
Synergistic Inorganic Carbon and Denitrification Genes Contributed to Nitrite Accumulation in a Hydrogen-Based Membrane Biofilm Reactor

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup
2.2. Biomass Enrichment
2.3. Synthetic Medium and MBfR Operation
2.4. Sampling and Analyses
2.5. Alkalinity Calculation and Electron-Equivalent Fluxes Analysis
2.6. Inorganic Carbon for Synthesis in Full Denitrification and Partial Denitrification
2.7. DNA Extraction and qPCR Analysis
2.8. 16S rRNA Sequencing and Data Analysis
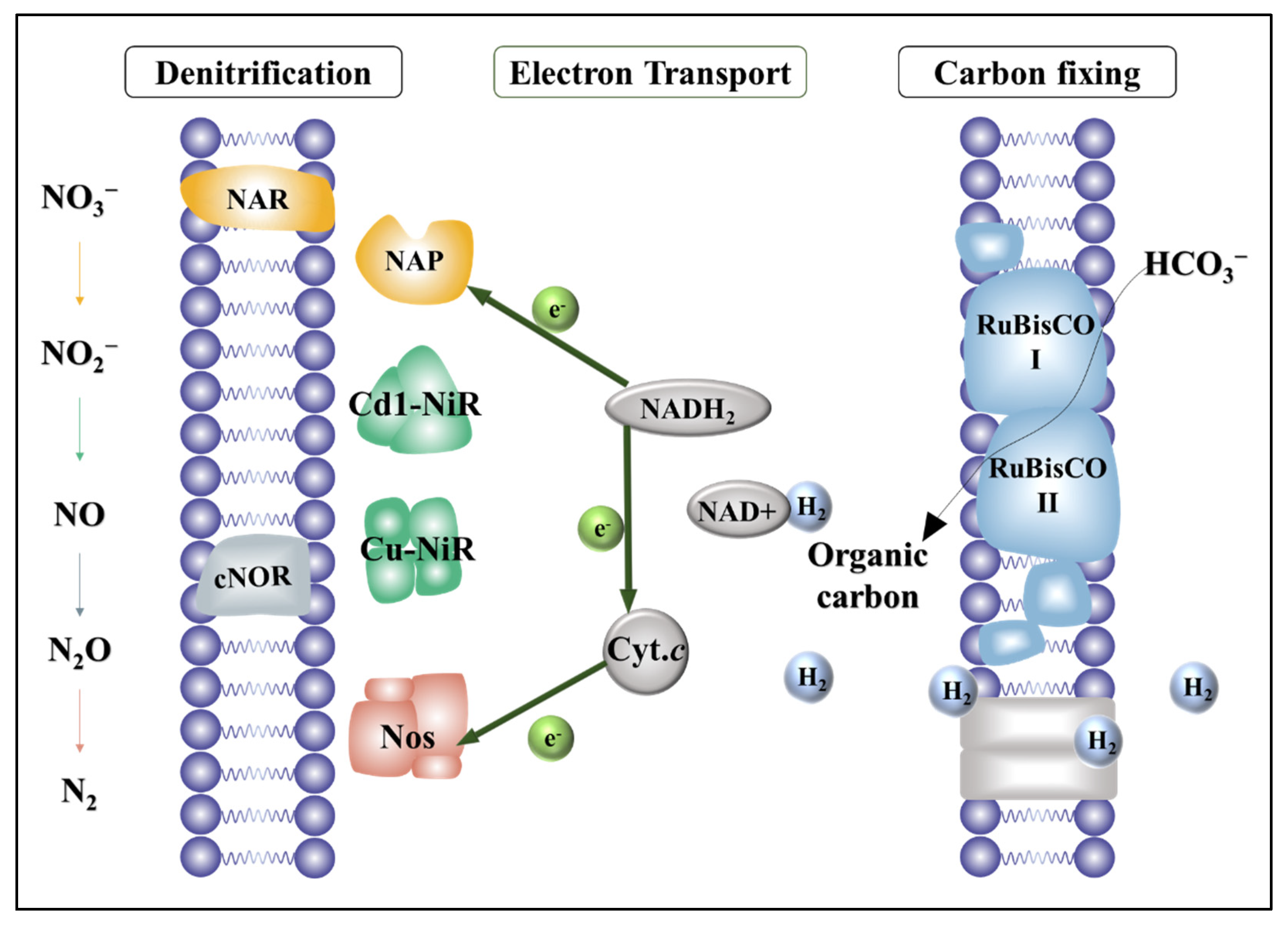
3. Results and Discussions
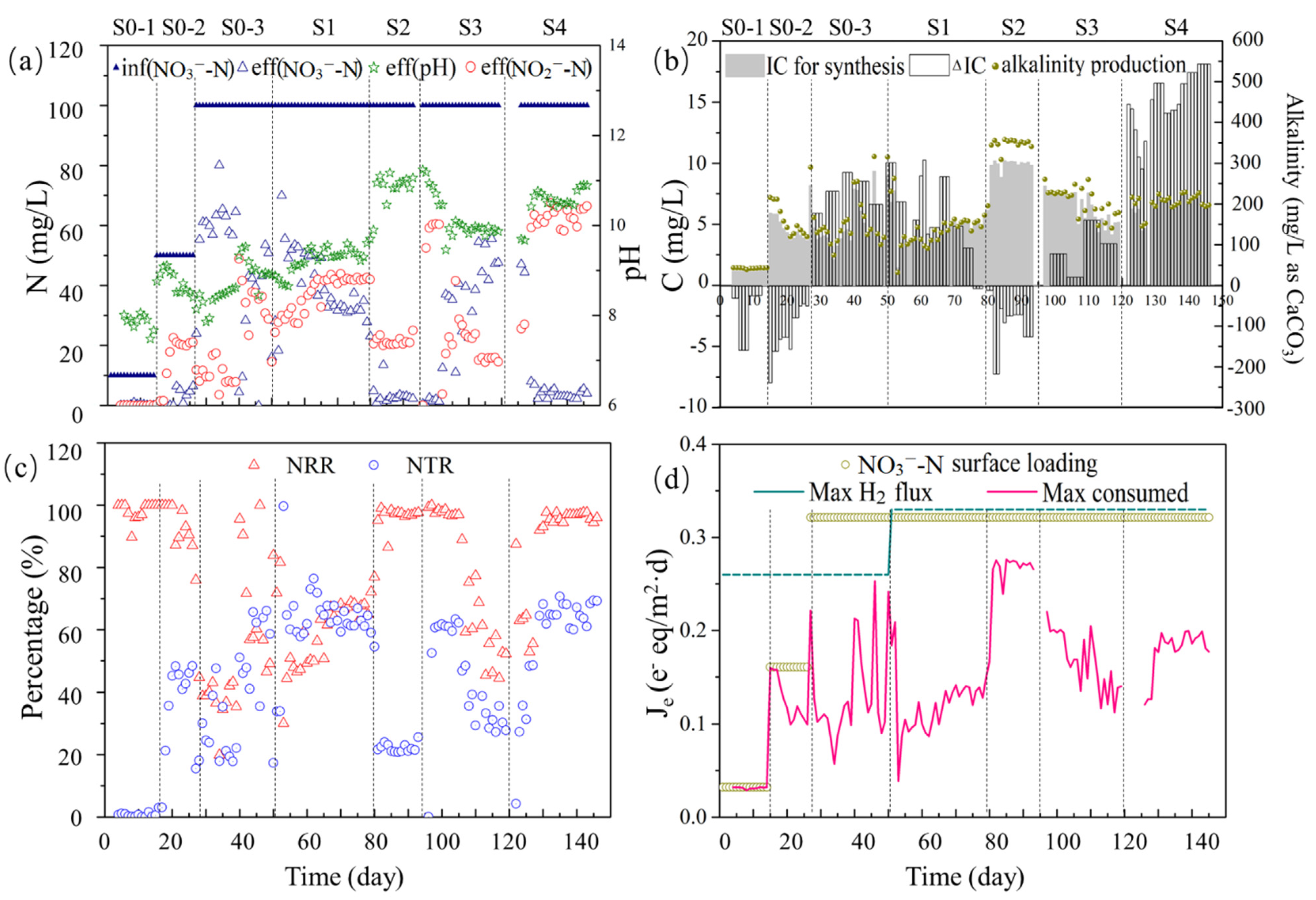
3.1. Reactor Performance
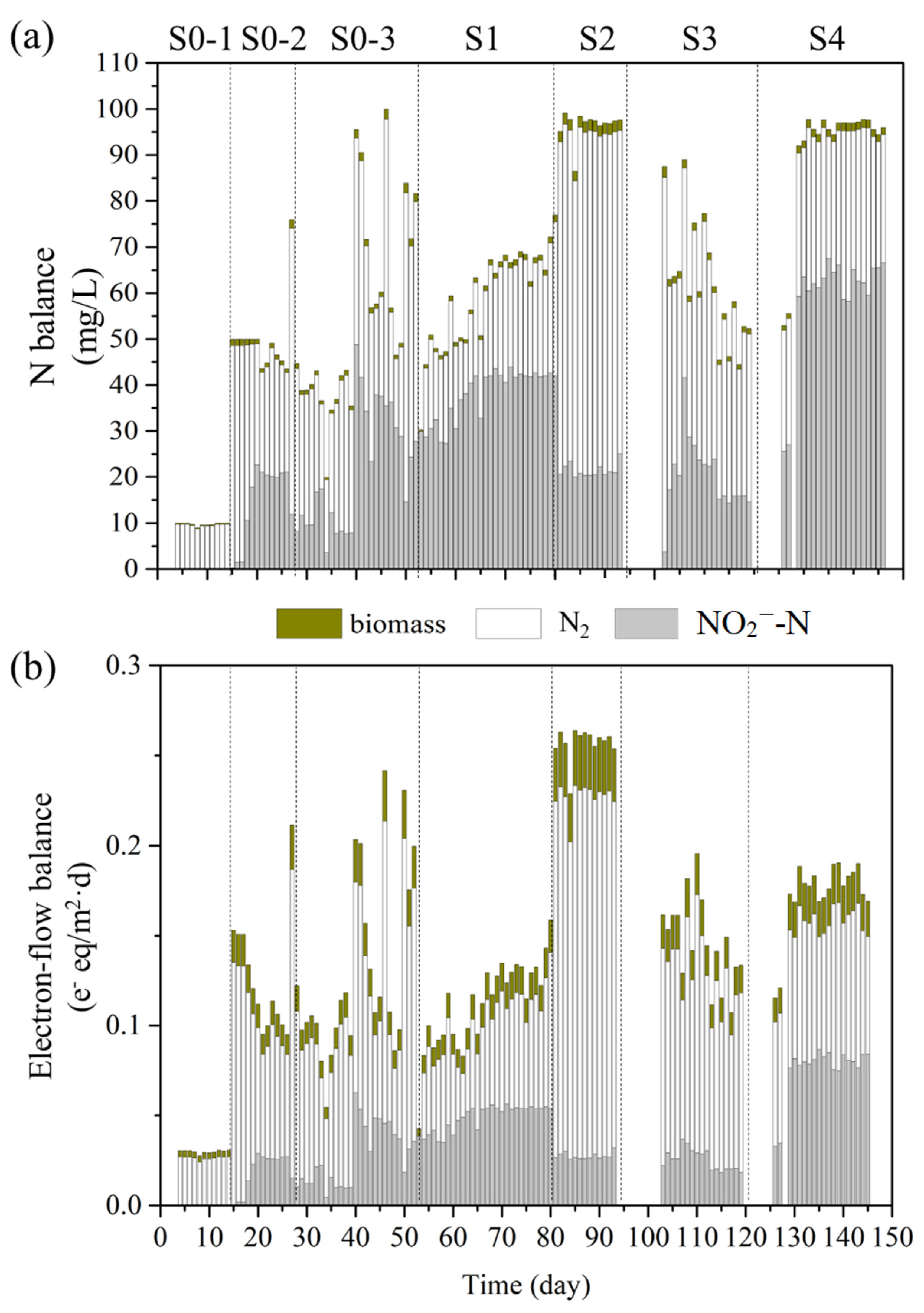
3.2. N- and Electron-Flow Balances for Full Denitrification vs. Partial Denitrification
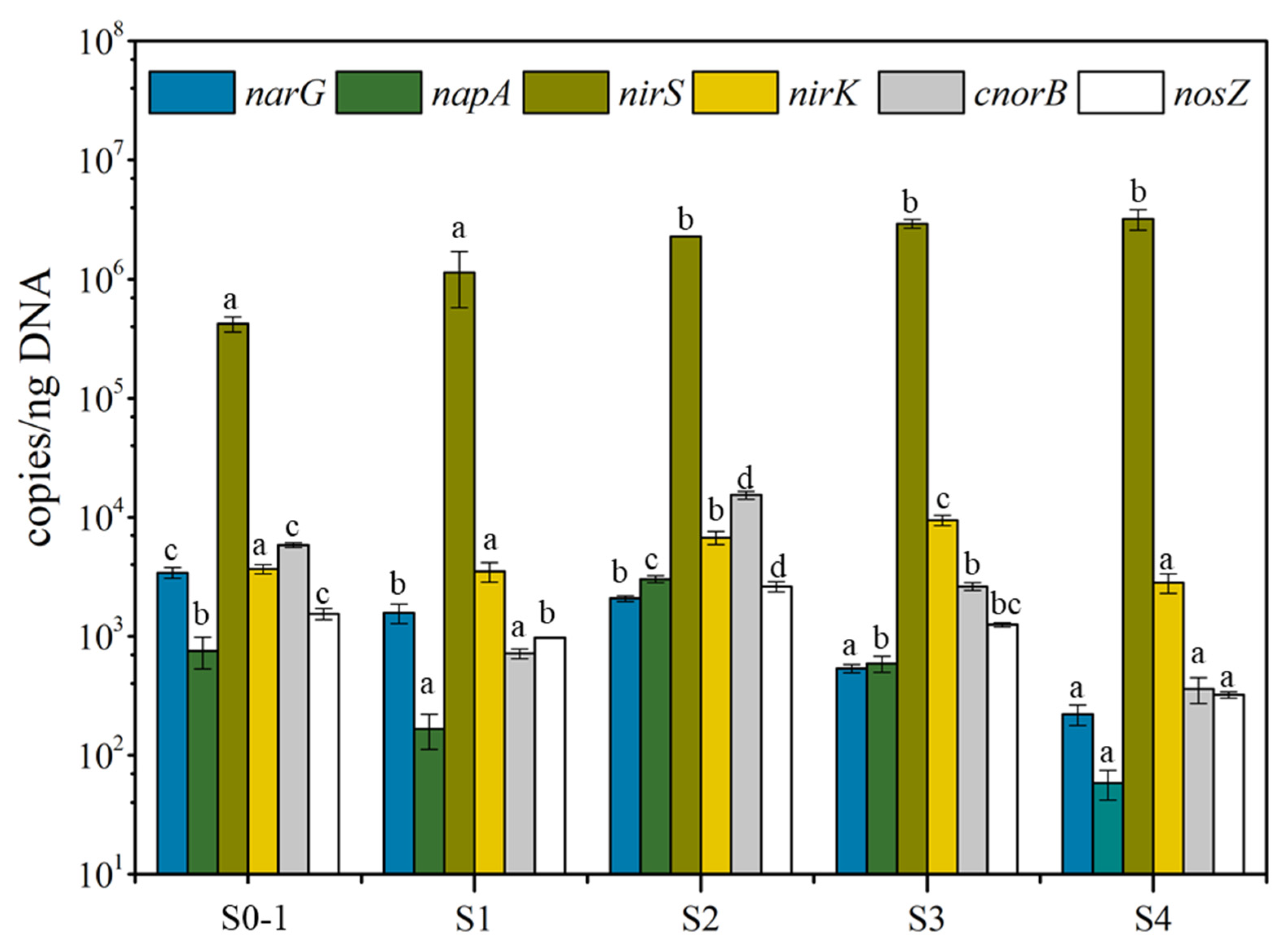
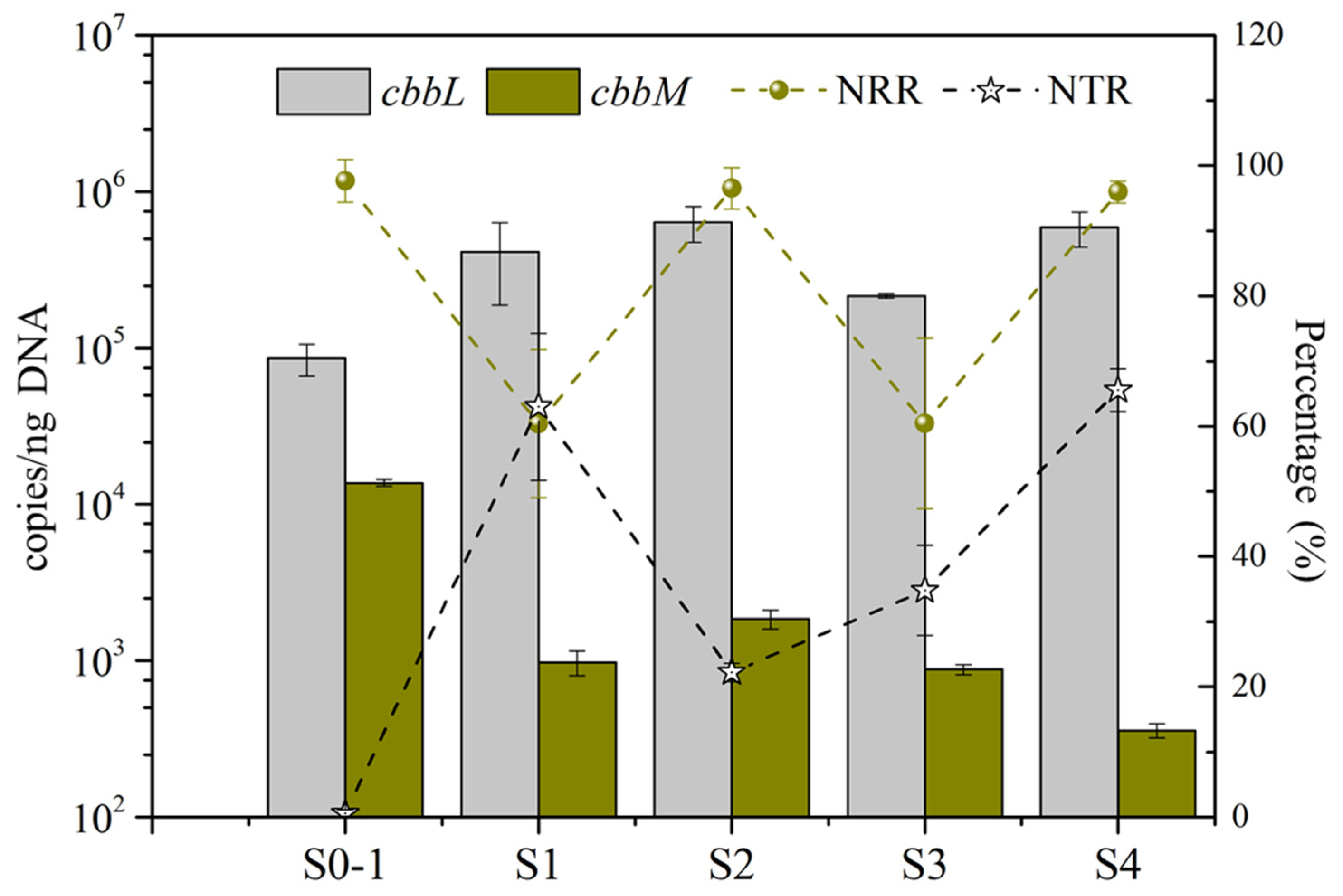
3.3. Synergy of Functional Genes
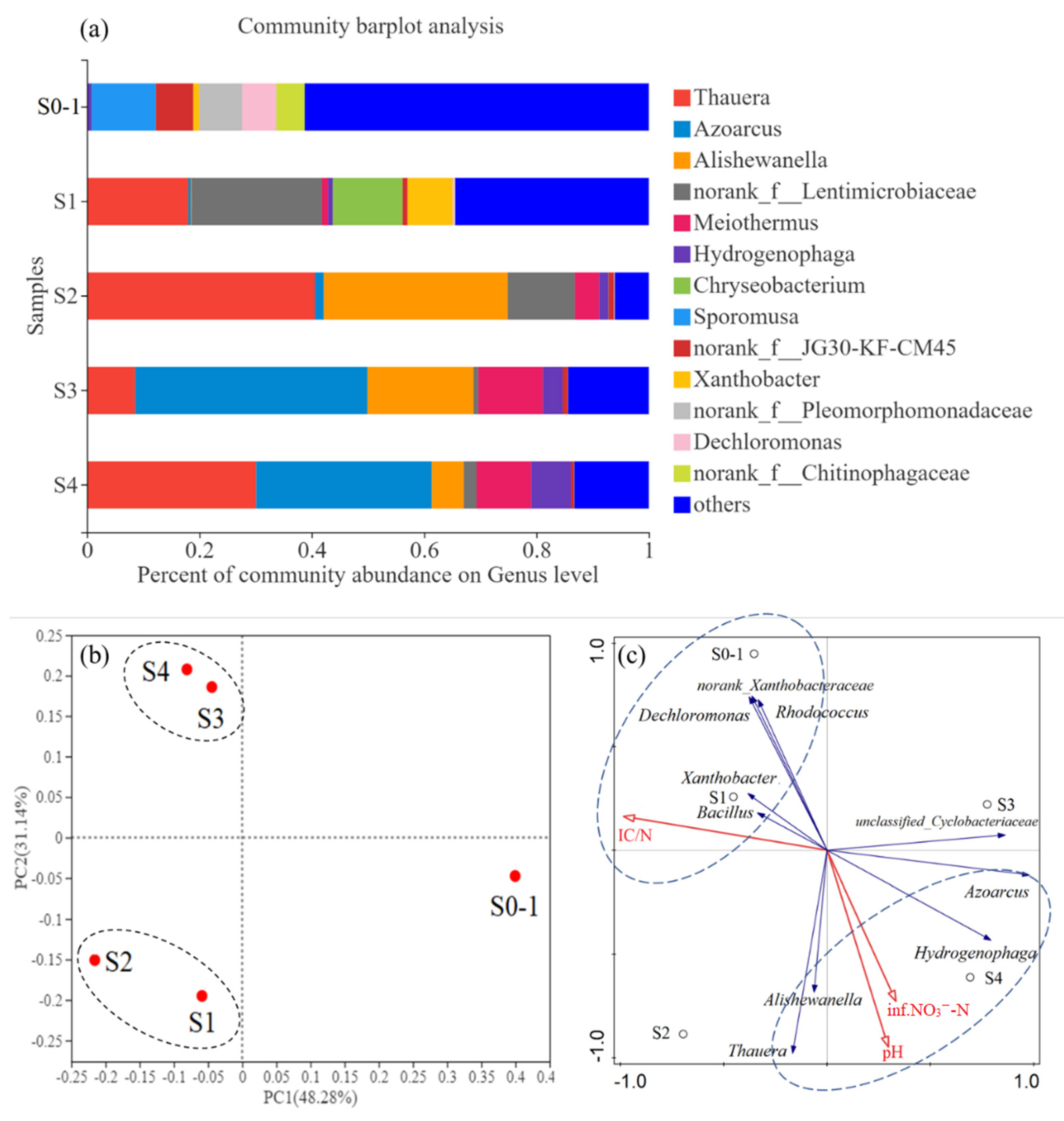
3.4. Microbial Community Characterizations of the Biofilms
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Du, R.; Cao, S.B.; Peng, Y.Z.; Zhang, H.Y.; Wang, S.Y. Combined Partial Denitrification (PD)-Anammox: A method for high nitrate wastewater treatment. Environ. Int. 2019, 126, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Lycus, P.; Bothun, K.L.; Bergaust, L.; Shapleigh, J.P.; Bakken, L.R.; Frostegard, A. Phenotypic and genotypic richness of denitrifiers revealed by a novel isolation strategy. ISME J. 2017, 11, 2219–2232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, S.B.; Du, R.; Li, B.K.; Wang, S.Y.; Ren, N.Q.; Peng, Y.Z. Nitrite production from partial-denitrification process fed with low carbon/nitrogen (C/N) domestic wastewater: Performance, kinetics and microbial community. Chem. Eng. J. 2017, 326, 1186–1196. [Google Scholar] [CrossRef]
