
Simultaneous Maturation of Single Chain Antibody Stability and Affinity by CHO Cell Display


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Construction and Expression
2.2. Cell Culture
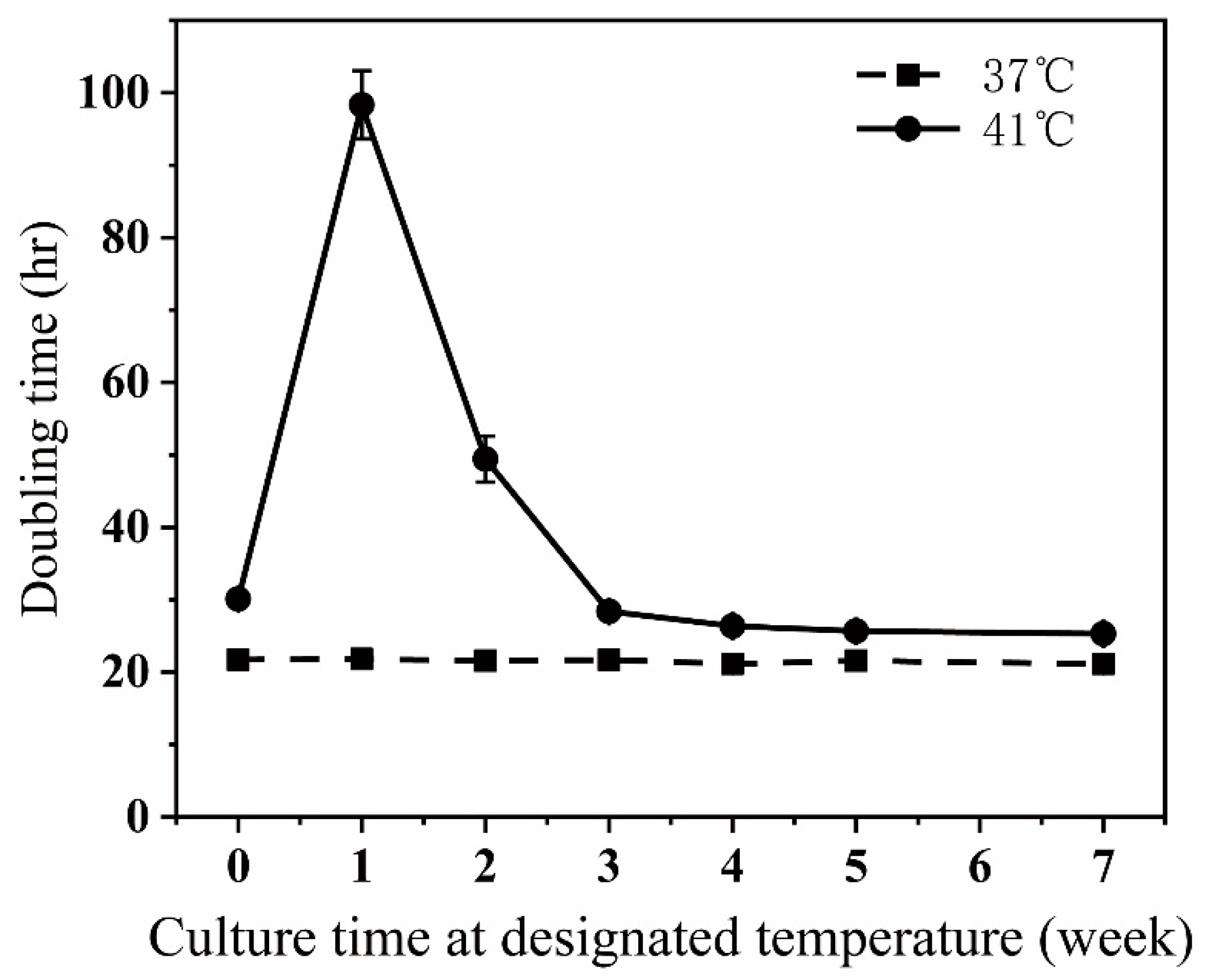
2.3. Thermo-Resistant Cells
2.4. Cell Survival Assays
2.5. Transfection and Antibody Affinity Maturation
2.6. Detection of Antibody Mutations
- CMV-F: CGCAAATGGGCGGTAGGCGTG;
- TM-R: CTGCGTGTCCTGGCCCACAGC.
