
Medical Grade Honey as a Promising Treatment to Improve Ovarian Tissue Transplantation

 , , ,
, , ,  , ,
, ,  ,
,  , and
, and 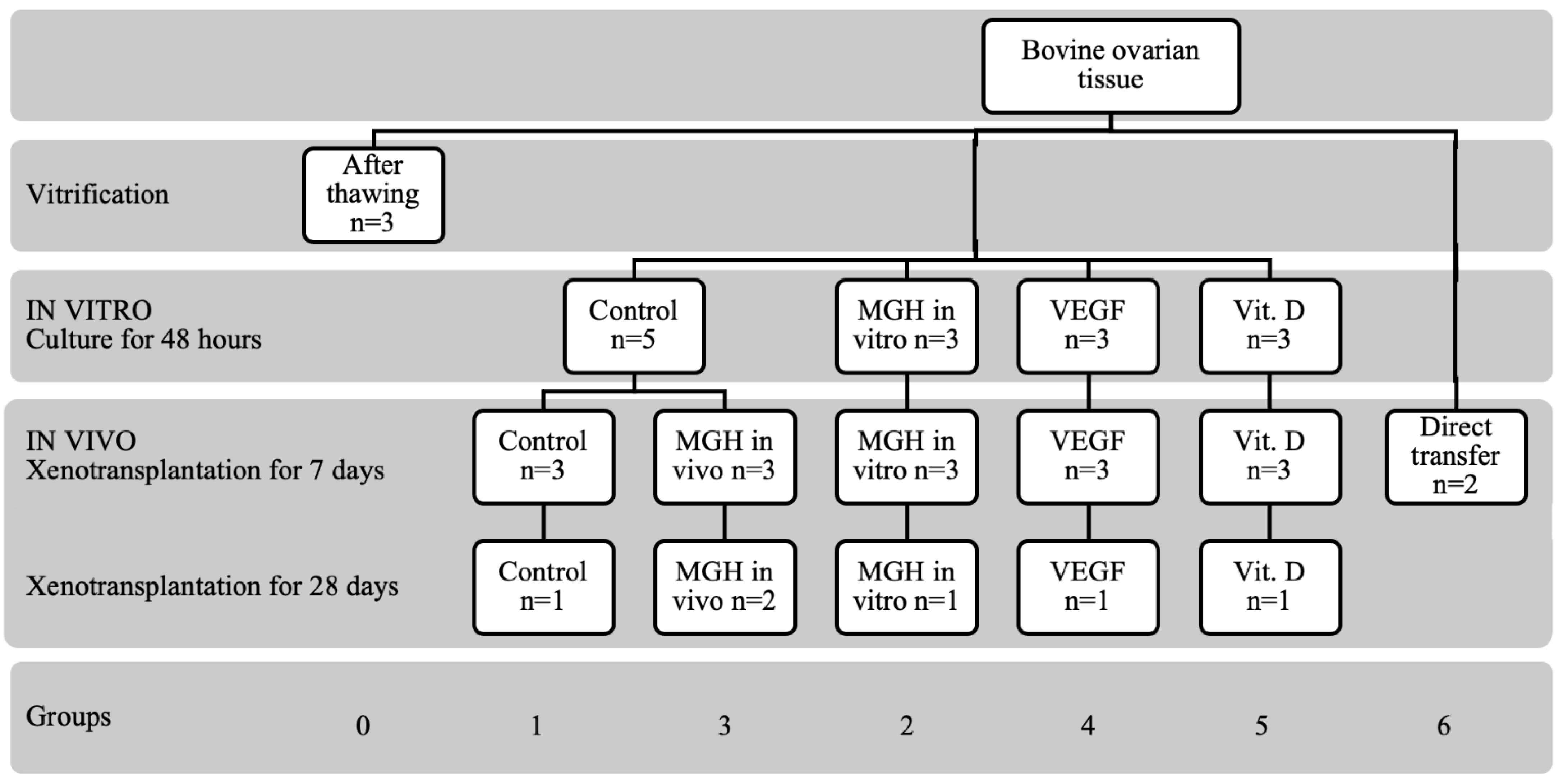{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Study Design
2.3. Ovaries Collection, Vitrification, Thawing and Culture
2.4. Experimental Animals
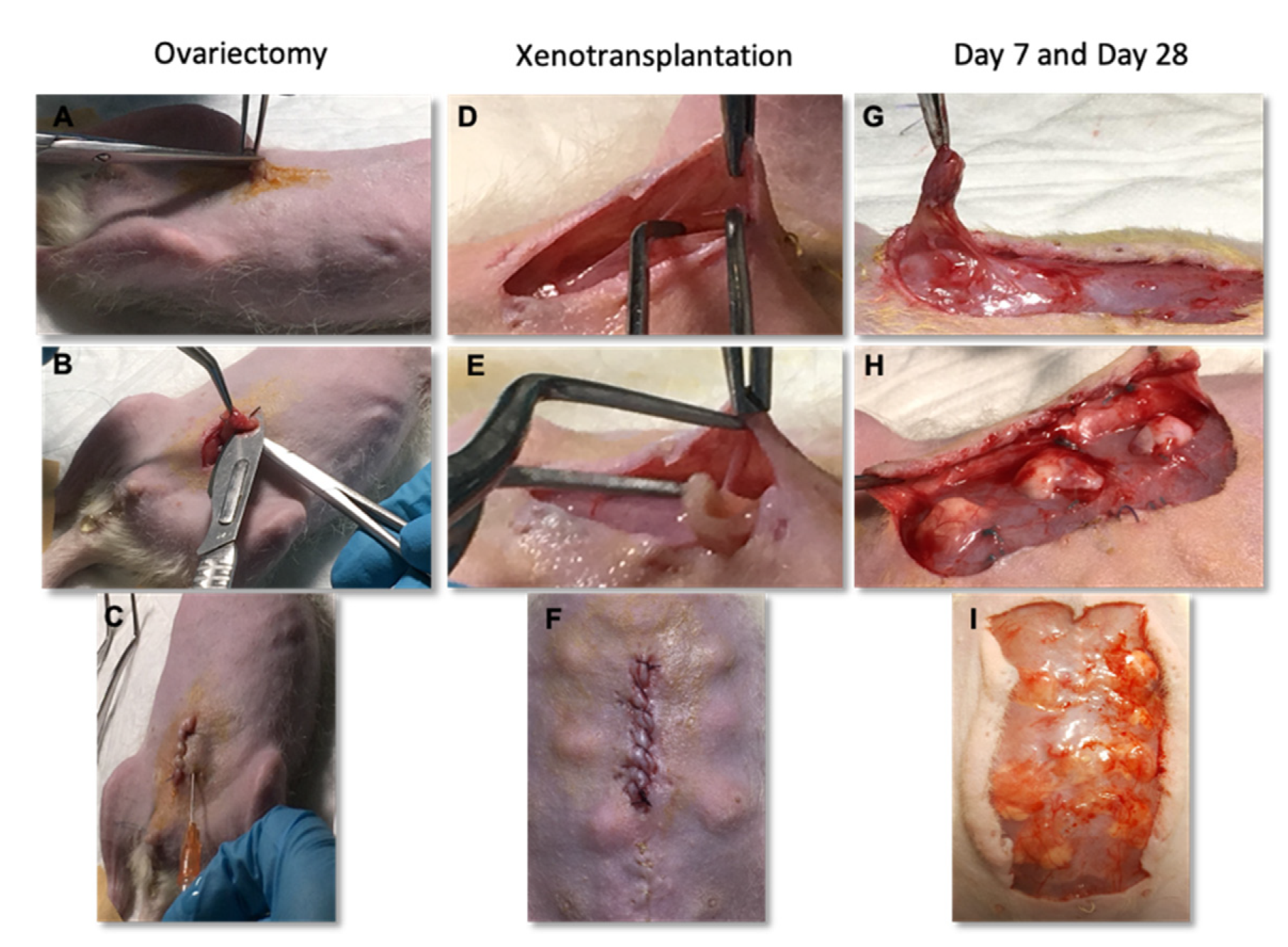
2.5. Ovariectomy
2.6. Xenotransplantation
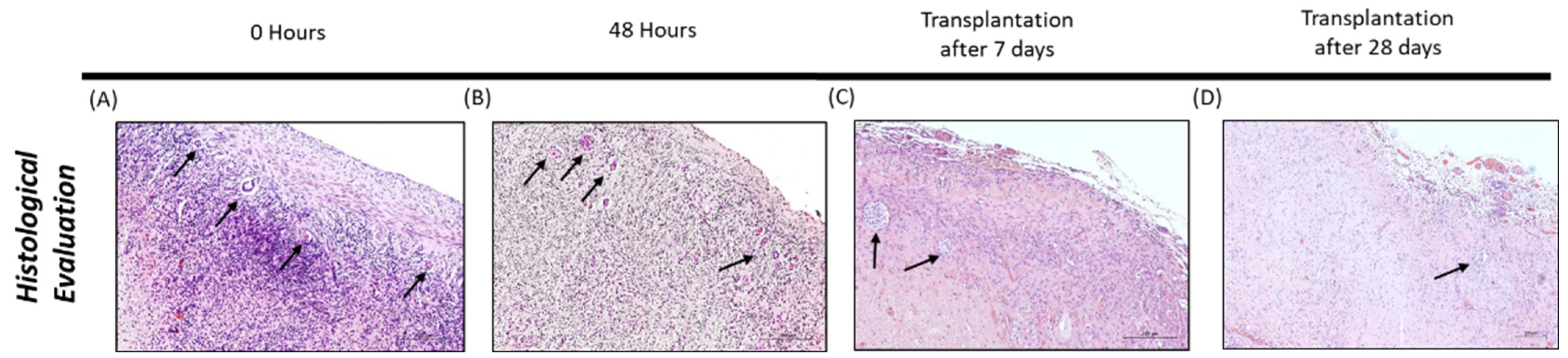
2.7. Histological Evaluation
2.8. Immunohistochemistry Evaluation
2.9. Statistical Analyses
3. Results
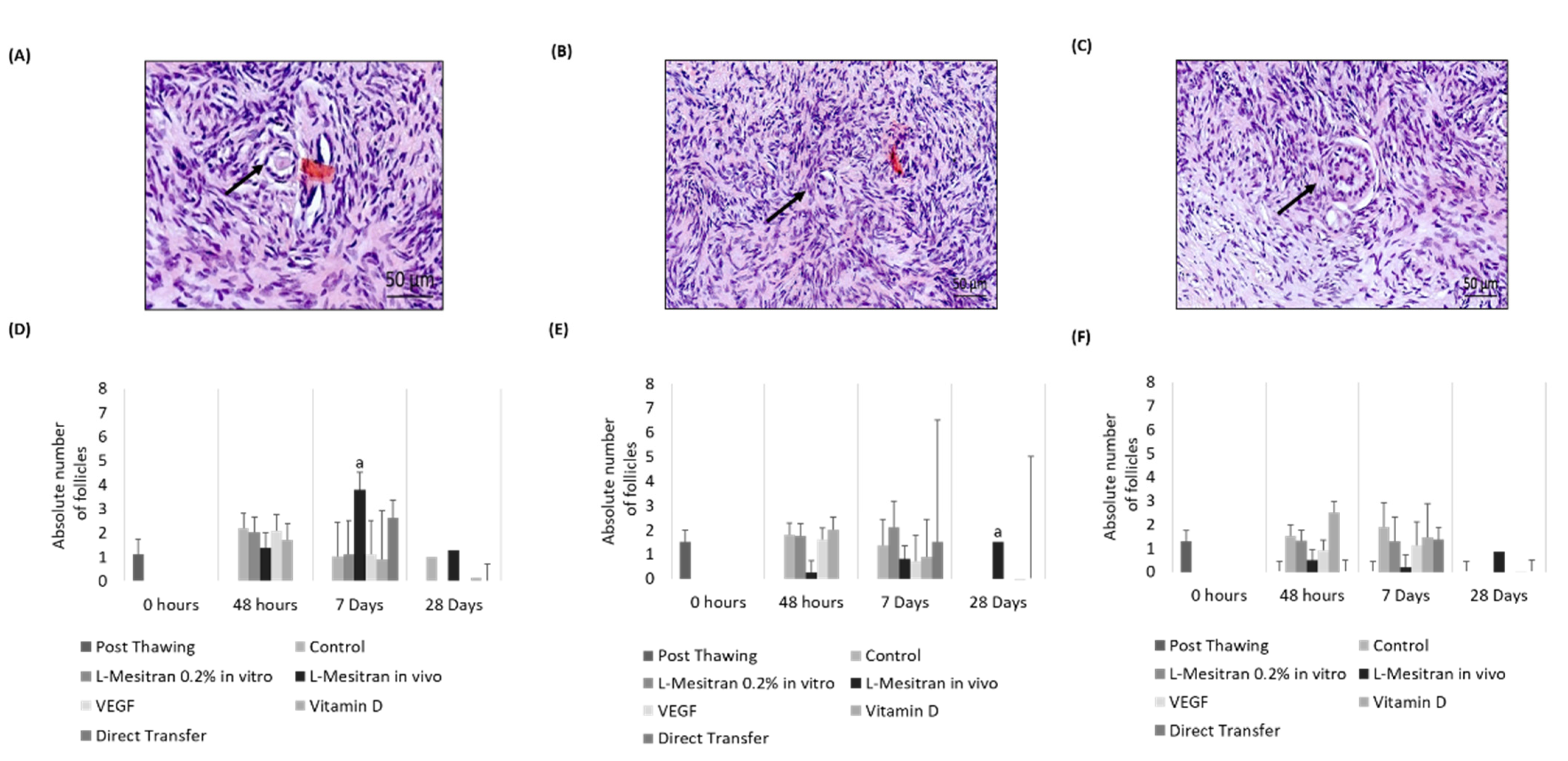
3.1. Histological Evaluation