- Li, W.; Li, H.; Liu, Y.D.; Zheng, P.; Shapleigh, J.P. Salinity-Aided Selection of Progressive Onset Denitrifiers as a Means of Providing Nitrite for Anammox. Environ. Sci. Technol. 2018, 52, 10665–10672. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Qiao, S.; Yang, Y.; Jin, R.F.; Zhou, J.T.; Rittmann, B.E. Energy recovery in the form of N2O by denitrifying bacteria. Chem. Eng. J. 2019, 371, 500–506. [Google Scholar] [CrossRef]
- Tang, Y.N.; Zhou, C.; Ziv-El, M.; Rittmann, B.E. A pH-control model for heterotrophic and hydrogen-based autotrophic denitrification. Water Res. 2011, 45, 232–240. [Google Scholar] [CrossRef]
- Lee, K.C.; Rittmann, B.E. Effects of pH and precipitation on autohydrogenotrophic denitrification using the hollow-fiber membrane-biofilm reactor. Water Res. 2003, 37, 1551–1556. [Google Scholar] [CrossRef]
- Tang, Y.N.; Zhou, C.; Van Ginkel, S.W.; Ontiveros-Valencia, A.; Shin, J.; Rittmann, B.E. Hydrogen permeability of the hollow fibers used in H-2-based membrane biofilm reactors. J. Membr. Sci. 2012, 407, 176–183. [Google Scholar] [CrossRef]
- Ziv-El, M.C.; Rittmann, B.E. Systematic evaluation of nitrate and perchlorate bioreduction kinetics in groundwater using a hydrogen-based membrane biofilm reactor. Water Res. 2009, 43, 173–181. [Google Scholar] [CrossRef]
- Moreno-Vivian, C.; Cabello, P.; Martinez-Luque, M.; Blasco, R.; Castillo, F. Prokaryotic nitrate reduction: Molecular properties and functional distinction among bacterial nitrate reductases. J. Bacteriol. 1999, 181, 6573–6584. [Google Scholar] [CrossRef] [Green Version]
- Flanagan, D.A.; Gregory, L.G.; Carter, J.P.; Karakas-Sen, A.; Richardson, D.J.; Spiro, S. Detection of genes for periplasmic nitrate reductase in nitrate respiring bacteria and in community DNA. FEMS Microbiol. Lett. 1999, 177, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Levy-Booth, D.J.; Prescott, C.E.; Grayston, S.J. Microbial functional genes involved in nitrogen fixation, nitrification and denitrification in forest ecosystems. Soil Biol. Biochem. 2014, 75, 11–25. [Google Scholar] [CrossRef]
