
Biodiversity of Terrestrial Testate Amoebae in Western Siberia Lowland Peatlands

 , , , , , , ,
, , , , , , , 
Abstract
:1. Summary
2. Data Description
2.1. Dataset Description
2.2. Species Diversity and Community Structure in the Dataset
3. Methods
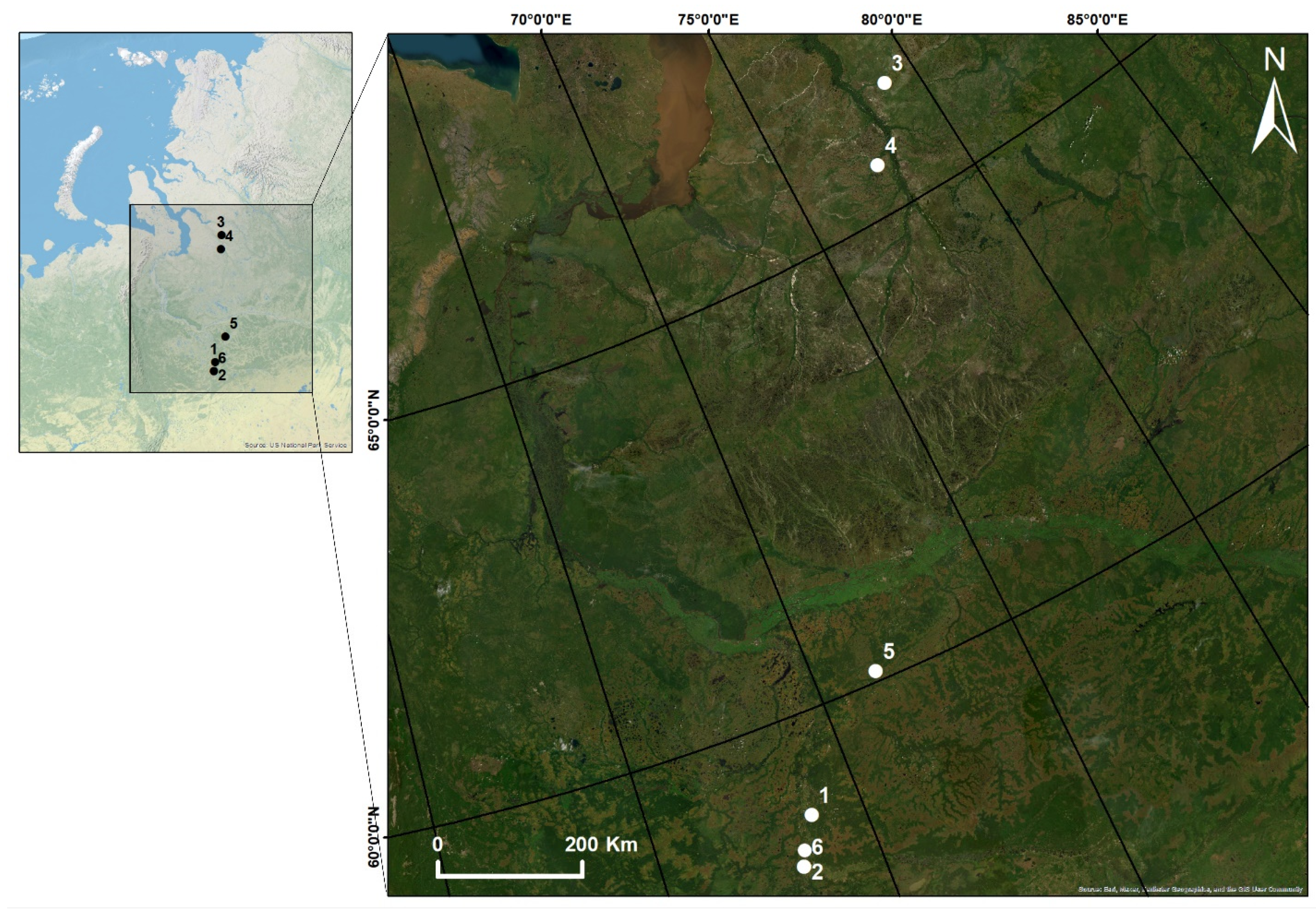
3.1. Study Area
3.2. Sample Collection and Treatment
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Adl, S.M.; Simpson, A.G.; Lane, C.E.; Lukeš, J.; Bass, D.; Bowser, S.S.; Brown, M.W.; Burki, F.; Dunthorn, M.; Hampl, V.; et al. The revised classification of eukaryotes. J. Eukaryot. Microbiol. 2012, 59, 429–514. [Google Scholar] [CrossRef] [PubMed]
- Kosakyan, A.; Gomaa, F.; Lara, E.; Lahr, D.J. Current and future perspectives on the systematics, taxonomy and nomenclature of testate amoebae. Eur. J. Protistol. 2016, 55, 105–117. [Google Scholar] [CrossRef] [PubMed]
- González-Miguéns, R.; Soler-Zamora, C.; Villar-Depablo, M.; Todorov, M.; Lara, E. Multiple convergences in the evolutionary history of the testate amoeba family Arcellidae (Amoebozoa: Arcellinida: Sphaerothecina): When the ecology rules the morphology. Zool. J. Linn. Soc. 2021, 194, 1044–1071. [Google Scholar] [CrossRef]
- Kosakyan, A.; Siemansma, F.; Fernández, L.D.; Burdman, L.; Krashevska, V.; Lara, E. Amoebae. In Keys to Neotropical and Antartic Fauna—Thorp and Covich’s Freshwater Invertebrates; Academic Press: Cambridge, UK, 2020; pp. 13–37. [Google Scholar]
- Gilbert, D.; Mitchell, E.A.D. Chapter 13 Microbial Diversity in Sphagnum Peatlands. Dev. Earth Surf. Process. 2006, 9, 287–318. [Google Scholar]
- Gilbert, D.; Amblard, C.; Bourdier, G.; Francez, A.-J. The Microbial Loop at the Surface of a Peatland: Structure, Function, and Impact of Nutrient Input. Microb. Ecol. 1998, 35, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Potapov, A.M.; Rozanova, O.L.; Semenina, E.E.; Leonov, V.D.; Belyakova, O.I.; Bogatyreva, V.Y.; Degtyarev, M.I.; Esaulov, A.S.; Korotkevich, A.Y.; Kudrin, A.A.; et al. Size Compartmentalization of Energy Channeling in Terrestrial Belowground Food Webs. Ecology 2021, 102, e03421. [Google Scholar] [CrossRef]