2.7. Protein Expression and Purification
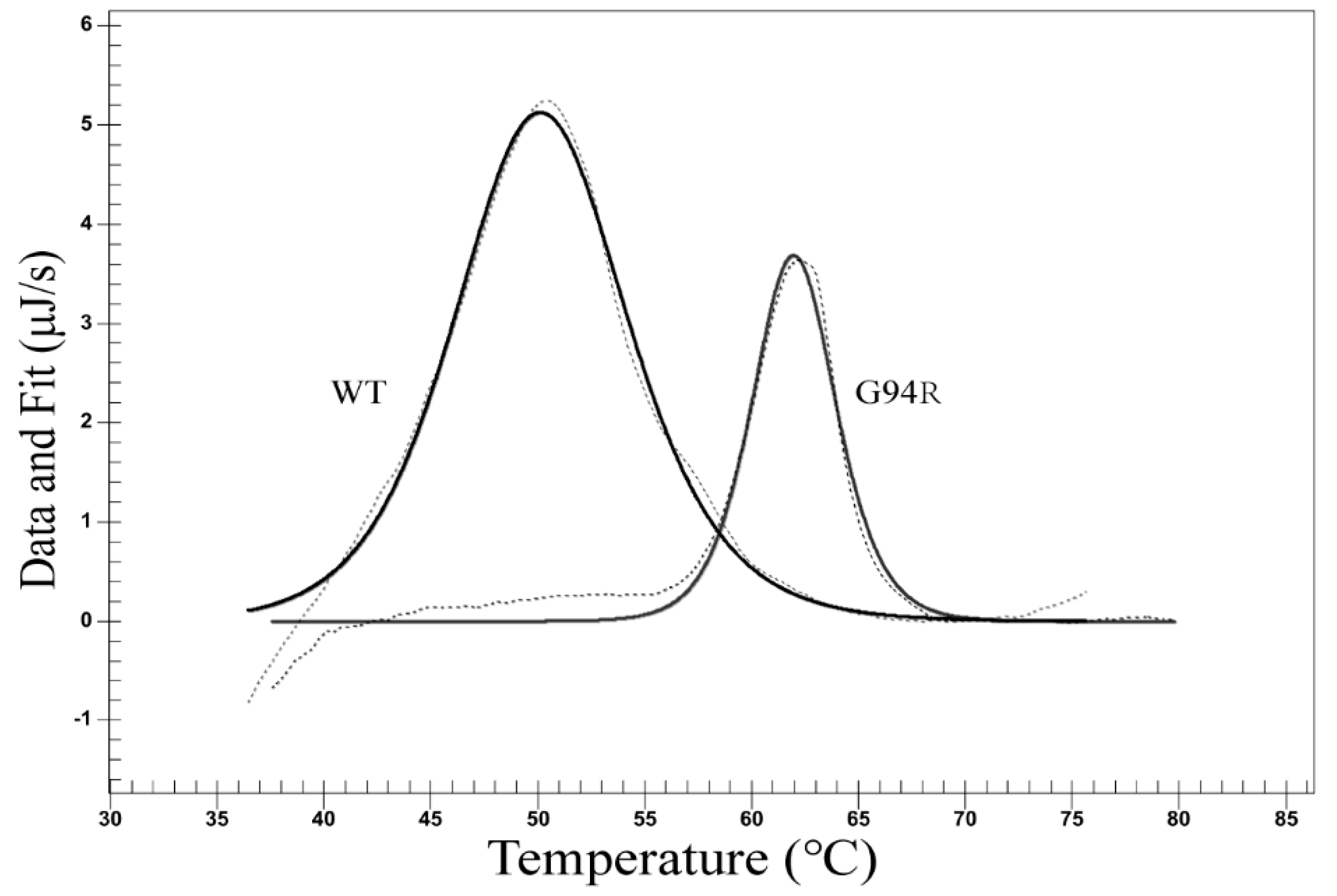
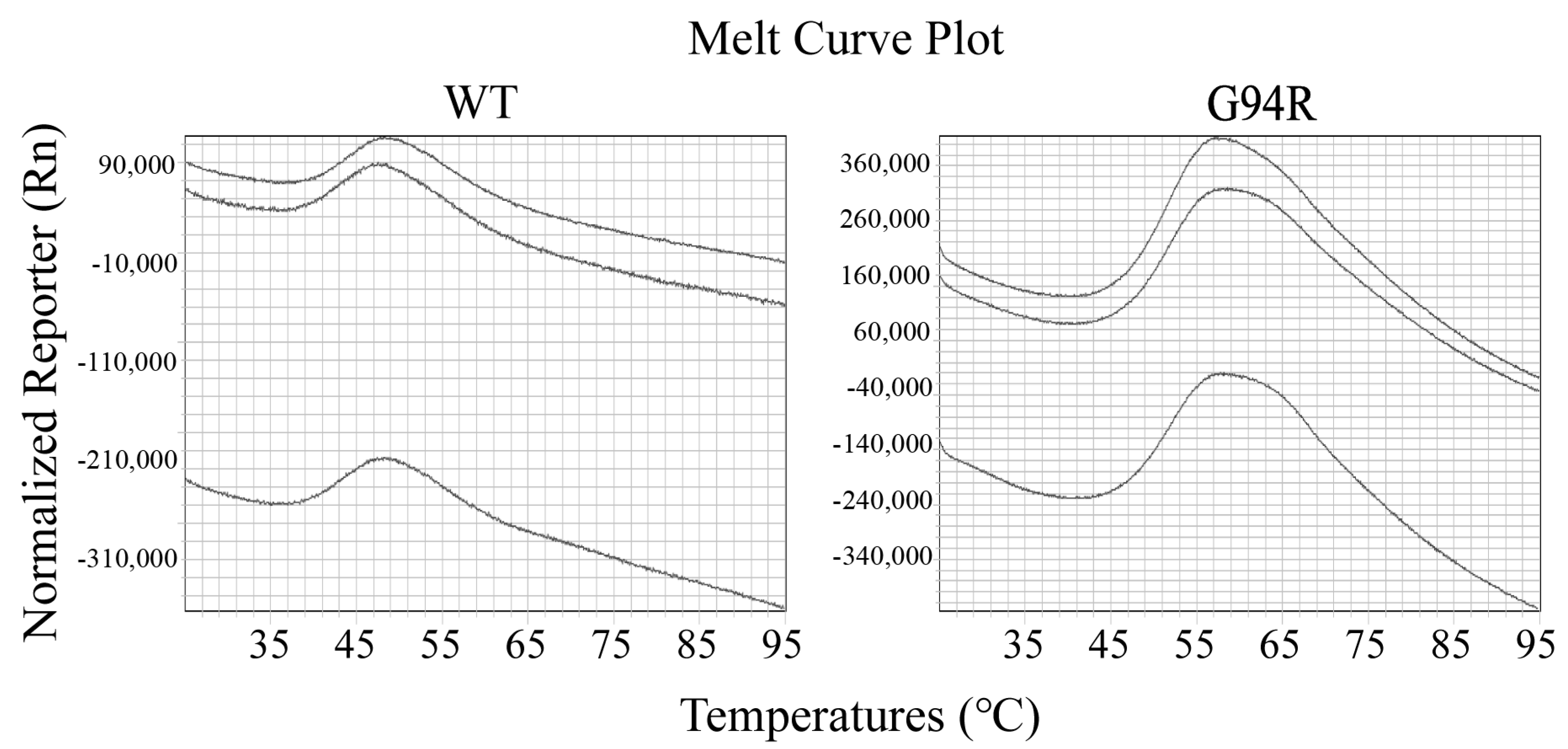
2.8. Antibody Thermal Stability Measurements
2.8.1. Differential Scanning Calorimetry (DSC)
2.8.2. Thermofluor Assay
2.8.3. Antibody Affinity Measurement
2.9. Analytical Ultracentrifugation
2.10. Maturation of Antibody for Improved Stability and Affinity
2.11. Prediction of the 3D Structure of the Mutation
3. Results
3.1. Generating Thermo-Resistant Cells
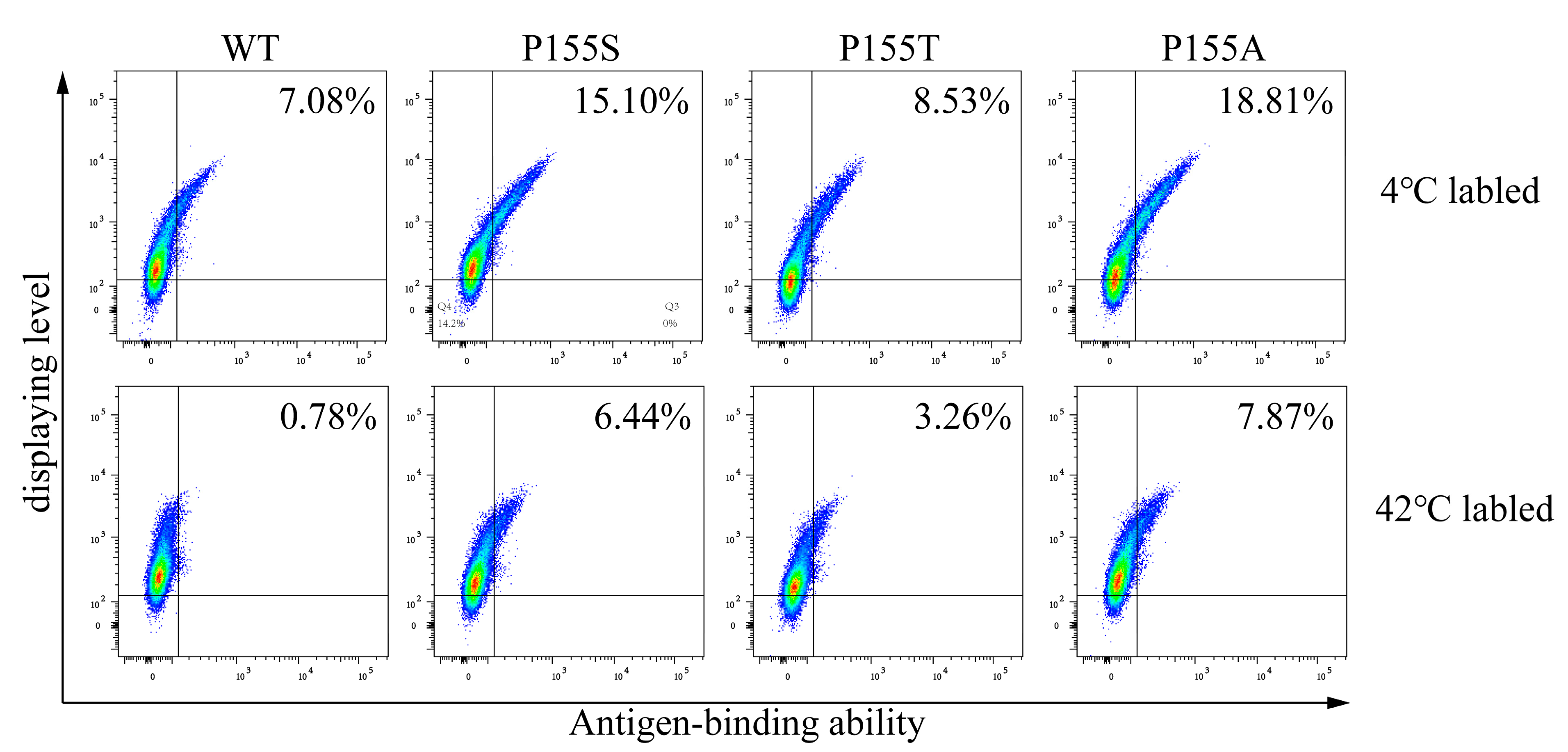
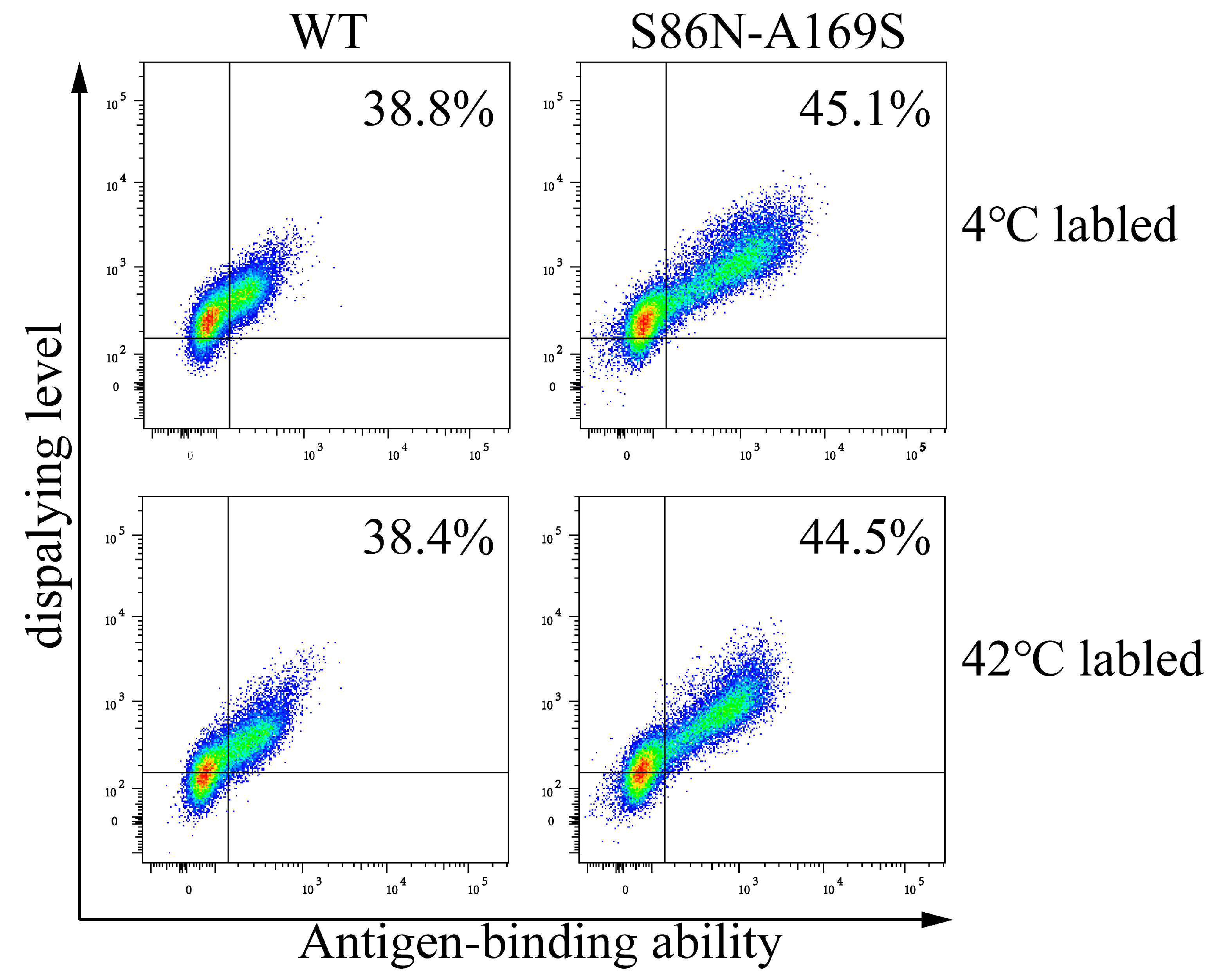
3.2. Conditions for Labeling Cells
3.3. Evolution to Simultaneously Improve Antibody’s Stability and Affinity
3.4. The Affinity and Stability Improvement of the Two Other Antibodies
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Baneyx, F.; Mujacic, M. Recombinant protein folding and misfolding in Escherichia coli. Nat. Biotechnol. 2004, 22, 1399–1408. [Google Scholar] [CrossRef] [PubMed]
- Kjeldsen, T.; Ludvigsen, S.; Diers, I.; Balschmidt, P.; Sørensen, A.R.; Kaarsholm, N.C. Engineering-enhanced Protein Secretory Expression in Yeast with Application to Insulin. J. Biol. Chem. 2002, 277, 18245–18248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, D.; Dudgeon, K.; Rouet, R.; Schofield, P.; Jermutus, L.; Christ, D. Aggregation, stability, and formulation of human antibody therapeutics. Adv. Protein Chem. Struct. Biol. 2011, 84, 41–61. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, F.R. Recombinant expression systems in the pharmaceutical industry. Appl. Microbiol. Biotechnol. 2004, 65, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Shusta, E.V.; Kieke, M.C.; Parke, E.; Kranz, D.M.; Wittrup, K. Yeast polypeptide fusion surface display levels predict thermal stability and soluble secretion efficiency. J. Mol. Biol. 1999, 292, 949–956. [Google Scholar] [CrossRef]
- Wang, X.X.; Shusta, E.V. The use of scFv-displaying yeast in mammalian cell surface selections. J. Immunol. Methods 2005, 304, 30–42. [Google Scholar] [CrossRef]
- Wang, F.; Sen, S.; Zhang, Y.; Ahmad, I.; Zhu, X.; Wilson, I.A.; Smider, V.V.; Magliery, T.J.; Schultz, P.G. Somatic hypermutation maintains antibody thermodynamic stability during affinity maturation. Proc. Natl. Acad. Sci. USA 2013, 110, 4261–4266. [Google Scholar] [CrossRef] [Green Version]
- Franklin, E.; Cunningham, O.; Fennell, B. Parallel Evolution of Antibody Affinity and Thermal Stability for Optimal Biotherapeutic Development. Methods Mol. Biol. 2018, 1827, 457–477. [Google Scholar] [CrossRef] [PubMed]
- Lombana, T.N.; Dillon, M.; Iii, J.B.; Spiess, C. Optimizing antibody expression by using the naturally occurring framework diversity in a live bacterial antibody display system. Sci. Rep. 2015, 5, 17488. [Google Scholar] [CrossRef] [Green Version]
- Ho, M.; Pastan, I. Mammalian Cell Display for Antibody Engineering. Methods Mol. Biol. 2009, 525, 337–352. [Google Scholar] [CrossRef] [Green Version]