- Zhang, K.Y.; Gu, J.; Wang, X.J.; Zhang, X.; Hu, T.; Zhao, W.Y. Analysis for microbial denitrification and antibiotic resistance during anaerobic digestion of cattle manure containing antibiotic. Bioresour. Technol. 2019, 291, 121803. [Google Scholar] [CrossRef] [PubMed]
- Chee-Sanford, J.C.; Connor, L.; Krichels, A.; Yang, W.H.; Sanford, R.A. Hierarchical detection of diverse Clade II (atypical) nosZ genes using new primer sets for classical- and multiplex PCR array applications. J. Microbiol. Meth. 2020, 172, 105908. [Google Scholar] [CrossRef]
- Chon, K.; Chang, J.S.; Lee, E.; Lee, J.; Ryu, J.; Cho, J. Abundance of denitrifying genes coding for nitrate (narG), nitrite (nirS), and nitrous oxide (nosZ) reductases in estuarine versus wastewater effluent-fed constructed wetlands. Ecol. Eng. 2011, 37, 64–69. [Google Scholar] [CrossRef]
- Ge, S.J.; Peng, Y.Z.; Wang, S.Y.; Lu, C.C.; Cao, X.; Zhu, Y.P. Nitrite accumulation under constant temperature in anoxic denitrification process: The effects of carbon sources and COD/NO3-N. Bioresour. Technol. 2012, 114, 137–143. [Google Scholar] [CrossRef]
- Du, R.; Cao, S.B.; Li, B.K.; Niu, M.; Wang, S.Y.; Peng, Y.Z. Performance and microbial community analysis of a novel DEAMOX based on partial-denitrification and anammox treating ammonia and nitrate wastewaters. Water Res. 2017, 108, 46–56. [Google Scholar] [CrossRef]
- Le, T.; Peng, B.; Su, C.Y.; Massoudieh, A.; Torrents, A.; Al-Omari, A.; Murthy, S.; Wett, B.; Chandran, K.; DeBarbadillo, C.; et al. Impact of carbon source and COD/N on the concurrent operation of partial denitrification and anammox. Water Environ. Res. 2019, 91, 185–197. [Google Scholar] [CrossRef]
