Prevalence of Winter Ticks (Dermacentor albipictus) in Hunter-Harvested Wild Elk (Cervus canadensis) from Pennsylvania, USA (2017–2018)

 ,
, 
 and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Tick Collection and Storage
2.2. Tick Identification
2.3. Statistics
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mooring, M.S.; Samuel, W.M. Premature loss of winter hair in free-ranging moose (Alces alces) infested with winter ticks (Dermacentor albipictus) is correlated with grooming rate. Can. J. Zool. 1999, 77, 148–156. [Google Scholar] [CrossRef]
- Musante, A.R.; Pekins, P.J.; Scarpitti, D.L. Metabolic impacts of winter tick infestations on calf. Alces 2007, 43, 101–110. [Google Scholar]
- DeIgiudice, G.D.; Peterson, R.O.; Samuel, W.M. Trends of Winter Nutritional Restriction, Ticks, and Numbers of Moose on Isle Royale. J. Wildl. Manag. 1997, 61, 895. [Google Scholar] [CrossRef]
- Ewing, S.A.; Panciera, R.J.; Kocan, K.M.; Ge, N.L.; Welsh, R.D.; Olson, R.W.; Barker, R.W.; Rice, L.E. A Winter Outbreak of Anaplasmosis in a Nonendemic Area of Oklahoma: A Possible Role for Dermacentor albipictus. J. Vet. Diagn. Investig. 1997, 9, 206–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swei, A.; O’Connor, K.E.; Couper, L.I.; Thekkiniath, J.; Conrad, P.A.; Padgett, K.A.; Burns, J.; Yoshimizu, M.H.; Gonzales, B.; Munk, B.; et al. Evidence for transmission of the zoonotic apicomplexan parasite Babesia duncani by the tick Dermacentor albipictus. Int. J. Parasitol. 2019, 49, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Ameri, M.; Anderson, W.I.; Holman, P.J.; Palmer, G.W. Babesia odocoilei infection in a North American elk (Cervus elaphus canadensis). Comp. Clin. Pathol. 2012, 21, 363–365. [Google Scholar] [CrossRef]
- Zaugg, J.L.; Goff, W.L.; Foreyt, W.; Hunter, D.L. Susceptibility of elk (Cervus elaphus) to experimental infection with Anaplasma marginale and A. ovis. J. Wildl. Dis. 1996, 32, 62–66. [Google Scholar] [CrossRef] [Green Version]
- Billeter, S.A.; Osikowicz, L.M.; Burns, J.E.; Konde, L.; Gonzales, B.J.; Hu, R.; Kosoy, M.Y. Molecular surveillance for Bartonella, Borrelia, and Rickettsia species in ticks from desert bighorn sheep (Ovis canadensis) and mule deer (Odocoileus hemionus) in Southern California, USA. J. Wildl. Dis. 2018, 54, 161. [Google Scholar] [CrossRef]
