.M. A new chromosomal rearrangement improves the adaptation of wine yeasts to sulfite. Environ. Microbiol. 2019, 21, 1771–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleiss, A.; O’Donnell, S.; Fournier, T.; Lu, W.; Agier, N.; Delmas, S.; Schacherer, J.; Fischer, G. Reshuffling yeast chromosomes with CRISPR/Cas9. PLoS Genet. 2019, 15, e1008332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordente, A.G.; Heinrich, A.; Pretorius, I.S.; Swiegers, J.D. Isolation of sulfite reductase variants of a commercial wine yeast with significantly reduced hydrogen sulfide production. FEMS Yeast Res. 2009, 9, 446–459. [Google Scholar] [CrossRef] [Green Version]
- Salvadó, Z.; Ramos-Alonso, L.; Tronchoni, J.; Penacho, V.; García-Ríos, E.; Morales, P.; Gonzalez, R.; Guillamón, J.M. Genome-wide identification of genes involved in growth and fermentation activity at low temperature in Saccharomyces cerevisiae. Int. J. Food Microbiol. 2016, 236, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Ganatsios, V.; Terpou, A.; Gialleli, A.I.; Kanellaki, M.; Bekatorou, A.; Koutinas, A.A. A ready-to-use freeze-dried juice and immobilized yeast mixture for low temperature sour cherry (Prunus cerasus) wine making. Food Bioprod. Process. 2019, 117, 373–379. [Google Scholar] [CrossRef]
- Terpou, A.; Ganatsios, V.; Kanellaki, M.; Koutinas, A.A. Entrapped psychrotolerant yeast cells within pine sawdust for low temperature wine making: Impact on wine quality. Microorganisms 2020, 8, 764. [Google Scholar] [CrossRef]
- García-Ríos, E.; Morard, M.; Parts, L.; Liti, G.; Guillamón, J.M. The genetic architecture of low-temperature adaptation in the wine yeast Saccharomyces cerevisiae. BMC Genom. 2017, 18, 159. [Google Scholar] [CrossRef] [Green Version]
- Deed, R.C.; Fedrizzi, B.; Gardner, R.C. Saccharomyces cerevisiae FLO1 gene demonstrates genetic linkage to increased fermentation rate at low temperatures. G3 (Bethesda) 2017, 7, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- García-Ríos, E.; Ramos-Alonso, L.; Guillamón, J.M. Correlation between low temperature adaptation and oxidative stress in Saccharomyces cerevisiae. Front. Microbiol. 2016, 7, 1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessi-Pérez, E.I.; Molinet, J.; Martínez, C. Disentangling the genetic bases of Saccharomyces cerevisiae nitrogen consumption and adaptation to low nitrogen environments in wine fermentation. Biol. Res. 2020, 53, 2. [Google Scholar] [CrossRef] [PubMed]
- Kessi-Pérez, E.I.; Molinet, J.; García, V.; Aguilera, O.; Cepeda, F.; López, M.A.; Sari, S.; Cuello, R.; Ciklic, I.; Rojo, M.C.; et al. Generation of a non-transgenic genetically improved yeast strain for wine production from nitrogen-deficient musts. Microorganisms 2020, 8, 1194. [Google Scholar] [CrossRef]
- Kessi-Pérez, E.I.; Ponce, B.; Li, J.; Molinet, J.; Baeza, C.; Figueroa, D.; Bastías, C.; Gaete, M.; Liti, G.; Díaz-Barrera, A.; et al. Differential Gene Expression and Allele Frequency Changes Favour Adaptation of a heterogeneous yeast population to nitrogen-limited fermentations. Front. Microbiol. 2020, 11, 1204. [Google Scholar] [CrossRef] [PubMed]
- James, R. Broach. Nutritional control of growth and development in yeast. Genetics 2012, 192, 73–105. [Google Scholar] [CrossRef] [Green Version]