- Payne, R.J. Seven Reasons Why Protists Make Useful Bioindicators. Acta Protozool. 2013, 52, 105–113. [Google Scholar]
- Amesbury, M.J.; Swindles, G.T.; Bobrov, A.; Charman, D.J.; Holden, J.; Lamentowicz, M.; Mallon, G.; Mazei, Y.; Mitchell EA, D.; Payne, R.J.; et al. Development of a new pan-European testate amoeba transfer function for reconstructing peatland palaeohydrology. Quat. Sci Rev. 2016, 152, 132–151. [Google Scholar] [CrossRef]
- Mazei, Y.u.A.; Tsyganov, A.N.; Bubnova, O.A. Structure of a community of testate amoebae in a sphagnum dominated bog in upper Sura flow (Middle Volga Territory). Biol. Bull. 2007, 34, 382–394. [Google Scholar] [CrossRef]
- Booth, R.; Sullivan, M.; Sousa, V. Ecology of testate amoebae in a North Carolina pocosin and their potential use as environmental and paleoenvironmental indicators. Ecoscience 2008, 15, 277–289. [Google Scholar] [CrossRef]
- Portsmuth, A.; Avel-Niinemets, E.; Pensa, M. Distribution of testate amoebae along a gradient hummock-lawn-hollow in a Sphagnum-bog: Potential implications for palaeoecological reconstructions. Pol. J. Ecol. 2011, 59, 551–566. [Google Scholar]
- Song, L.; Li, H.; Wang, K.; Wu, D.; Wu, H. Ecology of testate amoebae and their potential use as palaeohydrologic indicators from peatland in Sanjiang Plain, Northeast China. Front. Earth Sci. 2014, 8, 564–572. [Google Scholar] [CrossRef]
- Tsyganov, A.N.; Babeshko, K.V.; Novenko, E.Y.; Malysheva, E.A.; Payne, R.J.; Mazei, Y.A. Quantitative reconstruction of peatland hydrological regime with fossil testate amoebae communities. Russ. J. Ecol. 2017, 48, 191–198. [Google Scholar] [CrossRef]
- Krashevska, V.; Tsyganov, A.; Esaulov, A.; Mazei, Y.; Hapsari, K.A.; Saad, A.; Sabiham, S.; Behling, H.; Biagioni, S. Testate Amoeba Species-and Trait-Based Transfer Functions for Reconstruction of Hydrological Regime in Tropical Peatland of Central Sumatra, Indonesia. Front. Ecol. 2020, 8, 225. [Google Scholar] [CrossRef]