- McConnell, A.D.; Spasojevich, V.; Macomber, J.L.; Krapf, I.P.; Chen, A.; Sheffer, J.C.; Berkebile, A.; Horlick, R.A.; Neben, S.; King, D.J.; et al. An integrated approach to extreme thermostabilization and affinity maturation of an antibody. Protein Eng. Des. Sel. 2013, 26, 151–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Qiu, J.; Chen, C.; Liu, C.; Liu, Y.; An, L.; Jia, J.; Tang, J.; Wu, L.; Hang, H. Affinity maturation of anti-TNF-alpha scFv with somatic hypermutation in non-B cells. Protein Cell 2012, 3, 460–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Li, N.; Zhao, Y.; Hang, H. Coupling recombinase-mediated cassette exchange with somatic hypermutation for antibody affinity maturation in CHO cells. Biotechnol. Bioeng. 2016, 113, 39–51. [Google Scholar] [CrossRef]
- Zacharias, D.A.; Violin, J.D.; Newton, A.C.; Tsien, R.Y. Partitioning of lipid-modified monomeric GFPs into membrane microdomains of live cells. Science 2002, 296, 913–916. [Google Scholar] [CrossRef] [Green Version]
- Leem, J.; Dunbar, J.; Georges, G.; Shi, J.; Deane, C.M. ABodyBuilder: Automated antibody structure prediction with data–driven accuracy estimation. MAbs 2016, 8, 1259–1268. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Song, L.; Miao, Z.; Hu, Y.; Tian, L.; Jiang, T. Improved side-chain modeling by coupling clash-detection guided iterative search with rotamer relaxation. Bioinformatics 