



Gastrointestinal Polyparasitism in Bushmeat in Zadie Department in Northeast Gabon

 ,
,  , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
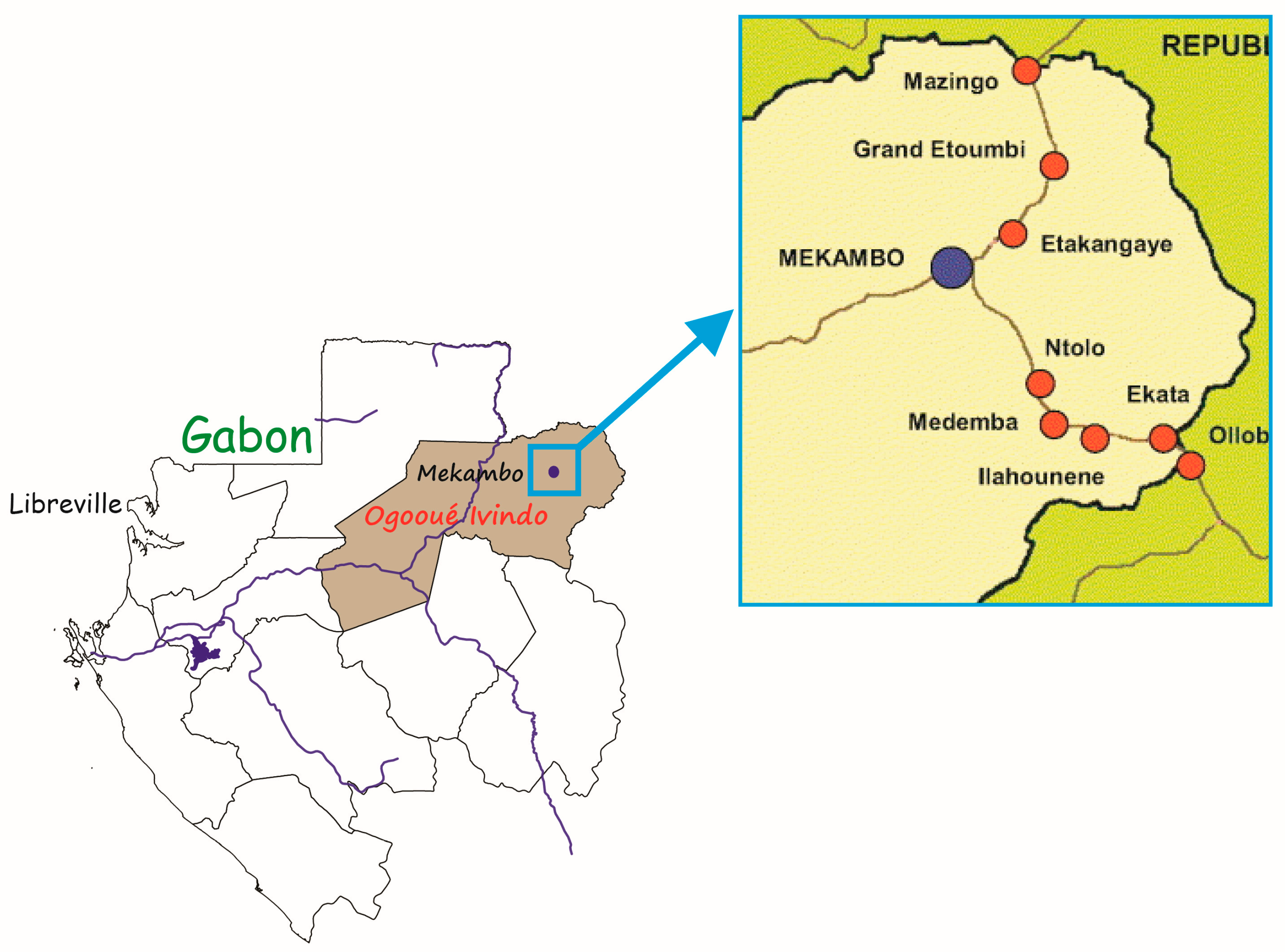
2.1. Study Areas and Sample Collection
2.2. Parasitological Analysis
2.3. Statistical Analysis
3. Results
3.1. Types of Parasites Taxa Identified
3.2. Overall Parasite Prevalence and Infestation Rate of Identified Parasites
3.3. Factors Influencing Gastrointestinal Parasitism in Bushmeat
3.3.1. The Type of Bushmeat
3.3.2. Sex
3.3.3. Age
4. Discussion
5. Conclusions and Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fa, J.E.; Wright, J.H.; Funk, S.M.; Márquez, A.L.; Olivero, J.; Farfán, M.; Guio, F.; Mayet, L.; Malekani, D.; Louzolo, C.H.; et al. Mapping the availability of bushmeat for consumption in Central African cities. Environ. Res. Lett. 2019, 14, 094002. [Google Scholar] [CrossRef]
- Fargeot, C. La Chasse Commerciale en Afrique Centrale: Une Menace Pour la Biodiversité ou une Activité Économique Durable? Le Cas de la République Centrafricaine. Ph.D. Thesis, Université Paul Valéry, Montpellier, France, 2013. Available online: https://agritrop.cirad.fr/572003/ (accessed on 14 December 2022).