- Si, Z.; Peng, Y.Z.; Yang, A.M.; Zhang, S.J.; Li, B.K.; Wang, B.; Wang, S.Y. Rapid nitrite production via partial denitrification: Pilot-scale operation and microbial community analysis. Environ. Sci.-Water Res. 2018, 4, 80–86. [Google Scholar] [CrossRef]
- Hussain, S.; Min, Z.; Zhu, X.X.; Khan, M.H.; Li, L.F.; Cao, H. Significance of Fe(II) and environmental factors on carbon-fixing bacterial community in two paddy soils. Ecotoxicol. Environ. Saf. 2019, 182, 109456. [Google Scholar] [CrossRef]
- Benson, A.A.; Nonomura, A.M.; Gerard, V.A. The Path of Carbon in Photosynthesis. XXV. Plant and Algal Growth Responses to Glycopyranosides. J. Plant Nutr. 2009, 32, 1185–1200. [Google Scholar] [CrossRef] [Green Version]
- Tabita, F.R. Microbial ribulose 1,5-bisphosphate carboxylase/oxygenase: A different perspective. Photosynth. Res. 1999, 60, 1–28. [Google Scholar] [CrossRef]
- Campbell, B.J.; Cary, S.C. Abundance of reverse tricarboxylic acid cycle genes in free-living microorganisms at deep-sea hydrothermal vents. Appl. Environ. Microb. 2004, 70, 6282–6289. [Google Scholar] [CrossRef] [Green Version]
- Nanba, K.; King, G.M.; Dunfield, K. Analysis of facultative lithotroph distribution and diversity on volcanic deposits by use of the large subunit of ribulose 1,5-bisphosphate carboxylase/oxygenase. Appl. Environ. Microb. 2004, 70, 2245–2253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.Z.; Wang, C.; Liu, Q.; Zhu, Z.K.; Lynn, T.M.; Shen, J.L.; Whiteley, A.S.; Kumaresan, D.; Ge, T.D.; Wu, J.S. Abundance of microbial CO2-fixing genes during the late rice season in a long-term management paddy field amended with straw and straw-derived biochar. Can. J. Soil Sci. 2018, 98, 306–316. [Google Scholar] [CrossRef]