- Kur’ina, I.V.; Preis, Y.I.; Bobrov, A.A. Testate amoebae inhabiting middle taiga bogs in Western Siberia. Biol. Bull. 2010, 37, 357–362. [Google Scholar] [CrossRef]
- Mazei, Y.A.; Chernyshov, V.A. Testate amoebae communities in the southern tundra and forest-tundra of Western. Sib. Biol. Bull. 2011, 38, 789–796. [Google Scholar] [CrossRef]
- Tsyganov, A.N.; Zarov, E.A.; Mazei, Y.u.A.; Kulkov, M.G.; Babeshko, K.V.; Yushkovets, S.Y.; Payne, R.J.; Ratcliffe, J.L.; Fatynina, Y.u.A.; Zazovskaya, E.; et al. Key periods of peatland development and environmental changes in the middle taiga zone of Western Siberia during the Holocene. Ambio J. Hum. Environ. 2021, 50, 1896–1909. [Google Scholar] [CrossRef]
- Qin, Y.; Li, H.; Mazei, Y.; Kurina, I.; Swindles, G.T.; Bobrov, A.; Tsyganov, A.N.; Gu, Y.; Huang, X.; Xue, J.; et al. Developing a continental-scale testate amoeba hydrological transfer function for Asian peatlands. Quat. Sci. Rev. 2021, 258, 106868. [Google Scholar] [CrossRef]
- Payne, R. The standard preparation method for testate amoebae leads to selective loss of the smallest taxa. Quat. Newsl. 2009, 119, 16–20. [Google Scholar]
- Kurina, I.V.; Golovatskaya, E.A. Testate Amoebae Assemblages (Rhizopoda and Testacea) in the Peat Deposits of the Floodplain Terrace Swamp (the South of Forested Zone of Western Siberia). Biol. Bull. 2018, 45, 91–99. [Google Scholar] [CrossRef]
- Halaś, A.; Lamentowicz, M.; Łuców, D.; Słowiński, M. Developing a new testate amoeba hydrological transfer function for permafrost peatlands of NW Siberia. Quat. Sci. Rev. 2023, 308, 108067. [Google Scholar] [CrossRef]
- Mazei, Y.; Chernyshov, V.; Bukhkalo, S.; Mazei, N.; Creevy, A.L.; Payne, R. Exploring the diversity and ecology of testate amoebae in West Siberian peatlands. Acta Protozool. 2017, 56, 59–70. [Google Scholar]
- Occurrence Data: GBIF IPT User Manual. Available online: https://ipt.gbif.org/manual/en/ipt/latest/occurrence-data (accessed on 23 August 2023).
- List of Darwin Core Terms. Available online: https://dwc.tdwg.org/list/ (accessed on 23 August 2023).