- Crosbie, P.R.; Goff, W.L.; Stiller, D.; Jessup, D.A.; Boyce, W.M. The Distribution of Dermacentor hunteri and Anaplasma sp. in Desert Bighorn Sheep (Ovis canadensis). J. Parasitol. 1997, 83, 31. [Google Scholar] [CrossRef]
- Hairgrove, T.B.; Craig, T.M.; Budke, C.M.; Rodgers, S.J.; Gill, R.J. Seroprevalence of Anaplasma marginale in Texas Cattle. Prev. Vet. Med. 2014, 116, 188–192. [Google Scholar] [CrossRef]
- Kocan, K.; Coetzee, J.; Step, D.; Fuente, J.; Blouin, E.; Reppert, E.; Simpson, K.; Boileau, M. Current challenges in the diagnosis and control of bovine anaplasn1osis. Bov. Pract. 2012, 46, 67–77. [Google Scholar]
- Addison, E.M.; McLaughlin, R.F. Growth and Development of Winter Tick, Dermacentor albipictus, on Moose, Alces alces. J. Parasitol. 1988, 74, 670–678. [Google Scholar] [CrossRef]
- Bondo, K.J.; Macbeth, B.; Schwantje, H.; Orsel, K.; Culling, D.; Culling, B.; Tryland, M.; Nymo, I.H.; Kutz, S. Health survey of boreal caribou (Rangifer tarandus caribou) in Northeastern British Columbia, Canada. J. Wildl. Dis. 2019, 55, 544. [Google Scholar] [CrossRef] [PubMed]
- Welch, D.; Samuel, W.; Wilke, C. Suitability of moose, elk, mule deer, and white-tailed deer as hosts for winter ticks (Dermacentor albipictus). Can. J. Zool. 1991, 69, 2300–2305. [Google Scholar] [CrossRef]
- Mooring, M.; Samuel, W. The biological basis of grooming in moose (Alces alces): Programmed versus stimulus-driven grooming. Anim. Behav. 1998, 56, 1561–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mooring, M.; Samuel, W.M. Tick-removal grooming by elk (Cervus elaphus): Testing the principles of the programmed-grooming hypothesis. Can. J. Zool. 1998, 76, 740–750. [Google Scholar] [CrossRef]
- Samuel, W.M.; Welch, D.; Smith, B.L. Ectoparasites from elk (Cervus elaphus nelsoni) from Wyoming. J. Wildl. Dis. 1991, 27, 446–451. [Google Scholar] [CrossRef] [Green Version]
- Calvente, E.; Chinnici, N.; Brown, J.; Banfield, J.E.; Brooks, J.W.; Yabsley, M.J. Winter Tick (Dermacentor albipictus)-Associated Dermatitis in a Wild Elk (Cervus canadensis) in Pennsylvania, USA. J. Wildl. Dis. 2020, 56, 247–250. [Google Scholar] [CrossRef]
- Samuel, W.M. Factors affecting epizootics of winter ticks and mortality of moose. Alces 2007, 43, 39–48. [Google Scholar]