- Xia, S.Q.; Wang, C.H.; Xu, X.Y.; Tang, Y.N.; Wang, Z.W.; Gu, Z.L.; Zhou, Y. Bioreduction of nitrate in a hydrogen-based membrane biofilm reactor using CO2 for pH control and as carbon source. Chem. Eng. J. 2015, 276, 59–64. [Google Scholar] [CrossRef]
- Chung, J.; Nerenberg, R.; Rittmann, B.E. Bio-reduction of soluble chromate using a hydrogen-based membrane biofilm reactor. Water Res. 2006, 40, 1634–1642. [Google Scholar] [CrossRef]
- APHA. Standard Methods for Water and Wastewater Examination; American Public Health Association: Washington, DC, USA, 2005. [Google Scholar]
- Xia, S.Q.; Zhang, Y.H.; Zhong, F.H. A continuous stirred hydrogen-based polyvinyl chloride membrane biofilm reactor for the treatment of nitrate contaminated drinking water. Bioresour. Technol. 2009, 100, 6223–6228. [Google Scholar] [CrossRef]
- Rittmann, B.E.; McCarty, P.L. Environmental Biotechnology: Principles and Applications, 2nd ed.; McGraw-Hill Companies, Inc.: New York, NY, USA, 2020. [Google Scholar]
- Xia, S.Q.; Wu, C.Y.; Yang, X.X.; Zhou, Y.; Zhou, L.M.; Ran, Y.J.; Rittmann, B.E. Bioreduction of nitrate in high-sulfate water using a hydrogen-based membrane biofilm reactor equipped with a separate carbon dioxide module. Chem. Eng. J. 2020, 385, 123831. [Google Scholar] [CrossRef]
- McNamara, D.P.; Whitney, K.M.; Goss, S.L. Use of a physiologic bicarbonate buffer system for dissolution characterization of ionizable drugs. Pharm. Res. 2003, 20, 1641–1646. [Google Scholar] [CrossRef]
- Qian, W.T.; Ma, B.; Li, X.Y.; Zhang, Q.; Peng, Y.Z. Long-term effect of pH on denitrification: High pH benefits achieving partial-denitrification. Bioresour. Technol. 2019, 278, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Z.; Zhang, Y.; Chen, Y.G. Comparative Metagenomic and Metatranscriptomic Analyses Reveal the Functional Species and Metabolic Characteristics of an Enriched Denitratation Community. Environ. Sci. Technol. 2020, 54, 14312–14321. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.H.; Cicek, N.; Oleszkiewicz, J.A. Inorganic precipitation during autotrophic denitrification under various operating conditions. Environ. Technol. 2009, 30, 1475–1485. [Google Scholar] [CrossRef] [PubMed]