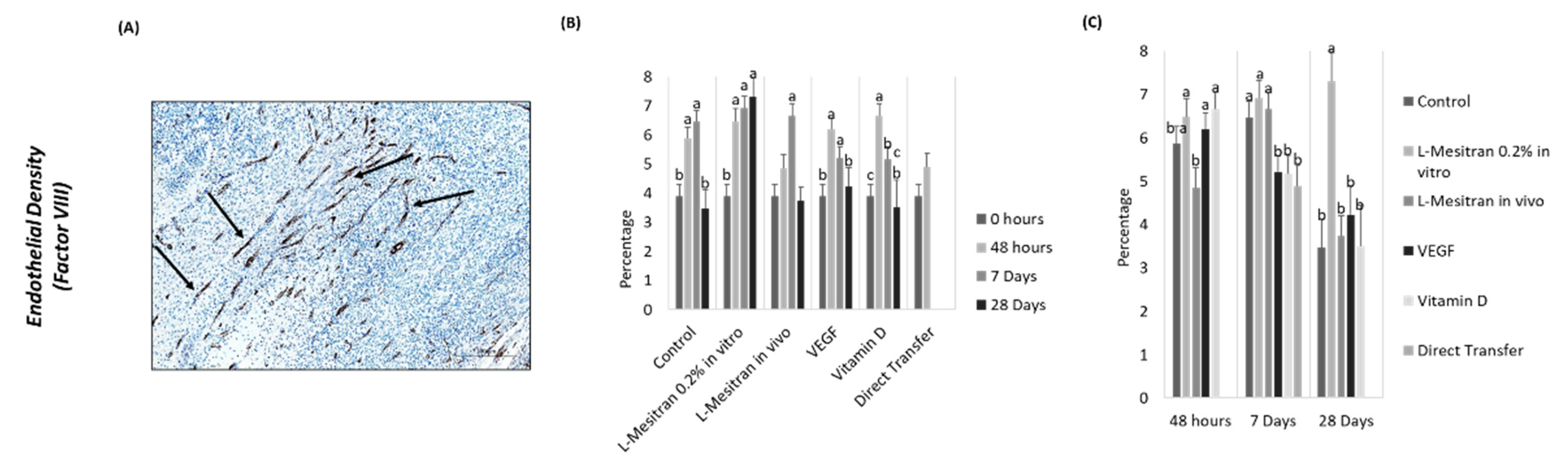
3.2. Endothelial Density
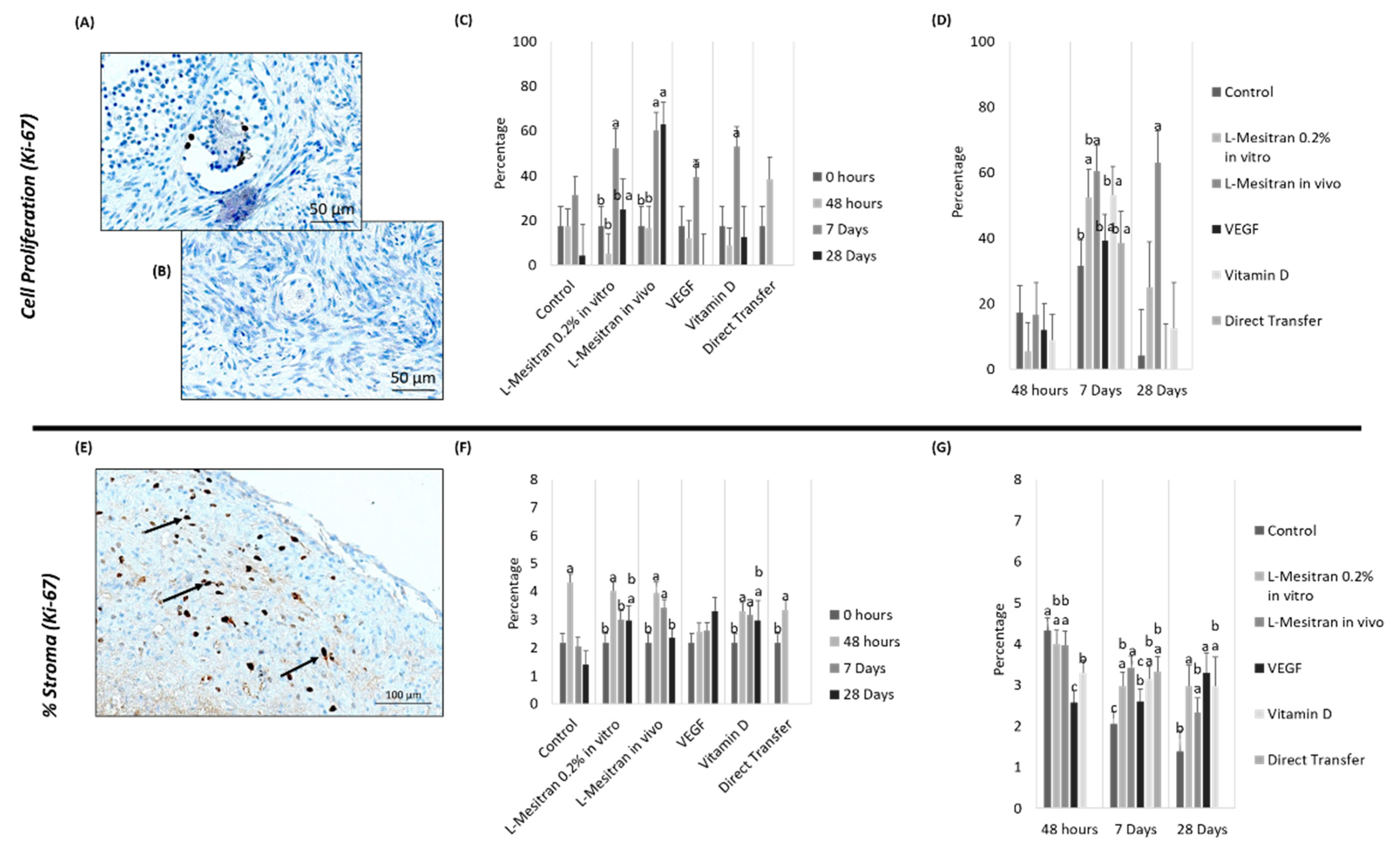
3.3. Cell Proliferation
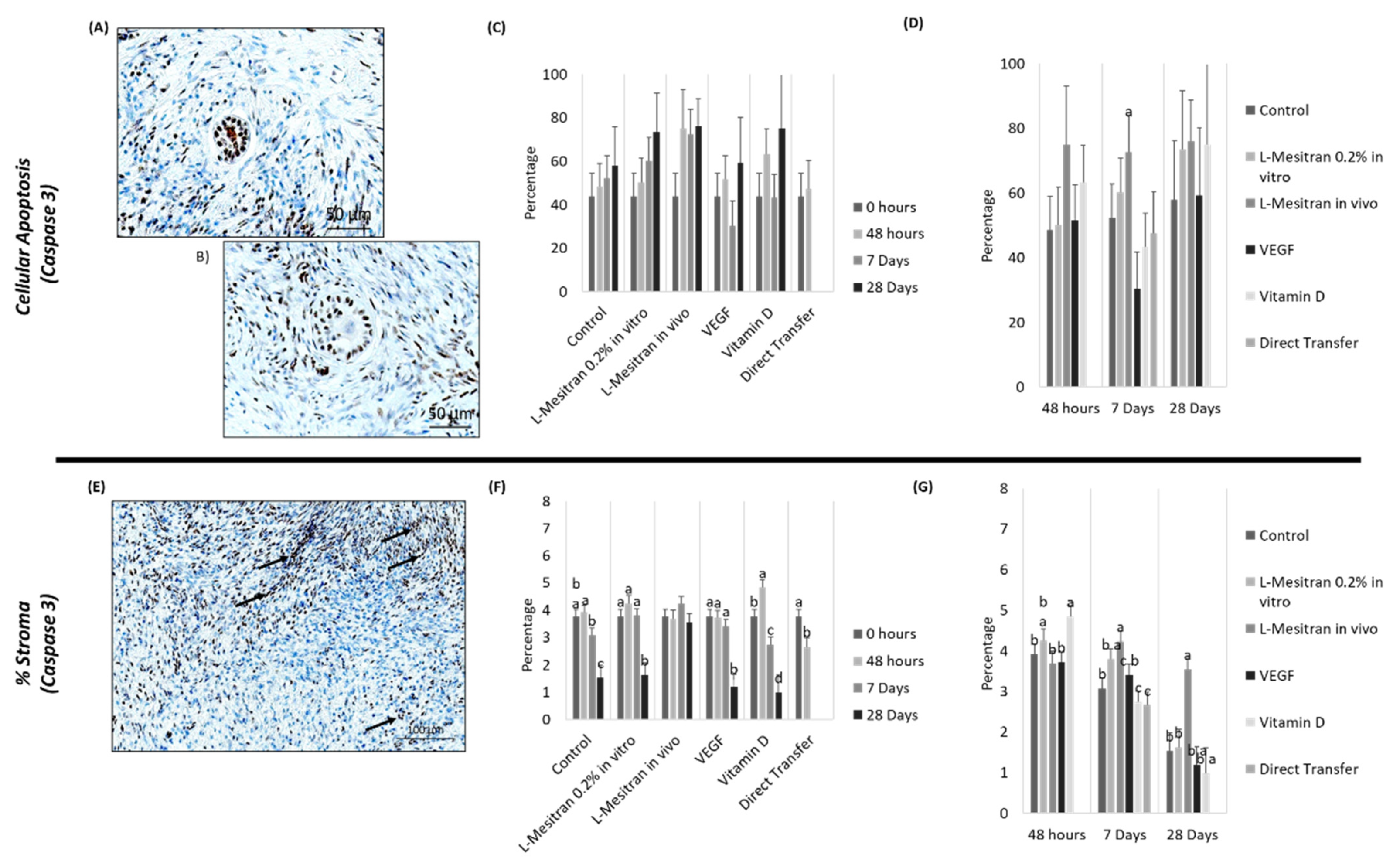
3.4. Cell Apoptosis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fisch, B.; Abir, R. Female fertility preservation: Past, present and future. Reproduction 2018, 156, F11–F27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ESHRE Guideline Group on Female Fertility Preservation; Anderson, R.A.; Amant, F.; Braat, D.; D’Angelo, A.; Chuva de Sousa Lopes, S.M.; Demeestere, I.; Dwek, S.; Frith, L.; Lambertini, M.; et al. ESHRE guideline: Female fertility preservation. Hum. Reprod. Open 2020, 2020, hoaa052. [Google Scholar] [CrossRef] [PubMed]
- Practice Committee of the American Society for Reproductive Medicine. Fertility preservation in patients undergoing gonadotoxic therapy or gonadectomy: A committee opinion. Fertil. Steril. 2019, 112, 1022–1033. [Google Scholar] [CrossRef] [PubMed]
- Donnez, J.; Dolmans, M.-M.; Pellicer, A.; Diaz-Garcia, C.; Sanchez Serrano, M.; Schmidt, K.T.; Ernst, E.; Luyckx, V.; Andersen, C.Y. Restoration of ovarian activity and pregnancy after transplantation of cryopreserved ovarian tissue: A review of 60 cases of reimplantation. Fertil. Steril. 2013, 99, 1503–1513. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ying, Y.F.; Ouyang, Y.L.; Wang, J.F.; Xu, J. VEGF and bFGF increase survival of xenografted human ovarian tissue in an experimental rabbit model. J. Assist. Reprod. Genet. 2013, 30, 1301–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donnez, J.; Cacciottola, L.; Dolmans, M.-M. Techniques for Ovarian Tissue Transplantation. In Female and Male Fertility Preservation; Grynberg, M., Patrizio, P., Eds.; Springer: Berlin/Heidelberg, Germany, 2022; pp. 163–174. ISBN 978-3-030-47767-7. [Google Scholar]
- Dolmans, M.-M.; Donnez, J. Fertility preservation in women for medical and social reasons: Oocytes vs ovarian tissue. Best Pract. Res. Clin. Obstet. Gynaecol. 2021, 70, 63–80. [Google Scholar] [CrossRef] [PubMed]
- Logsdon, E.A.; Finley, S.D.; Popel, A.S.; Mac Gabhann, F. A systems biology view of blood vessel growth and remodelling. J. Cell. Mol. Med. 2014, 18, 1491–1508. [Google Scholar] [CrossRef] [PubMed]
- Tahergorabi, Z.; Khazaei, M. A review on angiogenesis and its assays. Iran. J. Basic Med. Sci. 2012, 15, 1110–1126. Available online: https://pubmed.ncbi.nlm.nih.gov/23653839 (accessed on 1 June 2022).