- Fargeot, C.; Drouet-Hoguet, N.; Le Bel, S. The role of bushmeat in urban household consumption: Insights from Bangui, the capital city of the Central African Republic. Bois For. Trop. 2017, 332, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Nasi, R.; Taber, A.; Van Vliet, N. Empty forests, empty stomachs? Bushmeat and livelihoods in the Congo and Amazon Basins. Int. For. Rev. 2011, 13, 355–368. [Google Scholar] [CrossRef]
- Wilkie, D.S.; Carpenter, J.F. Bushmeat hunting in the Congo Basin: An assessment of impacts and options for mitigation. Biodivers. Conserv. 1999, 8, 927–955. [Google Scholar] [CrossRef]
- Binot, A.; Cornélis, D. Synthèse Bibliographique du Secteur ‘Viandes de Brousse’ au Gabon, Rapport Final; Rapport Cirad-Emvt N° 04-14; CIRAD: Montpellier, France, 2004; pp. 1–106. Available online: https://agritrop.cirad.fr/520619/1/document_520619.pdf (accessed on 5 November 2022).
- Bahuchet, S. La Filière ‘Viande De Brousse’ Les Peuples Des Forêts Tropicales Aujourd’hui: Volume II, Une Approche Thématique; Les Peuples Des Forêts Tropicales Aujourd’hui—Rapport Du Programme Apft; Commission Européenne-Apft: Bruxelles, Belgium, 2000; pp. 331–363. Available online: https://hal.archives-ouvertes.fr/hal-00547945 (accessed on 14 December 2022).
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef]
- McCarthy, J.; Moore, T.A. Emerging helminth zoonoses. Int. J. Parasitol. 2000, 30, 1351–1359. [Google Scholar] [CrossRef] [PubMed]
- Thillement, D. La Contamination Parasitaire Liée à la Consommation de Viandes, de Poissons et de Végétaux Dans les Pays Industrialisés. Doctoral Dissertation, Université de Lorraine, Nancy, France, 2015. [Google Scholar]
- Harvell, C.D.; Mitchell, C.E.; Ward, J.R.; Altizer, S.; Dobson, A.P.; Ostfeld, R.S.; Samuel, M.D. Climate Warming and Disease Risks for Terrestrial and Marine Biota. Science 2002, 296, 2158–2162. [Google Scholar] [CrossRef] [Green Version]
- Jog, M.; Watve, M. Role of parasites and commensals in shaping host behaviour. Curr. Sci. 2005, 89, 1181–1194. [Google Scholar]
- Kashid, K.; Shrikhande, G.; Bhojne, G. Incidence of gastro-intestinal helminths in captive wild animals at different locations. Zoos’ Print J. 2003, 18, 1053–1054. [Google Scholar] [CrossRef]
- Opara, M.; Osuji, C.T.; Opara, J.A. Gastrointestinal parasitism in captive animals at the zoological garden, nekede owerri, southeast nigeria. Rep. Opin. 2010, 2, 21–28. [Google Scholar]
- Dryden, P.M.W.; Payne, P.P.A.; Ridley, P.R.; Smith, R.V. Comparison of Common Fecal Flotation Techniques for the Recovery of Parasite Eggs and Oocysts. Vet. Ther. 2005, 6, 15–28. [Google Scholar]
- Gillespie, T.R. Noninvasive Assessment of Gastrointestinal Parasite Infections in Free-Ranging Primates. Int. J. Primatol. 2006, 27, 1129–1143. [Google Scholar] [CrossRef] [Green Version]
- Soulsby, E.J.L. Helminths, Arthropods and Protozoa of Domesticated Animals. 1968. Available online: https://www.cabdirect.org/cabdirect/abstract/19682902735 (accessed on 6 November 2022).