2011, 27, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Dewey, W.C.; Hopwood, L.E.; Sapareto, S.A.; Gerweck, L.E. Cellular Responses to Combinations of Hyperthermia and Radiation. Radiology 1977, 123, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.; Nagata, S.; Pastan, I. Isolation of anti-CD22 Fv with high affinity by Fv display on human cells. Proc. Natl. Acad. Sci. USA 2006, 103, 9637–9642. [Google Scholar] [CrossRef] [Green Version]
- Ghaderi, D.; Zhang, M.; Hurtado-Ziola, N.; Varki, A. Production platforms for biotherapeutic glycoproteins. Occur-rence, impact, and challenges of non-human sialylation. Biotechnol. Genet. Eng. Rev. 2012, 28, 147–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingram, J.R.; Blomberg, O.S.; Rashidian, M.; Ali, L.; Garforth, S.; Fedorov, E.; Fedorov, A.A.; Bonanno, J.B.; Le Gall, C.; Crowley, S.; et al. Anti–CTLA-4 therapy requires an Fc domain for efficacy. Proc. Natl. Acad. Sci. USA 2018, 115, 3912–3917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, T.; Yang, Y.; Ng, S.K. Advances in Mammalian Cell Line Development Technologies for Recombinant Protein Production. Pharmaceuticals 2013, 6, 579–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traxlmayr, M.W.; Obinger, C. Directed evolution of proteins for increased stability and expression using yeast display. Arch. Biochem. Biophys. 