- Van Ginkel, S.W.; Kim, B.O.; Yang, Z.M.; Sittmann, R.; Sholin, M.; Micelli, J.; Rittmann, B.E. Effect of NaCl on nitrate removal from ion-exchange spent brine in the membrane biofilm reactor (MBfR). Water Sci. Technol. 2012, 65, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.B.; Wang, S.Y.; Peng, Y.Z.; Wu, C.C.; Du, R.; Gong, L.X.; Ma, B. Achieving partial denitrification with sludge fermentation liquid as carbon source: The effect of seeding sludge. Bioresour. Technol. 2013, 149, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Du, R.; Peng, Y.Z.; Cao, S.B.; Li, B.K.; Wang, S.Y.; Niu, M. Mechanisms and microbial structure of partial denitrification with high nitrite accumulation. Appl. Microbiol. Biotechnol. 2016, 100, 2011–2021. [Google Scholar] [CrossRef]
- Zumft, W.G. Cell biology and molecular basis of denitrification. Microbiol. Mol. Biol. Rev. 1997, 61, 533–616. [Google Scholar]
- Li, W.; Shan, X.Y.; Wang, Z.Y.; Lin, X.Y.; Li, C.X.; Cai, C.Y.; Abbas, G.; Zhang, M.; Shen, L.D.; Hu, Z.Q.; et al. Effect of self-alkalization on nitrite accumulation in a high-rate denitrification system: Performance, microflora and enzymatic activities. Water Res. 2016, 88, 758–765. [Google Scholar] [CrossRef]
- Joshi, H.M.; Tabita, F.R. A global two component signal transduction system that integrates the control of photosynthesis, carbon dioxide assimilation, and nitrogen fixation. Proc. Natl. Acad. Sci. USA 1996, 93, 14515–14520. [Google Scholar] [CrossRef] [Green Version]