- Posit|The Open-Source Data Science Company. Available online: https://posit.co/ (accessed on 24 December 2022).
- Okoye, I.C.; Ozioko, K.U.; Obiezue, N.R.; Ikele, B.C. Intestinal parasitic fauna and zoonotic potentials of commonly consumed wildlife. Helminthologia 2015, 52, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Belem, A.M.G.; Bakoné, É.U. Parasites gastro-intestinaux d’antilopes et de buffles (Syncerus caffer brachyceros) du ranch de gibier de Nazinga au Burkina Faso. Biotechnol. Agron. Soc. Environ. 2009, 13, 493–498. [Google Scholar]
- Graber, M.; Doutre, M.-P.; Finelle, P.; Kéravec, J.; Ducroz, G.; Mokotaingar, P. Les helminthes de quelques artiodactyles sauvages appartenant aux familles des bovidés et des suidés: Ces mammifères, en République du Tchad et en R.C.A., sont-ils des réservoirs de parasites pour les animaux domestiques vivant à leur contact? Rev. Elev. Med. Vet. Pays Trop. 1964, 17, 377–420. Available online: https://agritrop.cirad.fr/433708/ (accessed on 14 December 2022). [CrossRef] [Green Version]
- Pourrut, X.; Diffo, J.; Somo, R.; Bilong, C.B.; Delaporte, E.; LeBreton, M.; Gonzalez, J. Prevalence of gastrointestinal parasites in primate bushmeat and pets in Cameroon. Vet. Parasitol. 2011, 175, 187–191. [Google Scholar] [CrossRef]
- Parker, J.M.; Goldenberg, S.Z.; Letitiya, D.; Wittemyer, G. Strongylid infection varies with age, sex, movement and social factors in wild African elephants. Parasitology 2020, 147, 348–359. [Google Scholar] [CrossRef] [Green Version]
- Aissa, S.; Rachida, M.; Fatima, H.; Widade, O. Gastrointestinal Nematode Infections in Antelopes from Morocco: A Coprological Survey. Acta Vet. 2021, 71, 47–60. [Google Scholar] [CrossRef]
- Omonona, A.O.; Ademola, I.O.; Ayansola, V.I. Prevalence of gastrointestinal parasites of Walter’s duiker (Philantomba walteri) in Ondo State, Nigeria. Afr. J. Biomed. Res. 2019, 22, 73–78. [Google Scholar]
- Abara, P.N.; Adjeroh, L.; Nwachukwu, M.O.; Osinomumu, I. Preliminary Survey of the Intestinal Helminths of Grasscutter and Antelope (Bush Meat) in Omagwa Rivers State. Caribb. J. Sci. Technol. 2021, 9, 1–6. [Google Scholar] [CrossRef]
- Beiromvand, M.; Akhlaghi, L.; Massom, S.H.F.; Meamar, A.R.; Darvish, J.; Razmjou, E. Molecular Identification of Echinococcus multilocularis Infection in Small Mammals from Northeast, Iran. PLoS Negl. Trop. Dis. 2013, 7, e2313. [Google Scholar] [CrossRef] [Green Version]