- Mazei, Y.; Chernyshov, V.; Tsyganov, A.N.; Payne, R.J. Testing the Effect of Refrigerated Storage on Testate Amoeba Samples. Microb. Ecol. 2015, 70, 861–864. [Google Scholar] [CrossRef] [PubMed]
- Avel, E.; Pensa, M. Preparation of testate amoebae samples affects water table depth reconstructions in peatland palaeoecological studies. Est. J. Earth Sci. 2013, 62, 113–119. [Google Scholar] [CrossRef]
- Mazei, Y.; Tsyganov, A.N. Freshwater Testate Amoebae; KMK: Moscow, Russia, 2006. [Google Scholar]
- Tsyganov, A.N.; Babeshko, K.V.; Mazei, Y.A. A Guide to Testate Amoebae with the Keys to Genera; Penza University Press: Penza, Russia, 2016; 132p. [Google Scholar]
- González-Miguéns, R.; Todorov, M.; Blandenier, Q.; Duckert, C.; Porfirio-Sousa, A.L.; Ribeiro, G.M.; Ramos, D.; Lahr, D.J.G.; Buckley, D.; Lara, E. Deconstructing Difflugia: The tangled evolution of lobose testate amoebae shells (Amoebozoa: Arcellinida) illustrates the importance of convergent evolution in protist phylogeny. Mol. Phylogenet. Evol. 2022, 175, 107557. [Google Scholar] [CrossRef]


{kind=link}
| Column Label | Column Description |
|---|---|
| eventID | An identifier for the set of information associated with an Event |
| occurrenceID | An identifier for the occurrence (as opposed to a particular digital record of the occurrence) |
| basisOfRecord | The specific nature of the data record |
| eventDate | The date when material was collected or the sampling period |
| Kingdom | The full scientific name of the Kingdom in which the taxon is classified |
| scientificName | The full scientific name, including the genus name and the lowest level of taxonomic rank with the authority |
| Family | The full scientific name of the Family in which the taxon is classified |
| Class | The full scientific name of the Class in which the taxon is classified |
| taxonRank | The taxonomic rank of the most specific name in the scientificName |
| decimalLatitude | The geographic latitude of a location in decimal degrees |
| decimalLongitude | The geographic longitude of a location in decimal degrees |
| countryCode | The standard code for the country in which the location is found |
| individualCount | The number of individuals present at the time of the occurrence |
| organismQuantity | A number or enumeration value for the quantity of organisms (counted shells) |
| organismQuantityType | The type of quantification system used for the quantity of organisms |
| measurementType | The nature of the measurement, fact, characteristic, or assertion (substrate moisture) |
| measurementUnit | The units associated with the dwc:measurementValue (%) |
| measurementValue | The value of the measurement, fact, characteristic, or assertion |
| locationRemarks | Comments or notes about the dcterms:Location (microtopography) |
| Families | Number of Genera | Number of Species | Number of Occurrences |
|---|---|---|---|
| Amphitremidae Poche, 1913 | 1 | 1 | 44 |
| Arcellidae Ehrenberg, 1843 | 2 | 9 | 131 |
| Assulinidae Lara et al., 2007 | 2 | 3 | 180 |
| Centropyxidae Jung, 1942 | 4 | 15 | 284 |
| Cryptodifflugiidae Jung, 1942 | 1 | 1 | 46 |