2012, 526, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Bowers, P.M.; Horlick, R.A.; Neben, T.Y.; Toobian, R.M.; Tomlinson, G.L.; Dalton, J.L.; Jones, H.A.; Chen, A.; Altobell, L., 3rd; Zhang, X.; et al. Coupling mammalian cell surface display with somatic hypermutation for the discovery and maturation of human antibodies. Proc. Natl. Acad. Sci. USA 2011, 108, 20455–20460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maute, R.L.; Gordon, S.R.; Mayer, A.T.; McCracken, M.N.; Natarajan, A.; Ring, N.G.; Kimura, R.; Tsai, J.M.; Manglik, A.; Kruse, A.C.; et al. Engineering high-affinity PD-1 variants for optimized immunotherapy and immuno-PET imaging. Proc. Natl. Acad. Sci. USA 2015, 112, E6506–E6514. [Google Scholar] [CrossRef] [Green Version]
- Luo, R.; Zhao, Y.; Fan, Y.; An, L.; Jiang, T.; Ma, S.; Hang, H. High efficiency CHO cell display-based antibody matura-tion. Sci. Rep. 2020, 10, 8102. [Google Scholar] [CrossRef] [PubMed]
- Robertson, N.; Lopez-Anton, N.; Gurjar, S.A.; Khalique, H.; Khalaf, Z.; Clerkin, S.; Leydon, V.R.; Parker-Manuel, R.; Raeside, A.; Payne, T.; et al. Development of a novel mammalian display system for selection of antibodies against membrane proteins. J. Biol. Chem. 2020, 295, 18436–18448. [Google Scholar] [CrossRef]
- See, K.; Kadonosono, T.; Miyamoto, K.; Tsubaki, T.; Ota, Y.; Katsumi, M.; Ryo, S.; Aida, K.; Minegishi, M.; Isozaki, T.; et al. Antibody-guided design and identification of CD25-binding small an-tibody mimetics using mammalian cell surface display. Sci. Rep. 2021, 11, 22098. [Google Scholar] [CrossRef]