- Tesnière, C.; Brice, C.; Blondin, B. Responses of Saccharomyces cerevisiae to nitrogen starvation in wine alcoholic fermentation. Appl. Microbiol. Biotechnol. 2015, 99, 7025–7034. [Google Scholar] [CrossRef]
- Vallejo, B.; Matallana, E.; Aranda, A. Saccharomyces cerevisiae nutrient signaling pathways show an unexpected early activation pattern during winemaking. Microb. Cell Fact. 2020, 19, 124. [Google Scholar] [CrossRef]
- Molinet, J.; Salinas, F.; Guillamón, J.M.; Martínez, C. GTR1 affects nitrogen consumption and TORC1 activity in Saccharomyces cerevisiae under fermentation conditions. Front. Genet. 2020, 11, 519. [Google Scholar] [CrossRef]
- Vallejo, B.; Peltier, E.; Garrigós, V.; Matallana, E.; Marullo, P.; Aranda, A. Role of Saccharomyces cerevisiae nutrient signaling pathways during winemaking: A Phenomics Approach. Front. Bioeng. Biotechnol. 2020, 8, 853. [Google Scholar] [CrossRef]
- Husnik, J.I.; Volschenk, H.; Bauer, J.; Colavizza, D.; Luo, Z.; van Vuuren, H.J. Metabolic engineering of malolactic wine yeast. Metab. Eng. 2006, 8, 315–323. [Google Scholar] [CrossRef]
- Hirosawa, I.; Aritomi, K.; Hoshida, H.; Kashiwagi, S.; Nishizawa, Y.; Akada, R. Construction of a self-cloning sake yeast that overexpresses alcohol acetyltransferase gene by a two-step gene replacement protocol. Appl. Microbiol. Biotechnol. 2004, 65, 68–73. [Google Scholar] [CrossRef]
- Fischer, S.; Procopio, S.; Becker, T. Self-cloning brewing yeast: A new dimension in beverage production. Eur. Food Res. Technol. 2013, 237, 851–863. [Google Scholar] [CrossRef]
- Steensels, J.; Snoek, T.; Meersman, E.; Picca Nicolino, M.; Voordeckers, K.; Verstrepen, K.J. Improving industrial yeast strains: Exploiting natural and artificial diversity. FEMS Microbiol. Rev. 2014, 38, 947–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Gianvito, P.; Tesnière, C.; Suzzi, G.; Blondin, B.; Tofalo, R. Different genetic responses to oenological conditions between a flocculent wine yeast and its FLO5 deleted strain: Insights from the transcriptome. Food Res. Int. 2018, 114, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Vigentini, I.; Gonzalez, R.; Tronchoni, J. Genetic improvement of wine yeasts. In Yeasts in the Production of Wine; Romano, P., Ciani, M., Fleet, G.H., Eds.; Springer: New York, NY, USA, 2019; pp. 315–342. ISBN1 978-1-4939-9780-0. ISBN2 978-1-4939-9782-4. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molina-Espeja, P. Next Generation Winemakers: Genetic Engineering in Saccharomyces cerevisiae for Trendy Challenges. Bioengineering 2020, 7, 128. https://doi.org/10.3390/bioengineering7040128
Molina-Espeja P. Next Generation Winemakers: Genetic Engineering in Saccharomyces cerevisiae for Trendy Challenges. Bioengineering. 2020; 7(4):128. https://doi.org/10.3390/bioengineering7040128
Chicago/Turabian StyleMolina-Espeja, Patricia. 2020. "Next Generation Winemakers: Genetic Engineering in Saccharomyces cerevisiae for Trendy Challenges" Bioengineering 7, no. 4: 128. https://doi.org/10.3390/bioengineering7040128
APA StyleMolina-Espeja, P. (2020). Next Generation Winemakers: Genetic Engineering in Saccharomyces cerevisiae for Trendy Challenges. Bioengineering, 7(4), 128. https://doi.org/10.3390/bioengineering7040128