- Deplazes, P.; van Knapen, F.; Schweiger, A.; Overgaauw, P.A. Role of pet dogs and cats in the transmission of helminthic zoonoses in Europe, with a focus on echinococcosis and toxocarosis. Vet. Parasitol. 2011, 182, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Sowemimo, O.; Asaolu, S. Epidemiology of intestinal helminth parasites of dogs in Ibadan, Nigeria. J. Helminthol. 2008, 82, 89–93. [Google Scholar] [CrossRef]
- Onah, I.E.; Umeike, E.C. Intestinal parasitic fauna and prevalence in african giant rat (Cricetomys gambianus) in nsukka metropolis south eastern nigeria. Fudma J. Sci. 2022, 6, 101–106. [Google Scholar] [CrossRef]
- Apio, A.; Plath, M.; Wronski, T. Patterns of gastrointestinal parasitic infections in the bushbuck Tragelaphus scriptus from the Queen Elizabeth National Park, Uganda. J. Helminthol. 2006, 80, 213–218. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Village | N° | Sex | Age | Type of Bushmeat | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Males | Females | Young | Adult | Antelope | Duiker | Porcupine | Small Monkey | Pangolin | Nandinia | Genet | Crocodile | ||
| Ekata | 10 | 5 | 5 | 0 | 10 | 1 | 8 | 1 | 0 | 0 | 0 | 0 | 0 |
| Ilahounéné | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Mékouma | 12 | 7 | 5 | 3 | 9 | 3 | 8 | 1 | 0 | 0 | 0 | 0 | 0 |
| Malassa | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Ntolo | 5 | 3 | 2 | 0 | 5 | 2 | 0 | 2 | 1 | 0 | 0 | 0 | 0 |
| Ego-Poma | 4 | 3 | 1 | 0 | 4 | 0 | 3 | 1 | 0 | 0 | 0 | 0 | 0 |
| Grand-Etoumbi | 34 | 15 | 19 | 7 | 27 | 6 | 17 | 3 | 5 | 1 | 1 | 0 | 1 |
| Zoula | 9 | 5 | 4 | 3 | 6 | 0 | 5 | 4 | 0 | 0 | 0 | 0 | 0 |
| Etchéla-Edounga | 21 | 7 | 14 | 3 | 18 | 6 | 11 | 3 | 0 | 0 | 1 | 0 | 0 |
| Komambela | 13 | 8 | 5 | 1 | 12 | 2 | 5 | 3 | 2 | 0 | 0 | 1 | 0 |
| Malouma | 3 | 1 | 2 | 0 | 3 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 113 | 54 | 59 | 17 | 96 | 24 | 58 | 18 | 8 | 1 | 2 | 1 | 1 |
| Parasite Diversity | Adult or Larvae | Eggs or Cyst | Infestation Rate (%) |
|---|---|---|---|
| Strongylids eggs | √ | √ | 54% |
| Ascaris spp. | √ | √ | 18.6 |
| Balantidium spp. | - | √ | 10.6 |
| Capillaria spp. | - | √ | 7.96 |
| Eimeria spp. | - | √ | 15.04 |
| Entamoeba spp. | - | √ | 7.96 |
| Enterobius spp. | - | √ | 7.08 |
| Fasciola spp. | - | √ | 15.9 |
| Mammomonogamus spp. | - | √ | 4.42 |
| Paramphistomum spp. | - | √ | 18.6 |
| Protostrongylus spp. | √ | - | 4.42 |
| Strongyloïdes spp. | - | √ | 15.6 |
| Taenia spp. | √ | - | 0.9 |
| Toxocara spp. | √ | - | 6.2 |
| Trichuris spp. | √ | √ | 34.51 |
| Parasites Taxa | Animal Species | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Antelope | Nandinia | Crocodile | Duiker | Genet | Pangolin | Cercopithecus | Porcupine | ||
| (n = 24) | (n = 2) | (n = 1) | (n = 58) | (n = 1) | (n = 1) | (n = 8) | (n = 18) | ||