- Holger, T.; Lars, T.; Jiandong, L.; Reinhard, V.W.; Gunther, B.; Thomas, S.; Isabel Tourais, E.; Behrens, C.K.; Bénédict, F.; Nathalie, F. Rise and fall of an anti-MUC1 specific antibody. PLoS ONE 2011, 6, e15921. [Google Scholar]
- Liu, B.; Conrad, F.; Roth, A.; Drummond, D.C.; Simko, J.P.; Marks, J.D. Recombinant full-length human IgG1s targeting hormone-refractory prostate cancer. J. Mol. Med. 2007, 85, 1113. [Google Scholar] [CrossRef]
- Steinwand, M.; Droste, P.; Frenzel, A.; Hust, M.; Dübel, S.; Schirrmann, T. The influence of antibody fragment format on phage display based affinity maturation of IgG. MAbs 2014, 6, 204–218. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; An, L.; Zhao, Y.; Zhang, C.; Li, S.; Ye, C.; Jing, S.; Hang, H. In vitro affinity maturation of antibody against membrane-bound GPCR molecules. Appl. Microbiol. Biotechnol. 2019, 103, 7703–7717. [Google Scholar] [CrossRef] [PubMed] [Green Version]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clone (Base) | Clone (Amino Acid) | KD (M) | Koff (1/s) | Kon (1/MS) |
|---|---|---|---|---|
| WT | No | 1.553 × 10−6 | 8.727 × 10−3 | 5.618 × 103 |
| G316A | G94R | 9.688 × 10−9 | 1.586 × 10−3 | 1.637 × 105 |
| Clone (Base) | Clone (Amino Acid) | KD (M) | Koff (1/s) | Kon (1/MS) |
|---|---|---|---|---|
| WT | WT | 9.499 × 10−10 | 7.120 × 10−4 | 7.496 × 105 |
| C463T | P155S | 2.857 × 10−10 | 1.860 × 10−4 | 6.512 × 105 |