| Difflugiidae Wallich, 1864 | 1 | 3 | 50 |
| Euglyphidae Wallich, 1864, emend. Lara et al., 2007 | 2 | 16 | 489 |
| Heleoperidae Jung, 1942 | 1 | 2 | 20 |
| Hyalospheniidae Schultze, 1877 emend. Kosakyan and Lara, 2012 | 6 | 11 | 177 |
| Incertae sedis (Class: Tubulinea) | 5 | 7 | 109 |
| Lesquereusiidae Jung, 1942 | 2 | 3 | 48 |
| Netzeliidae Kosakyan et al., 2016, emend. Gonzales-Miguens et al., 2021 | 2 | 5 | 49 |
| Phryganellidae Jung, 1942 | 1 | 2 | 84 |
| Sphenoderiidae Chatelain et al., 2013 | 1 | 2 | 9 |
| Trinematidae Hoogenraad & De Groot, 1940, emend Adl et al., 2012 | 2 | 8 | 461 |
| Total | 32 | 88 | 2181 |
| Taxa | Study Area | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | |||||||
| ab. | occ. | ab. | occ. | ab. | occ. | ab. | occ. | ab. | occ. | ab. | occ. | |
| Alabasta longicollis (Penard 1890) Duckert, Blandenier, Kosakyan & Singer 2018 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.04 | 1 | 0.45 | 2 | 0.00 | 0 |
| Alabasta militaris (Penard 1890) Duckert, Blandenier, Kosakyan & Singer 2018 | 2.46 | 10 | 0.67 | 1 | 1.18 | 8 | 0.14 | 8 | 0.22 | 1 | 0.00 | 0 |
| Arcella gibbosa Penard, 1890 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.02 | 1 | 0.00 | 0 | 0.30 | 5 |
| Arcella hemisphaerica Perty, 1852 | 0.00 | 0 | 0.92 | 1 | 0.00 | 0 | 0.11 | 4 | 0.00 | 0 | 0.75 | 7 |
| Arcella rotundata Playfair, 1918 | 0.76 | 1 | 3.91 | 2 | 0.00 | 0 | 0.11 | 4 | 8.91 | 6 | 1.14 | 7 |
| Archerella flavum (Archer, 1877) Loeblich and Tappan, 1961 | 0.00 | 0 | 1.58 | 3 | 0.59 | 2 | 0.61 | 17 | 2.45 | 5 | 2.16 | 17 |
| Assulina muscorum Greeff, 1888 | 7.19 | 16 | 11.4 | 10 | 5.84 | 10 | 5.71 | 45 | 5.01 | 7 | 6.25 | 30 |
| Assulina seminulum Ehrenberg, 1848 | 0.12 | 2 | 0.08 | 1 | 7.74 | 10 | 1.58 | 26 | 0.11 | 1 | 0.00 | 0 |
| Awerintzewia cyclostoma Schouteden, 1906 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.02 | 1 | 0.00 | 0 | 0.33 | 7 |
| Bullinularia indica (Penard, 1907) Deflandre, 1953 | 0.76 | 5 | 0.58 | 3 | 2.95 | 11 | 0.91 | 28 | 0.33 | 2 | 2.31 | 17 |
| Centropyxis aculeata (Ehrenberg, 1838) Stein, 1859 | 0.06 | 1 | 0.00 | 0 | 0.00 | 0 | 0.04 | 2 | 0.00 | 0 | 0.00 | 0 |
| Centropyxis aculeata oblonga Deflandre, 1929 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.99 | 6 |
| Centropyxis aerophila Deflandre, 1929 | 0.76 | 4 | 0.00 | 0 | 3.28 | 10 | 11.38 | 50 | 0.78 | 1 | 1.23 | 9 |
| Centropyxis aerophila sphangicola Deflandre, 1929 | 3.04 | 8 | 2.41 | 2 | 0.07 | 1 | 3.08 | 21 | 0.33 | 2 | 0.12 | 3 |
| Centropyxis cassis (Wallich, 1864) Deflandre, 1929 | 0.12 | 1 | 0.00 | 0 | 3.41 | 6 | 0.21 | 6 | 0.00 | 0 | 0.00 | 0 |
| Centropyxis constricta (Ehrenberg, 1841) Penard, 1890 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.03 | 1 |
| Centropyxis ecornis (Ehrenberg, 1841) Leidy, 1879 | 0.00 | 0 | 0.00 | 0 | 0.26 | 1 | 0.00 | 0 | 0.00 | 0 | 0.18 | 5 |
| Centropyxis gibba Deflandre, 1929 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.90 | 7 |
| Centropyxis orbicularis Deflandre, 1929 | 0.35 | 3 | 5.16 | 3 | 0.00 | 0 | 0.05 | 3 | 3.56 | 5 | 1.29 | 9 |