- Musante, A. Characteristics and Dynamics of a Moose Population in Northern New Hampshire. Master’s Thesis, University of New Hampshire, Durham, NH, USA, December 2006. [Google Scholar]
- Ball, K. Moose Density, Habitat, and Winter Tick Epizootics in a Changing Climate. Master’s Thesis, University of New Hampshire, Durham, NH, USA, May 2017. [Google Scholar]
- Musante, A.R.; Pekins, P.J.; Scarpitti, D.L. Characteristics and dynamics of a regional moose Alces alces population in the northeastern United States. Wildl. Biol. 2010, 16, 185–204. [Google Scholar] [CrossRef] [Green Version]
- Keirans, J.E.; Litwak, T.R. Pictorial key to the adults of hard ticks, family Ixodidae (Ixodida: Ixodoidea), east of the Mississippi River. J. Med. Entomol. 1989, 26, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Krakowetz, C.N.; Lindsay, L.R.; Chilton, N.B. Genetic variation in the mitochondrial 16S ribosomal RNA gene of Ixodes scapularis (Acari: Ixodidae). Parasites Vectors 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Nadolny, R.M.; Wright, C.L.; Hynes, W.L.; Sonenshine, D.E.; Gaff, H.D. Ixodes affinis (Acari: Ixodidae) in southeastern Virginia and implications for the spread of Borrelia burgdorferi, the agent of Lyme disease. J. Vector Ecol. 2011, 36, 464–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, J.F.; Emery, D.; Bogema, D.R.; Jenkins, C. Detection of Theileria orientalis genotypes in Haemaphysalis longicornis ticks from southern Australia. Parasites Vectors 2015, 8, 229. [Google Scholar] [CrossRef] [Green Version]
- Baer-Lehman, M.; Light, T.; Fuller, N.; Barry-Landis, K.; Kindlin, C.; Stewart, R. Evidence for competition between Ixodes scapularis and Dermacentor albipictus feeding concurrently on white-tailed deer. Exp. Appl. Acarol. 2012, 58, 301–314. [Google Scholar] [CrossRef]
- Slabach, B.L.; McKinney, A.; Cunningham, J.; Hast, J.T.; Cox, J.J. A Survey of Tick Species in a Recently Reintroduced Elk (Cervus canadensis) Population in Southeastern Kentucky, USA, with Potential Implications for Interstate Translocation of Zoonotic Disease Vectors. J. Wildl. Dis. 2018, 54, 366–370. [Google Scholar] [CrossRef]
- Schulze, T.L.; Bowen, G.S.; Lakat, M.F.; Parkin, W.E.; Shisler, J.K. Seasonal abundance and hosts of Ixodes dammini (Acari: Ixodidae) and other ixodid ticks from an endemic Lyme disease focus in New Jersey, USA. J. Med. Entomol. 1986, 23, 105–109. [Google Scholar] [CrossRef]