- Rizov, M.; Andreeva, P.; Dimova, I. Molecular regulation and role of angiogenesis in reproduction. Taiwan. J. Obstet. Gynecol. 2017, 56, 127–132. [Google Scholar] [CrossRef]
- Denny, K.J.; Kelly, C.F.; Kumar, V.; Witham, K.L.; Cabrera, R.M.; Finnell, R.H.; Taylor, S.M.; Jeanes, A.; Woodruff, T.M. Autoantibodies against homocysteinylated protein in a mouse model of folate deficiency-induced neural tube defects. Birth. Defects Res. A Clin. Mol. Teratol. 2016, 106, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Bisht, M.; Dhasmana, D.C.; Bist, S.S. Angiogenesis: Future of pharmacological modulation. Indian J. Pharmacol. 2010, 42, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Rossiter, K.; Cooper, A.J.; Voegeli, D.; Lwaleed, B.A. Honey promotes angiogeneic activity in the rat aortic ring assay. J. Wound Care 2010, 19, 440–446. [Google Scholar] [CrossRef] [Green Version]
- Smaropoulos, E.; Cremers, N.A.J. Medical grade honey for the treatment of paediatric abdominal wounds: A case series. J. Wound Care 2020, 29, 94–99. [Google Scholar] [CrossRef]
- Pleeging, C.C.F.; Coenye, T.; Mossialos, D.; de Rooster, H.; Chrysostomou, D.; Wagener, F.A.D.T.G.; Cremers, N.A.J. Synergistic antimicrobial activity of supplemented medical-grade honey against pseudomonas aeruginosa biofilm formation and eradication. Antibiotics 2020, 9, 866. [Google Scholar] [CrossRef]
- Cheepa, F.F.; Liu, H.; Zhao, G. The Natural Cryoprotectant Honey for Fertility Cryopreservation. Bioengineering 2022, 9, 88. [Google Scholar] [CrossRef]
- Shahrokhi, S.Z.; Ghaffari, F.; Kazerouni, F. Role of vitamin D in female Reproduction. Clin. Chim. Acta 2016, 455, 33–38. [Google Scholar] [CrossRef]
- Xu, J.; Hennebold, J.D.; Seifer, D.B. Direct vitamin D3 actions on rhesus macaque follicles in three-dimensional culture: Assessment of follicle survival, growth, steroid, and antimüllerian hormone production. Fertil. Steril. 2016, 106, 1815–1820.e1. [Google Scholar] [CrossRef] [Green Version]
- Grundmann, M.; Haidar, M.; Placzko, S.; Niendorf, R.; Darashchonak, N.; Hubel, C.A.; von Versen-Höynck, F. Vitamin D improves the angiogenic properties of endothelial progenitor cells. Am. J. Physiol. Physiol. 2012, 303, C954–C962. [Google Scholar] [CrossRef]
- Nema, J.; Sundrani, D.; Joshi, S. Role of vitamin D in influencing angiogenesis in preeclampsia. Hypertens Pregnancy 2019, 38, 201–207. [Google Scholar] [CrossRef]
- Jamali, N.; Sorenson, C.M.; Sheibani, N. Vitamin D and regulation of vascular cell function. Am. J. Physiol. Heart Circ. Physiol. 2018, 314, H753–H765. [Google Scholar] [CrossRef] [Green Version]
- Aliashrafi, S.; Ebrahimi-Mameghani, M. A systematic review on vitamin D and angiogenesis. BMJ Open. 2017, 7 (Suppl. S1), A3–A4. [Google Scholar] [CrossRef]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Animal research: Reporting in vivo experiments: The ARRIVE guidelines. Br. J. Pharmacol. 2010, 160, 1577–1579. [Google Scholar] [CrossRef] [PubMed]
- Bols, P.E.J.; Aerts, J.M.J.; Langbeen, A.; Goovaerts, I.G.F.; Leroy, J.L.M.R. Xenotransplantation in immunodeficient mice to study ovarian follicular development in domestic animals. Theriogenology 2010, 73, 740–747. [Google Scholar] [CrossRef] [PubMed]
- Langbeen, A.; De, H.F.M.; Bartholomeus, E.; Leroy, J.L.M.R.; Bols, E.J. Bovine in vitro reproduction models can contribute to the development of (female) fertility preservation strategies. Theriogenology 2015, 84, 477–489. [Google Scholar] [CrossRef]
- Kagawa, N.; Silber, S.; Kuwayama, M. Successful vitrification of bovine and human ovarian tissue. Reprod. Biomed. Online 2009, 18, 568–577. [Google Scholar] [CrossRef]
- Aguiar, F.L.N.; Gastal, G.D.A.; Alves, K.A.; Alves, B.G.; Figueiredo, J.R.; Gastal, E.L. Supportive techniques to investigate in vitro culture and cryopreservation efficiencies of equine ovarian tissue: A review. Theriogenology 2020, 156, 296–309. [Google Scholar] [CrossRef]
- Rigalli, A.; Di Loreto, V.E. Experimental Surgical Models in the Laboratory Rat; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Lofgren, J.L.S.; Foley, P.L.; Golledge, H.D.R. Chapter 17—Anesthesia, Analgesia, and Euthanasia. In American College of Laboratory Animal Medicine; Suckow, M.A., Hankenson, F.C., Wilson, R.P., Foley, P., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 699–745. ISBN 978-0-12-814338-4. [Google Scholar]