- Baker, S.H.; Jin, S.M.; Aldrich, H.C.; Howard, G.T.; Shively, J.M. Insertion mutation of the form I cbbL gene encoding ribulose bisphosphate carboxylase/oxygenase (RuBisCO) in Thiobacillus neapolitanus results in expression of form II RuBisCO, loss of carboxysomes, and an increased CO2 requirement for growth. J. Bacteriol. 1998, 180, 4133–4139. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.B.; Mao, Y.J.; Bergaust, L.; Bakken, L.R.; Frostegard, A. Strains in the genus Thauera exhibit remarkably different denitrification regulatory phenotypes. Environ. Microbiol. 2013, 15, 2816–2828. [Google Scholar] [PubMed]
- Throback, I.N.; Enwall, K.; Jarvis, A.; Hallin, S. Reassessing PCR primers targeting nirS, nirK and nosZ genes for community surveys of denitrifying bacteria with DGGE. FEMS Microbiol. Ecol. 2004, 49, 401–417. [Google Scholar] [CrossRef] [PubMed]
- Braker, G.; Fesefeldt, A.; Witzel, K.P. Development of PCR primer systems for amplification of nitrite reductase genes (nirK and nirS) to detect denitrifying bacteria in environmental samples. Appl. Environ. Microb. 1998, 64, 3769–3775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braker, G.; Tiedje, J.M. Nitric oxide reductase (norB) genes from pure cultures and environmental samples. Appl. Environ. Microb. 2003, 69, 3476–3483. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Gutierrez, J.C.; Henry, S.; Hallet, S.; Martin-Laurent, F.; Catroux, G.; Philippot, L. Quantification of a novel group of nitrate-reducing bacteria in the environment by real-time PCR. J. Microbiol. Meth. 2004, 57, 399–407. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Influent NaNO3-N (mg/L) | Influent NaHCO3-C (mg/L) | H2 Gauge Pressure (MPa) | Effluent pH |
|---|---|---|---|---|
| S0-1 | 10 | 9 | 0.04 | 7.86 ± 0.18 |
| S0-2 | 50 | 43 | 0.04 | 8.75 ± 0.25 |
| S0-3 | 100 | 86 | 0.04 | 8.68 ± 0.44 |
| S1 | 100 | 86 | 0.08 | 9.25 ± 0.28 |
| S2 | 100 | 86 | 0.08 | 10.85 ± 0.33 |
| S3 | 100 | 43 | 0.08 | 9.93 ± 0.09 |