| Centropyxis platystoma (Penard, 1890) Deflandre, 1929 | 0.12 | 1 | 0.00 | 0 | 0.00 | 0 | 0.23 | 8 | 0.00 | 0 | 0.15 | 4 |
| Centropyxis sylvatica (Deflandre, 1929) Bonnet and Thomas, 1955 | 5.09 | 11 | 2.58 | 4 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.09 | 1 |
| Corythion dubium Taranek, 1871 | 6.90 | 13 | 4.25 | 5 | 16.08 | 12 | 9.10 | 47 | 1.89 | 6 | 2.79 | 12 |
| Corythion orbicularis (Penard, 1910) Iudina, 1996 | 2.98 | 12 | 0.58 | 2 | 2.10 | 8 | 2.00 | 30 | 0.56 | 4 | 1.08 | 10 |
| Cryptodifflugia oviformis Penard, 1902 | 2.51 | 7 | 0.58 | 2 | 0.20 | 3 | 0.58 | 11 | 1.45 | 4 | 4.39 | 19 |
| Cyclopyxis arcelloides (Penard, 1902) Deflandre, 1929 | 1.29 | 2 | 3.83 | 4 | 0.00 | 0 | 0.00 | 0 | 11.58 | 3 | 1.47 | 10 |
| Cyclopyxis eurystoma Deflandre, 1929 | 0.58 | 4 | 0.92 | 2 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 |
| Cyclopyxis kahli (Deflandre, 1929) | 1.23 | 1 | 0.17 | 1 | 0.00 | 0 | 0.04 | 2 | 0.00 | 0 | 0.00 | 0 |
| Cylindrifflugia bacillariarum (Perty, 1849) González-Miguéns et al., 2022 | 0.00 | 0 | 0.00 | 0 | 0.26 | 1 | 0.02 | 1 | 1.00 | 3 | 0.42 | 9 |
| Cylindrifflugia elegans (Penard, 1890) González-Miguéns et al., 2022 | 0.00 | 0 | 2.08 | 4 | 0.00 | 0 | 0.00 | 0 | 1.11 | 3 | 0.18 | 2 |
| Difflugia bacillifera Penard, 1890 | 0.00 | 0 | 0.00 | 0 | 0.72 | 2 | 0.16 | 6 | 0.00 | 0 | 0.24 | 4 |
| Difflugia globulosa Dujardin, 1837 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.35 | 8 | 0.00 | 0 | 0.36 | 8 |
| Difflugia penardi Cash and Hopkinson, 1909 | 0.00 | 0 | 0.00 | 0 | 1.05 | 5 | 0.28 | 9 | 1.56 | 4 | 0.18 | 4 |
| Ellipsopyxis pauliani Bonnet, 1965 | 0.18 | 1 | 0.00 | 0 | 0.26 | 1 | 0.19 | 8 | 0.00 | 0 | 1.17 | 13 |
| Euglypha acanthophora Ehrenberg, 1841 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.02 | 1 | 0.67 | 3 | 0.00 | 0 |
| Euglypha capsiosa Coûteaux, 1978 | 0.00 | 0 | 0.17 | 1 | 0.92 | 7 | 0.09 | 5 | 0.11 | 1 | 0.00 | 0 |
| Euglypha ciliata Ehrenberg, 1848 | 0.76 | 6 | 0.00 | 0 | 0.26 | 4 | 0.19 | 8 | 0.00 | 0 | 0.81 | 14 |
| Euglypha ciliata glabra Wailes, 1915 | 0.29 | 3 | 0.33 | 1 | 0.20 | 1 | 0.37 | 10 | 0.00 | 0 | 0.18 | 3 |
| Euglypha compressa Carter, 1864 | 1.40 | 6 | 0.83 | 3 | 0.13 | 1 | 0.47 | 18 | 0.11 | 1 | 0.42 | 12 |
| Euglypha compressa glabra Wailes, 1915 | 0.06 | 1 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 |
| Euglypha cristata decora Jung, 1942 | 0.00 | 0 | 0.00 | 0 | 0.20 | 1 | 0.05 | 2 | 0.00 | 0 | 0.09 | 1 |
| Euglypha cristata Leidy, 1874 | 0.00 | 0 | 0.00 | 0 | 0.52 | 2 | 0.23 | 10 | 0.00 | 0 | 2.58 | 9 |
| Euglypha denticulata Brown, 1912 | 0.29 | 3 | 0.00 | 0 | 0.00 | 0 | 0.04 | 2 | 0.00 | 0 | 0.00 | 0 |
| Euglypha laevis (Ehrenberg, 1845) Perty, 1849 | 1.17 | 9 | 1.75 | 2 | 6.76 | 12 | 6.25 | 48 | 8.35 | 8 | 6.82 | 27 |
| Euglypha rotunda Ehrenberg, 1845 | 11.93 | 12 | 5.49 | 7 | 1.57 | 6 | 0.04 | 2 | 0.00 | 0 | 2.01 | 10 |
| Euglypha simplex Decloitre, 1965 | 0.06 | 1 | 0.33 | 1 | 0.66 | 5 | 0.04 | 2 | 0.00 | 0 | 0.12 | 4 |
| Euglypha strigosa (Ehrenberg, 1848) Leidy, 1878 | 1.05 | 9 | 0.00 | 0 | 1.77 | 11 | 1.02 | 25 | 0.00 | 0 | 0.03 | 1 |