- Shibutani, M. Chapter 26—Anesthesia, Artificial Ventilation and Perfusion Fixation. In Handbook of Experimental Animals; Krinke, G.J., Bullock, G., Bunton, T., Eds.; Academic Press: London, UK, 2000; pp. 511–521. ISBN 1874480X. [Google Scholar]
- Khajuria, D.K.; Razdan, R.; Mahapatra, D.R. Descrição de um novo método de ooforectomia em ratas. Rev. Bras. Reumatol. 2012, 52, 466–470. [Google Scholar] [CrossRef]
- Terren, C.; Bindels, J.; Nisolle, M.; Noël, A.; Munaut, C. Evaluation of an alternative heterotopic transplantation model for ovarian tissue to test pharmaceuticals improvements for fertility restoration. Reprod. Biol. Endocrinol. 2022, 20, 35. [Google Scholar] [CrossRef]
- de Macêdo, L.B.; Pimentel, M.M.L.; Dos Santos, F.A.; Bezerra, M.B.; Ladd, F.V.L.; de Moura, C.E.B. Equine chorionic gonadotrophin improved vascularization of feline ovarian tissue xenografted into immunosuppressed mice. Theriogenology 2018, 121, 78–81. [Google Scholar] [CrossRef]
- Olesen, H.Ø.; Pors, S.E.; Jensen, L.B.; Grønning, A.P.; Lemser, C.E.; Nguyen Heimbürger, M.T.H.; Mamsen, L.S.; Getreu, N.; Christensen, S.T.; Andersen, C.Y.; et al. N-acetylcysteine protects ovarian follicles from ischemia-reperfusion injury in xenotransplanted human ovarian tissue. Hum. Reprod. 2021, 36, 429–443. [Google Scholar] [CrossRef]
- American Society for Reproductive Medicine Ovarian tissue cryopreservation: A committee opinion. Fertil. Steril. 2014, 101, 1237–1243. [CrossRef]
- Dath, C.; Van Eyck, A.S.; Dolmans, M.M.; Romeu, L.; Delle Vigne, L.; Donnez, J.; Van Langendonckt, A. Xenotransplantation of human ovarian tissue to nude mice: Comparison between four grafting sites. Hum. Reprod. 2010, 25, 1734–1743. [Google Scholar] [CrossRef] [Green Version]
- Hormozi, M.; Talebi, S.; Khorram Khorshid, H.R.; Zarnani, A.-H.; Kamali, K.; Jeddi-Tehrani, M.; Soltangoraee, H.; Akhondi, M.M. The effect of Setarud (IMOD(TM)) on angiogenesis in transplanted human ovarian tissue to nude mice. Iran. J. Reprod. Med. 2015, 13, 605–614. [Google Scholar]
- Cha, S.K.; Shin, D.H.; Kim, B.Y.; Yoon, S.-Y.; Yoon, T.K.; Lee, W.S.; Chung, H.M.; Lee, D.R. Effect of Human Endothelial Progenitor Cell (EPC) or Mouse Vascular Endothelial Growth Factor-Derived Vessel Formation on the Survival of Vitrified/Warmed Mouse Ovarian Grafts. Reprod. Sci. 2014, 21, 859–868. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Ph, D.; Lee, H.; Sc, M.; Lee, C.; Sc, M.; Ko, S.; Sc, M. Assessment of vascular endothelial growth factor expression and apoptosis in the ovarian graft: Can exogenous gonadotropin promote angiogenesis after ovarian transplantation? Fertil. Steril. 2008, 90, 1550–1558. [Google Scholar] [CrossRef]
- Kong, H.S.; Lee, J.; Youm, H.W.; Kim, S.K.; Lee, J.R.; Suh, C.S.; Kim, S.H. Effect of treatment with angiopoietin-2 and vascular endothelial growth factor on the quality of xenografted bovine ovarian tissue in mice. PLoS ONE 2017, 12, e0184546. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.-M.; Yan, J.; Li, R.; Li, M.; Yan, L.-Y.; Wang, T.-R.; Zhao, H.-C.; Zhao, Y.; Yu, Y.; Qiao, J. Improvement in the quality of heterotopic allotransplanted mouse ovarian tissues with basic fibroblast growth factor and fibrin hydrogel. Hum. Reprod. 2013, 28, 2784–2793. [Google Scholar] [CrossRef] [Green Version]
- Gougeon, A. Dynamics of follicular growth in the human: A model from preliminary results. Hum Reprod. 1986, 1, 81–87. [Google Scholar] [CrossRef]
- Giatromanolaki, A.; Koukourakis, M.I.; Theodossiou, D.; Barbatis, K.; O’Byrne, K.; Harris, A.L.; Gatter, K.C. Comparative evaluation of angiogenesis assessment with anti-factor-VIII and anti-CD31 immunostaining in non-small cell lung cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 1997, 3, 2485–2492. [Google Scholar]
- Van de Goot, F.R.W.; Korkmaz, H.I.; Fronczek, J.; Witte, B.I.; Visser, R.; Ulrich, M.M.W.; Begieneman, M.P.V.; Rozendaal, L.; Krijnen, P.A.J.; Niessen, H.W.M. A new method to determine wound age in early vital skin injuries: A probability scoring system using expression levels of Fibronectin, CD62p and Factor VIII in wound hemorrhage. Forensic Sci. Int. 2014, 244, 128–135. [Google Scholar] [CrossRef]