- Dryden, M.W.; Payne, P.A. Biology and control of ticks infesting dogs and cats in North America. Vet. Ther. Res. Appl. Vet. Med. 2004, 5, 139–154. [Google Scholar]
- Osburn, R.L. Timing of Ecdysis and Spermatogenesis in the Winter Tick, Dermacentor albipictus (Packard). Ann. Entomol. Soc. Am. 1981, 74, 177–179. [Google Scholar] [CrossRef]
- Drummond, R.O. Seasonal Activity of Ticks (Acarina: Metastigmata) on Cattle in Southwestern Texas. Ann. Entomol. Soc. Am. 1967, 60, 439–447. [Google Scholar] [CrossRef] [Green Version]
- Thorne, E.T.; Dean, R.E.; Hepworth, W.G. Nutrition during Gestation in Relation to Successful Reproduction in Elk. J. Wildl. Manag. 1976, 40, 330–335. [Google Scholar] [CrossRef]
- Kocan, A.A.; Mukolwe, S.W.; Murphy, G.L.; Barker, R.W.; Kocan, K.M. Isolation of Borrelia burgdorferi (Spirochaetales: Spirochaetaceae) from Ixodes scapularis and Dermacentor albipictus ticks (Acari: Ixodidae) in Oklahoma. J. Med. Entomol. 1992, 29, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Baldridge, G.D.; Scoles, G.A.; Burkhardt, N.Y.; Schloeder, B.; Kurtti, T.J.; Munderloh, U.G. Transovarial Transmission of Francisella-Like Endosymbionts and Anaplasma phagocytophilum Variants in Dermacentor albipictus (Acari: Ixodidae). J. Med. Entomol. 2009, 46, 625–632. [Google Scholar] [PubMed] [Green Version]
- Randolph, S.; Gern, L.; Nuttal, P. Co-feeding ticks: Epidemiological significance for tick-borne pathogen transmission. Parasitol. Today 1996, 12, 472–490. [Google Scholar] [CrossRef] [PubMed]
- Kocan, K.M.; Goff, W.L.; Stiller, D.; Claypool, P.L.; Edwards, W.; Ewing, S.A.; Hair, J.A.; Barron, S.J. Persistence of Anaplasma marginale (Rickettsiales: Anaplasmataceae) in male Dermacentor andersoni (Acari: Ixodidae) transferred successively from infected to susceptible calves. J. Med. Entomol. 1992, 29, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.L.; Serfass, T.L.; Cogan, R.; Rhodes, O.E. Microsatellite variation in the reintroduced Pennsylvania elk herd. Mol. Ecol. 2002, 11, 1299–1310. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
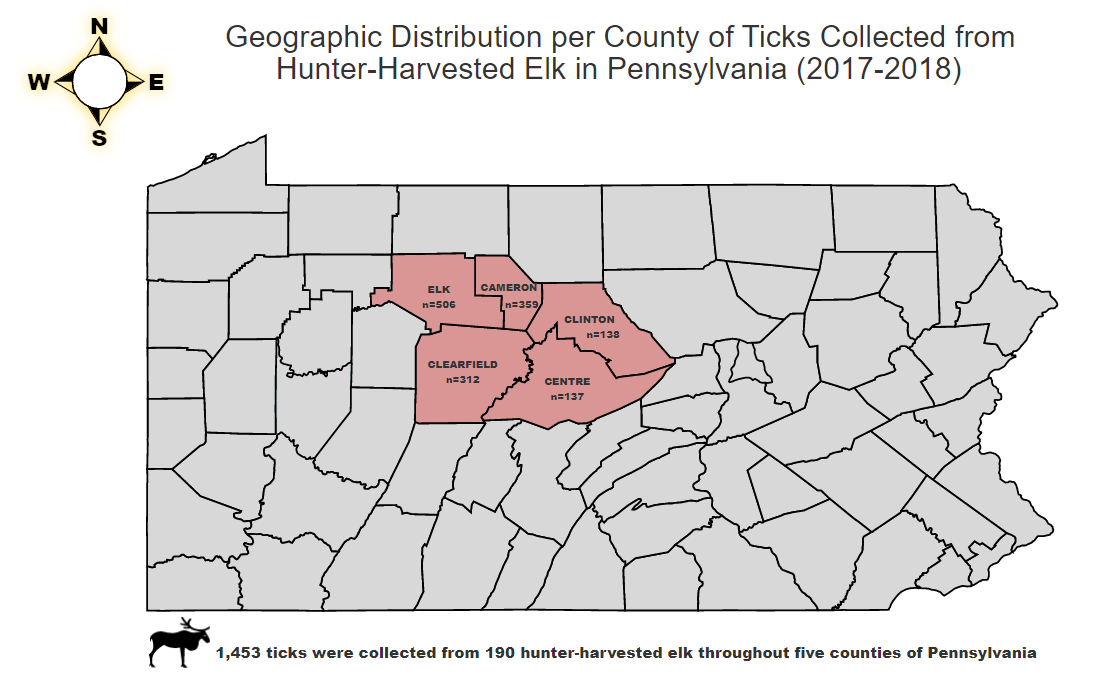
| Tick Counts per County | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| County | 2017 | 2018 | Total (2017/2018) | ||||||