| S4 | 100 | 43 | 0.08 | 10.62 ± 0.17 |
| Thauera | Azoarcus | Alishewanella | Hydrogenophaga | NRR | NTR | cbbL | cbbM | nirK | |
|---|---|---|---|---|---|---|---|---|---|
| Thauera | 1 | −0.065 | 0.594 | 0.325 | 0.322 | 0.383 | 0.982 ** | −0.633 | −0.040 |
| Azoarcus | 1 | 0.136 | 0.772 | −0.289 | 0.354 | −0.020 | −0.473 | 0.451 | |
| Alishewanella | 1 | 0.009 | 0.119 | −0.239 | 0.453 | −0.387 | 0.723 | ||
| Hydrogenophaga | 1 | 0.182 | 0.552 | 0.411 | −0.483 | −0.089 | |||
| NRR | 1 | −0.410 | 0.264 | 0.447 | −0.407 | ||||
| NTR | 1 | 0.537 | −0.790 | −0.286 | |||||
| cbbL | 1 | −0.691 | −0.155 | ||||||
| cbbM | 1 | −0.275 | |||||||
| nirK | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pang, S.; Rittmann, B.E.; Wu, C.; Yang, L.; Zhou, J.; Xia, S. Synergistic Inorganic Carbon and Denitrification Genes Contributed to Nitrite Accumulation in a Hydrogen-Based Membrane Biofilm Reactor. Bioengineering 2022, 9, 222. https://doi.org/10.3390/bioengineering9050222
Pang S, Rittmann BE, Wu C, Yang L, Zhou J, Xia S. Synergistic Inorganic Carbon and Denitrification Genes Contributed to Nitrite Accumulation in a Hydrogen-Based Membrane Biofilm Reactor. Bioengineering. 2022; 9(5):222. https://doi.org/10.3390/bioengineering9050222
Chicago/Turabian StylePang, Si, Bruce E. Rittmann, Chengyang Wu, Lin Yang, Jingzhou Zhou, and Siqing Xia. 2022. "Synergistic Inorganic Carbon and Denitrification Genes Contributed to Nitrite Accumulation in a Hydrogen-Based Membrane Biofilm Reactor" Bioengineering 9, no. 5: 222. https://doi.org/10.3390/bioengineering9050222
APA StylePang, S., Rittmann, B. E., Wu, C., Yang, L., Zhou, J., & Xia, S. (2022). Synergistic Inorganic Carbon and Denitrification Genes Contributed to Nitrite Accumulation in a Hydrogen-Based Membrane Biofilm Reactor. Bioengineering, 9(5), 222. https://doi.org/10.3390/bioengineering9050222