| Euglypha strigosa glabra Wailes, 1898 | 0.76 | 7 | 0.25 | 2 | 1.90 | 12 | 1.87 | 33 | 0.45 | 2 | 0.93 | 13 |
| Euglypha tuberculata Dujardin, 1841 | 1.23 | 7 | 0.50 | 2 | 1.25 | 9 | 1.98 | 41 | 1.89 | 5 | 0.42 | 9 |
| Galeripora arenaria (Greeff, 1866) González-Miguéns et al., 2021 | 0.47 | 2 | 0.00 | 0 | 0.00 | 0 | 0.46 | 11 | 0.56 | 2 | 1.92 | 17 |
| Galeripora arenaria compressa (Chardez, 1957) González-Miguéns et al., 2021 | 0.53 | 3 | 0.00 | 0 | 0.00 | 0 | 0.16 | 8 | 0.00 | 0 | 0.21 | 3 |
| Galeripora arenaria sphagnicola (Deflandre, 1928) González-Miguéns et al., 2021 | 0.00 | 0 | 0.00 | 0 | 0.07 | 1 | 0.05 | 1 | 0.00 | 0 | 0.00 | 0 |
| Galeripora catinus (Penard, 1890) González-Miguéns et al., 2021 | 0.99 | 3 | 0.58 | 2 | 1.57 | 7 | 0.18 | 7 | 1.67 | 7 | 1.65 | 15 |
| Galeripora discoides (Ehrenberg, 1871) González-Miguéns et al., 2021 | 0.00 | 0 | 0.00 | 0 | 0.07 | 1 | 0.00 | 0 | 0.22 | 1 | 0.03 | 1 |
| Galeripora megastoma (Penard, 1902) González-Miguéns et al., 2021 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.03 | 1 |
| Gibbocarina galeata (Penard, 1890) Kosakyan et al., 2016 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.02 | 1 | 3.56 | 5 | 2.25 | 20 |
| Heleopera sphagni Leidy, 1874 | 0.00 | 0 | 0.25 | 1 | 0.07 | 1 | 0.54 | 6 | 0.00 | 0 | 0.00 | 0 |
| Heleopera sylvatica Penard, 1890 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.11 | 1 | 1.50 | 11 |
| Hyalosphenia elegans Leidy, 1874 | 0.12 | 1 | 1.08 | 3 | 0.59 | 1 | 0.12 | 5 | 0.00 | 0 | 0.00 | 0 |
| Hyalosphenia papilio Leidy, 1874 | 0.70 | 2 | 1.33 | 2 | 1.05 | 6 | 0.04 | 2 | 1.78 | 3 | 0.42 | 10 |
| Lesquereusia epistomium Penard, 1902 | 0.06 | 1 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 6.90 | 5 | 6.43 | 19 |
| Nebela collaris (Ehrenberg, 1848) sensu Kosakyan et Gomaa, 2013 | 0.00 | 0 | 0.00 | 0 | 1.31 | 8 | 0.75 | 18 | 0.00 | 0 | 0.06 | 2 |
| Nebela tincta (Leidy, 1879) Awerintzew, 1906 | 10.12 | 12 | 0.08 | 1 | 6.30 | 10 | 0.68 | 15 | 0.67 | 4 | 0.57 | 9 |
| Netzelia oviformis (Cash, 1909) Ogden, 1979 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.89 | 3 | 2.10 | 9 |
| Netzelia wailesi (Ogden, 1980) Meisterfeld, 1984 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.81 | 8 |
| Padaungiella lageniformis Penard, 1890 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.03 | 1 |
| Padaungiella wailesi Deflandre, 1936 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.02 | 1 | 0.00 | 0 | 0.00 | 0 |
| Paraquadrula irregularis Wallich, 1863 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.04 | 1 | 0.00 | 0 | 0.00 | 0 |
| Phryganella acropodia (Hertwig & Lesser, 1874) Hopkinson, 1909 | 0.00 | 0 | 0.33 | 1 | 0.00 | 0 | 0.68 | 10 | 0.45 | 1 | 0.00 | 0 |
| Phryganella hemisphaerica Penard, 1902 | 5.73 | 10 | 2.33 | 4 | 2.89 | 10 | 5.08 | 43 | 2.45 | 3 | 0.24 | 2 |
| Physochila griseola (Penard, 1911) Jung, 1955 | 0.00 | 0 | 1.42 | 1 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 1.59 | 13 |
| Placocista spinosa Penard, 1899 | 0.18 | 2 | 0.00 | 0 | 0.13 | 1 | 0.51 | 15 | 0.00 | 0 | 0.21 | 4 |
| Plagiopyxis callida Penard, 1910 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.60 | 9 |