| Deer Tick | Winter Tick | Total Ticks | Deer Tick | Winter Tick | Total Ticks | Deer Tick | Winter Tick | Total Ticks | |
| Elk | 288 | 9 | 297 | 192 * | 17 | 201 * | 480 | 26 | 506 |
| Clearfield | 213 | 23 | 236 * | 72 * | 4 | 76 * | 285 | 27 | 312 |
| Cameron | 218 | 15 | 223 * | 125 | 1 | 126 | 343 * | 16 | 359 * |
| Centre | 62 | 11 | 73 * | 60 | 4 | 64 | 122 | 15 | 137 * |
| Clinton | 86 | 5 | 91 * | 39 | 8 | 47 | 125 | 13 | 138 |
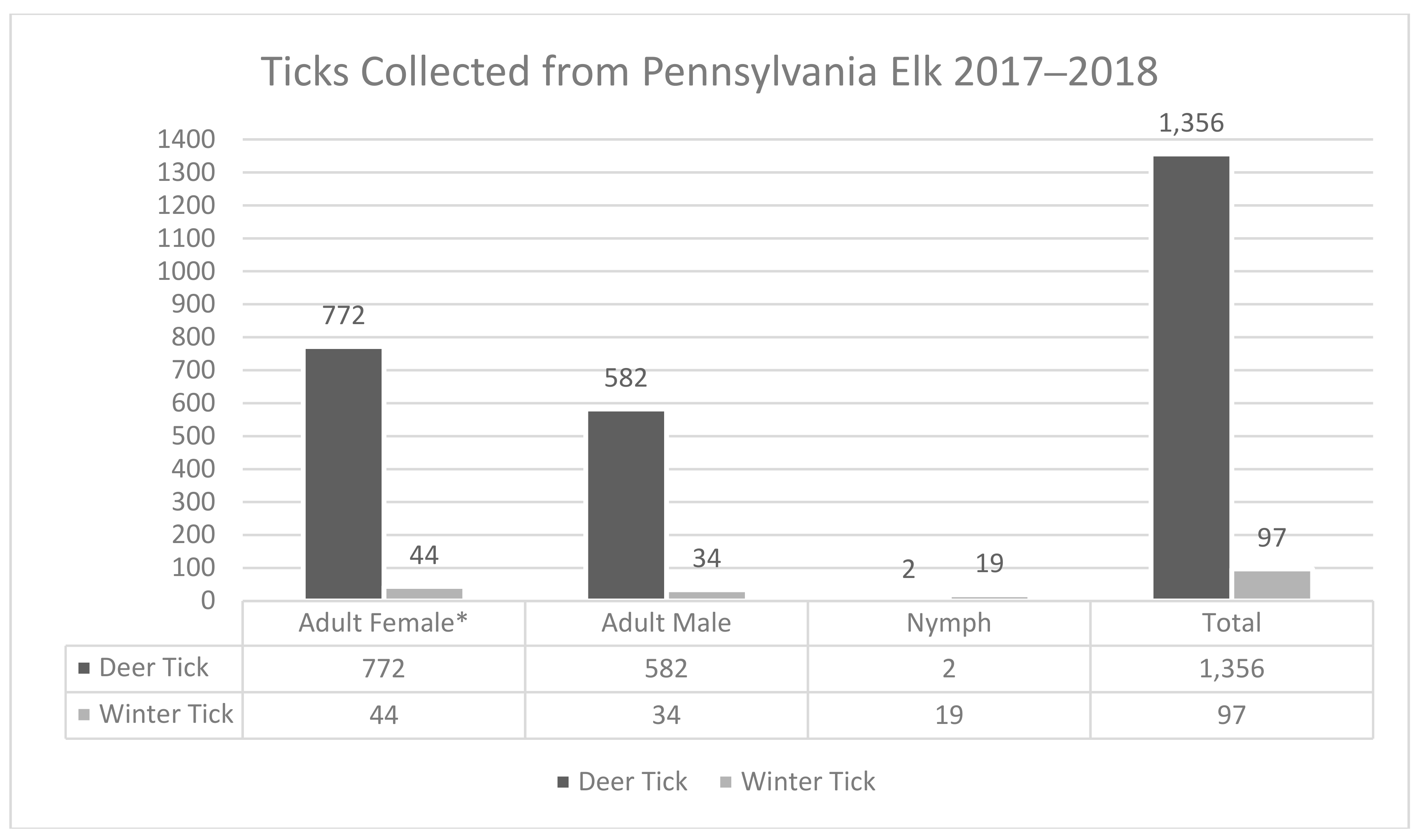
| Deer Tick | Winter Tick | Total Ticks | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 2017 | 2018 | 2017–2018 | 2017 | 2018 | Total | 2017 | 2018 | 2017–2018 | |
| Tick Collections | 868 | 488 | 1356 | 63 | 34 | 97 | 931 | 522 | 1453 |
| Average Tick Load | 8.8 | 5.4 | 7.1 | 0.6 | 0.4 | 0.5 | 9.4 | 5.7 | 7.7 |
| Range of Ticks Per Elk | 0–42 | 0–18 | 0–42 | 0–9 | 0–8 | 0–9 | 0–42 | 0–18 | 0–42 |
| Total Adult Male | 374 | 208 | 582 | 22 | 12 | 34 | 396 | 220 | 616 |
| Total Adult Female | 492 | 280 | 772 | 24 | 20 | 44 | 516 | 300 | 816 |
| Total Nymphs | 2 | - | 2 | 17 | 2 | 19 | 19 | 2 | 21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calvente, E.; Pelletier, S.; Banfield, J.; Brown, J.; Chinnici, N. Prevalence of Winter Ticks (Dermacentor albipictus) in Hunter-Harvested Wild Elk (Cervus canadensis) from Pennsylvania, USA (2017–2018). Vet. Sci. 2020, 7, 177. https://doi.org/10.3390/vetsci7040177
Calvente E, Pelletier S, Banfield J, Brown J, Chinnici N. Prevalence of Winter Ticks (Dermacentor albipictus) in Hunter-Harvested Wild Elk (Cervus canadensis) from Pennsylvania, USA (2017–2018). Veterinary Sciences. 2020; 7(4):177. https://doi.org/10.3390/vetsci7040177
Chicago/Turabian StyleCalvente, Elizabeth, Samantha Pelletier, Jeremiah Banfield, Justin Brown, and Nicole Chinnici. 2020. "Prevalence of Winter Ticks (Dermacentor albipictus) in Hunter-Harvested Wild Elk (Cervus canadensis) from Pennsylvania, USA (2017–2018)" Veterinary Sciences 7, no. 4: 177. https://doi.org/10.3390/vetsci7040177
APA StyleCalvente, E., Pelletier, S., Banfield, J., Brown, J., & Chinnici, N. (2020). Prevalence of Winter Ticks (Dermacentor albipictus) in Hunter-Harvested Wild Elk (Cervus canadensis) from Pennsylvania, USA (2017–2018). Veterinary Sciences, 7(4), 177. https://doi.org/10.3390/vetsci7040177