| Strongylid species | Eggs | 75 | 0 | 0 | 41.4 | 100 | 100 | 50 | 72.2 |
| Adult | 4.2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Total | 75 | 0 | 0 | 41.4 | 100 | 100 | 50 | 72.2 | |
| Strongyloïdes spp. | Eggs | 20.8 | 0 | 0 | 7 | 100 | 0 | 25 | 50 |
| Adult | 0 | 0 | 0 | 0 | 100 | 0 | 0 | 5.5 | |
| Total | 20.8 | 0 | 0 | 7 | 100 | 0 | 25 | 50 | |
| Eimeria spp. | Oocysts | 20.8 | 0 | 0 | 17.5 | 0 | 0 | 0 | 11.1 |
| Balantidium spp. | Oocysts | 8.3 | 0 | 0 | 8.8 | 100 | 0 | 0 | 22.2 |
| Entamoeba spp. | Oocysts | 16.7 | 0 | 0 | 3.5 | 0 | 0 | 25 | 5.6 |
| Ascaris spp. | Eggs | 20.8 | 50 | 0 | 19.0 | 0 | 0 | 0 | 11.1 |
| Adult | 0 | 0 | 0 | 5.2 | 0 | 0 | 0 | 0 | |
| Total | 20.8 | 50 | 0 | 20.7 | 0 | 0 | 0 | 11.1 | |
| Trichuris spp. | Eggs | 42 | 50 | 0 | 26 | 100 | 0 | 87.5 | 28 |
| Adult | 17 | 50 | 0 | 7 | 0 | 0 | 37.5 | 33.3 | |
| Total | 42 | 50 | 0 | 26 | 100 | 0 | 87.5 | 39 | |
| Capillaria spp. | Eggs | 0 | 0 | 0 | 3.5 | 0 | 100 | 25 | 22.2 |
| Protostrongylus spp. | Larvae | 8.3 | 0 | 0 | 0 | 0 | 0 | 12.5 | 11.1 |
| Enterobius spp. | Eggs | 0 | 0 | 0 | 5.3 | 0 | 0 | 25 | 16.7 |
| Fasciola spp. | Eggs | 37.5 | 0 | 0 | 15.8 | 0 | 0 | 0 | 0 |
| Paramphistomum spp. | Eggs | 25 | 0 | 0 | 26.3 | 0 | 0 | 0 | 0 |
| Mammomonogamus spp. | Eggs | 12.5 | 0 | 0 | 1.8 | 0 | 0 | 0 | 5.6 |
| Taenia spp. | Adults | 0 | 0 | 0 | 0 | 100 | 0 | 0 | 0 |
| Toxocara spp. | Adults | 4.2 | 0 | 0 | 5.2 | 0 | 0 | 37.5 | 0 |
| Distribution by Gender | Males | Females | χ2 | df | Comparison Test | S | M+ | F+ | ||
|---|---|---|---|---|---|---|---|---|---|---|
| N/53 | P (%) | N/59 | P (%) | p-value | ||||||
| Strongylids species | 29 | 54.7 | 31 | 52.5 | 0.92 | 2 | 0.63 | − | + | |
| Strongyloïdes spp. | 7 | 13.2 | 14 | 23.7 | 2.02 | 1 | 0.15 | − | + | |
| Eimeria spp. | 10 | 18.9 | 7 | 11.9 | 1.06 | 1 | 0.30 | − | + | |
| Balantidium spp. | 5 | 9.4 | 7 | 11.9 | 0.12 | 1 | 0.72 | − | + | |
| Entamoeba spp. | 3 | 5.7 | 6 | 10.2 | - | 1 | 0.49 | − | + | |
| Ascaris spp. | 8 | 15.1 | 13 | 22.0 | 0.88 | 1 | 0.34 | − | + | |
| Trichuris spp. | 18 | 33.9 | 20 | 33.9 | 5.1 | 1 | 0.99 | − | ||
| Capillaria spp. | 2 | 3.8 | 7 | 11.9 | - | 1 | 0.17 | − | + | |
| Protostrongylus spp. | 0 | 0 | 4 | 6.8 | - | 1 | 0.12 | − | + | |
| Enterobius spp. | 2 | 3.8 | 5 | 8.5 | - | 1 | 0.43 | − | + | |
| Fasciola spp. | 9 | 17.0 | 11 | 18.6 | - | 1 | 0.81 | − | + | |
| Paramphistomum spp. | 10 | 19.0 | 3 | 5.1 | - | 1 | 0.03 | + | + | |
| Mammomonogamus spp. | 2 | 3.8 | 3 | 5.1 | - | 1 | 1 | − | + | |
| Toxocara spp. | 4 | 7.5 | 0 | 0 | - | 1 | + | |||
| Taenia spp. | 1 | 1.9 | 0 | 0 | - | 1 | + | |||
| 5 | 9 | |||||||||