- Wu, D.; Lei, Y.; Tong, Y.; Tang, F.; Qian, Y.; Zhou, Y. Angiogenesis of the frozen-thawed human fetal ovarian tissue at the early stage after xenotransplantation and the positive effect of Salviae miltiorrhizae. Anat. Rec. 2010, 293, 2154–2162. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Yin, T.; Yan, J.; Yan, L.; Jin, C.; Lu, C.; Wang, T.; Zhu, X.; Zhi, X.; Wang, J.; et al. Mesenchymal Stem Cells Enhance Angiogenesis and Follicle Survival in Human Cryopreserved Ovarian Cortex Transplantation. Cell Transplant. 2015, 24, 1999–2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takae, S.; Suzuki, N. Current state and future possibilities of ovarian tissue transplantation. Reprod. Med. Biol. 2019, 18, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, A.; Bag, S.; Banerjee, P.; Chatterjee, J. Wound healing efficacy of Jamun honey in diabetic mice model through reepithelialization, collagen deposition and angiogenesis. J. Tradit. Complement. Med. 2020, 10, 529–543. [Google Scholar] [CrossRef]
- Rossi, M.; Marrazzo, P. The Potential of Honeybee Products for Biomaterial Applications. Biomimetics 2021, 6, 6. [Google Scholar] [CrossRef]
- Yin, H.; Kristensen, S.G.; Jiang, H.; Rasmussen, A.; Andersen, C.Y. Survival and growth of isolated pre-antral follicles from human ovarian medulla tissue during long-term 3D culture. Hum. Reprod. 2016, 31, 1531–1539. [Google Scholar] [CrossRef]
- Redmer, D.A.; Reynolds, L.P. Angiogenesis in the ovary. Rev. Reprod. 1996, 1, 182–192. [Google Scholar] [CrossRef]
- Gastal, G.D.A.; Aguiar, F.L.N.; Ishak, G.M.; Cavinder, C.A.; Willard, S.T.; Ryan, P.L.; Feugang, J.M.; Gastal, E.L. Effect of cryopreservation techniques on proliferation and apoptosis of cultured equine ovarian tissue. Theriogenology 2019, 126, 88–94. [Google Scholar] [CrossRef]
- Nakajima, Y.; Kuranaga, E. Caspase-dependent non-apoptotic processes in development. Cell Death Differ. 2017, 24, 1422–1430. [Google Scholar] [CrossRef]
- Kinnear, H.M.; Tomaszewski, C.E.; Chang, F.L.; Moravek, M.B.; Xu, M.; Padmanabhan, V.; Shikanov, A. The ovarian stroma as a new frontier. Reproduction 2020, 160, R25–R39. [Google Scholar] [CrossRef]
- Lee, S.; Cho, H.-W.; Kim, B.; Lee, J.K.; Kim, T. The Effectiveness of Anti-Apoptotic Agents to Preserve Primordial Follicles and Prevent Tissue Damage during Ovarian Tissue Cryopreservation and Xenotransplantation. Int. J. Mol. Sci. 2021, 22, 2534. [Google Scholar] [CrossRef]
- Silber, S. Ovarian tissue cryopreservation and transplantation: Scientific implications. J. Assist. Reprod. Genet. 2016, 33, 1595–1603. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azevedo, A.R.; Pais, A.S.; Almeida-Santos, T.; Pires, V.M.R.; Pessa, P.; Marques, C.C.; Nolasco, S.; Castelo-Branco, P.; Prates, J.A.M.; Lopes-da-Costa, L.; et al. Medical Grade Honey as a Promising Treatment to Improve Ovarian Tissue Transplantation. Bioengineering 2022, 9, 357. https://doi.org/10.3390/bioengineering9080357
Azevedo AR, Pais AS, Almeida-Santos T, Pires VMR, Pessa P, Marques CC, Nolasco S, Castelo-Branco P, Prates JAM, Lopes-da-Costa L, et al. Medical Grade Honey as a Promising Treatment to Improve Ovarian Tissue Transplantation. Bioengineering. 2022; 9(8):357. https://doi.org/10.3390/bioengineering9080357
Chicago/Turabian StyleAzevedo, Ana Rita, Ana Sofia Pais, Teresa Almeida-Santos, Virgínia M. R. Pires, Pedro Pessa, Carla C. Marques, Sofia Nolasco, Pedro Castelo-Branco, José A. M. Prates, Luís Lopes-da-Costa, and et al. 2022. "Medical Grade Honey as a Promising Treatment to Improve Ovarian Tissue Transplantation" Bioengineering 9, no. 8: 357. https://doi.org/10.3390/bioengineering9080357
APA StyleAzevedo, A. R., Pais, A. S., Almeida-Santos, T., Pires, V. M. R., Pessa, P., Marques, C. C., Nolasco, S., Castelo-Branco, P., Prates, J. A. M., Lopes-da-Costa, L., Laranjo, M., Botelho, M. F., Pereira, R. M. L. N., & Pimenta, J. M. B. G. A. (2022). Medical Grade Honey as a Promising Treatment to Improve Ovarian Tissue Transplantation. Bioengineering, 9(8), 357. https://doi.org/10.3390/bioengineering9080357