| C463G | P155A | 1.874 × 10−10 | 1.290 × 10−4 | 6.869 × 105 |
| C463A | P155T | 6.769 × 10−11 | 5.680 × 10−5 | 8.383 × 105 |
| Mutation | ΔΔG |
|---|---|
| P155A | −0.19 |
| P155S | −0.56 |
| P155T | −0.71 |
| Clone (Base) | Clone (Amino Acid) | KD (M) | Koff (1/s) | Kon (1/MS) |
|---|---|---|---|---|
| WT | WT | 1.876 × 10−8 | 2.288 × 10−3 | 1.220 × 105 |
| G257A-G505T | S86N-A169S | 5.287 × 10−9 | 7.352 × 10−4 | 1.391 × 105 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, R.; Qu, B.; An, L.; Zhao, Y.; Cao, Y.; Ren, P.; Hang, H. Simultaneous Maturation of Single Chain Antibody Stability and Affinity by CHO Cell Display. Bioengineering 2022, 9, 360. https://doi.org/10.3390/bioengineering9080360
Luo R, Qu B, An L, Zhao Y, Cao Y, Ren P, Hang H. Simultaneous Maturation of Single Chain Antibody Stability and Affinity by CHO Cell Display. Bioengineering. 2022; 9(8):360. https://doi.org/10.3390/bioengineering9080360
Chicago/Turabian StyleLuo, Ruiqi, Baole Qu, Lili An, Yun Zhao, Yang Cao, Peng Ren, and Haiying Hang. 2022. "Simultaneous Maturation of Single Chain Antibody Stability and Affinity by CHO Cell Display" Bioengineering 9, no. 8: 360. https://doi.org/10.3390/bioengineering9080360
APA StyleLuo, R., Qu, B., An, L., Zhao, Y., Cao, Y., Ren, P., & Hang, H. (2022). Simultaneous Maturation of Single Chain Antibody Stability and Affinity by CHO Cell Display. Bioengineering, 9(8), 360. https://doi.org/10.3390/bioengineering9080360