| Plagiopyxis minuta Penard, 1910 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.03 | 1 |
| Planocarina carinata (Archer 1867) Kosakyan et al., 2016 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.05 | 3 | 0.00 | 0 | 0.00 | 0 |
| Planocarina marginata (Archer 1867) Kosakyan et al., 2016 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.02 | 1 | 0.00 | 0 | 0.00 | 0 |
| Scutiglypha scutigera (Penard, 1911) Foissner & Schiller, 2001 | 0.23 | 2 | 0.33 | 1 | 0.00 | 0 | 0.07 | 2 | 0.00 | 0 | 0.00 | 0 |
| Sphenoderia fissirostris Penard, 1890 | 0.00 | 0 | 0.00 | 0 | 0.07 | 1 | 0.05 | 2 | 0.00 | 0 | 0.51 | 5 |
| Sphenoderia lenta Schlumberger, 1845 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.02 | 1 | 0.00 | 0 | 0.00 | 0 |
| Tracheleuglypha dentata (Vejdovsky, 1882) Deflandre, 1928 | 0.23 | 1 | 0.17 | 1 | 0.46 | 2 | 0.21 | 3 | 0.00 | 0 | 0.51 | 4 |
| Trigonopyxis arcula major Chardez, 1960 | 0.35 | 2 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 |
| Trigonopyxis arcula Penard, 1912 | 2.69 | 8 | 0.50 | 2 | 2.43 | 9 | 1.02 | 10 | 0.89 | 5 | 0.36 | 7 |
| Trigonopyxis minuta Schönborn and Peschke, 1988 | 1.35 | 4 | 0.00 | 0 | 1.90 | 6 | 0.82 | 3 | 0.33 | 1 | 0.15 | 3 |
| Trinema complanatum Penard, 1890 | 0.64 | 5 | 0.67 | 4 | 1.18 | 4 | 4.99 | 40 | 4.12 | 3 | 2.10 | 18 |
| Trinema enchelys Ehrenberg, 1838 | 0.70 | 5 | 0.00 | 0 | 4.72 | 11 | 5.27 | 38 | 0.00 | 0 | 5.23 | 17 |
| Trinema grandis (Chardez, 1960) Golemansky, 1963 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.02 | 1 | 0.00 | 0 | 0.00 | 0 |
| Trinema lineare Penard, 1890 | 19.18 | 15 | 39.54 | 12 | 13.06 | 13 | 28.06 | 57 | 22.27 | 8 | 25.42 | 29 |
| Trinema lineare truncatum Chardez, 1964 | 0.00 | 0 | 0.00 | 0 | 0.00 | 0 | 0.35 | 11 | 0.22 | 1 | 0.06 | 2 |
| Trinema penardi Thomas & Chardez, 1958 | 0.12 | 1 | 0.00 | 0 | 0.00 | 0 | 0.19 | 5 | 0.00 | 0 | 0.00 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saldaev, D.; Babeshko, K.; Chernyshov, V.; Esaulov, A.; Gu, X.; Kriuchkov, N.; Mazei, N.; Saldaeva, N.; Su, J.; Tsyganov, A.; et al. Biodiversity of Terrestrial Testate Amoebae in Western Siberia Lowland Peatlands. Data 2023, 8, 173. https://doi.org/10.3390/data8110173
Saldaev D, Babeshko K, Chernyshov V, Esaulov A, Gu X, Kriuchkov N, Mazei N, Saldaeva N, Su J, Tsyganov A, et al. Biodiversity of Terrestrial Testate Amoebae in Western Siberia Lowland Peatlands. Data. 2023; 8(11):173. https://doi.org/10.3390/data8110173
Chicago/Turabian StyleSaldaev, Damir, Kirill Babeshko, Viktor Chernyshov, Anton Esaulov, Xiuyuan Gu, Nikita Kriuchkov, Natalia Mazei, Nailia Saldaeva, Jiahui Su, Andrey Tsyganov, and et al. 2023. "Biodiversity of Terrestrial Testate Amoebae in Western Siberia Lowland Peatlands" Data 8, no. 11: 173. https://doi.org/10.3390/data8110173
APA StyleSaldaev, D., Babeshko, K., Chernyshov, V., Esaulov, A., Gu, X., Kriuchkov, N., Mazei, N., Saldaeva, N., Su, J., Tsyganov, A., Yakimov, B., Yushkovets, S., & Mazei, Y. (2023). Biodiversity of Terrestrial Testate Amoebae in Western Siberia Lowland Peatlands. Data, 8(11), 173. https://doi.org/10.3390/data8110173