| Mean | 7.9 | 14.9 | 10.2 | 17.3 | ||||||
| Median | 7.0 | 13.2 | 7.0 | 11.9 | ||||||
| Distribution by Age | Adult | Young | χ2 | df | Comparison Test | S | A+ | Y+ | ||
|---|---|---|---|---|---|---|---|---|---|---|
| N/96 | P (%) | N/17 | P (%) | p-value | ||||||
| Strongylids species | 52 | 54.2 | 9 | 53 | 1 | 1 | − | + | ||
| Strongyloides spp. | 19 | 19.8 | 2 | 11.8 | 1 | 0.66 | − | + | ||
| Eimeria spp. | 14 | 14.6 | 3 | 18.7 | 1 | 1 | − | + | ||
| Balantidium spp. | 10 | 10.4 | 2 | 11.8 | 1 | 1 | − | + | ||
| Entamoeba spp. | 9 | 9.4 | 0 | 0 | 1 | 0.41 | − | + | ||
| Ascaris spp. | 17 | 17.7 | 4 | 23.5 | 1 | 0.82 | − | + | ||
| Trichuris ssp. | 35 | 36.5 | 4 | 23.5 | 1 | 0.45 | − | + | ||
| Capillaria spp. | 6 | 6.25 | 3 | 17.65 | 1 | 0.27 | − | + | ||
| Protostrongylus spp. | 2 | 2.1 | 3 | 17.65 | 1 | 0.025 | + | + | ||
| Enterobius spp. | 7 | 7.3 | 1 | 5.88 | 1 | 1 | − | + | ||
| Fasciola spp. | 15 | 15.6 | 3 | 17.6 | 1 | 1 | − | + | ||
| Paramphistomum spp. | 17 | 17.7 | 4 | 25 | 1 | 0.73 | − | + | ||
| Mammomonogamus spp. | 5 | 5.2 | 0 | 0 | 1 | 1 | − | + | ||
| Toxocara spp. | 5 | 5.3 | 2 | 13.33 | 1 | 0.54 | − | + | ||
| Taenia spp. | 1 | 1.04 | 0 | 0 | 1 | 1 | − | + | ||
| 6 | 9 | |||||||||
| Mean | 15.5 | 16.1 | 2.7 | 16.8 | ||||||
| Median | 14 | 14.6 | 3 | 18.5 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maganga, G.D.; Makouloutou-Nzassi, P.; Boundenga, L.; Maganga Landjekpo, H.N.; Bangueboussa, F.; Ndong Mebaley, T.; Mounioko, F.; Gbati, O.B. Gastrointestinal Polyparasitism in Bushmeat in Zadie Department in Northeast Gabon. Vet. Sci. 2023, 10, 229. https://doi.org/10.3390/vetsci10030229
Maganga GD, Makouloutou-Nzassi P, Boundenga L, Maganga Landjekpo HN, Bangueboussa F, Ndong Mebaley T, Mounioko F, Gbati OB. Gastrointestinal Polyparasitism in Bushmeat in Zadie Department in Northeast Gabon. Veterinary Sciences. 2023; 10(3):229. https://doi.org/10.3390/vetsci10030229
Chicago/Turabian StyleMaganga, Gael Darren, Patrice Makouloutou-Nzassi, Larson Boundenga, Hurlis Nesla Maganga Landjekpo, Felicien Bangueboussa, Telstar Ndong Mebaley, Franck Mounioko, and Oubri Bassa Gbati. 2023. "Gastrointestinal Polyparasitism in Bushmeat in Zadie Department in Northeast Gabon" Veterinary Sciences 10, no. 3: 229. https://doi.org/10.3390/vetsci10030229
APA StyleMaganga, G. D., Makouloutou-Nzassi, P., Boundenga, L., Maganga Landjekpo, H. N., Bangueboussa, F., Ndong Mebaley, T., Mounioko, F., & Gbati, O. B. (2023). Gastrointestinal Polyparasitism in Bushmeat in Zadie Department in Northeast Gabon. Veterinary Sciences, 10(3), 229. https://doi.org/10.3390/vetsci10030229