
Characterization of Cecal Smooth Muscle Contraction in Laying Hens



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Feeding
2.2. Buffers and Solutions
2.3. Tissue Preparation
2.4. Isometric Force Measurement of the Muscle Contraction
2.5. Data Analyses
3. Results
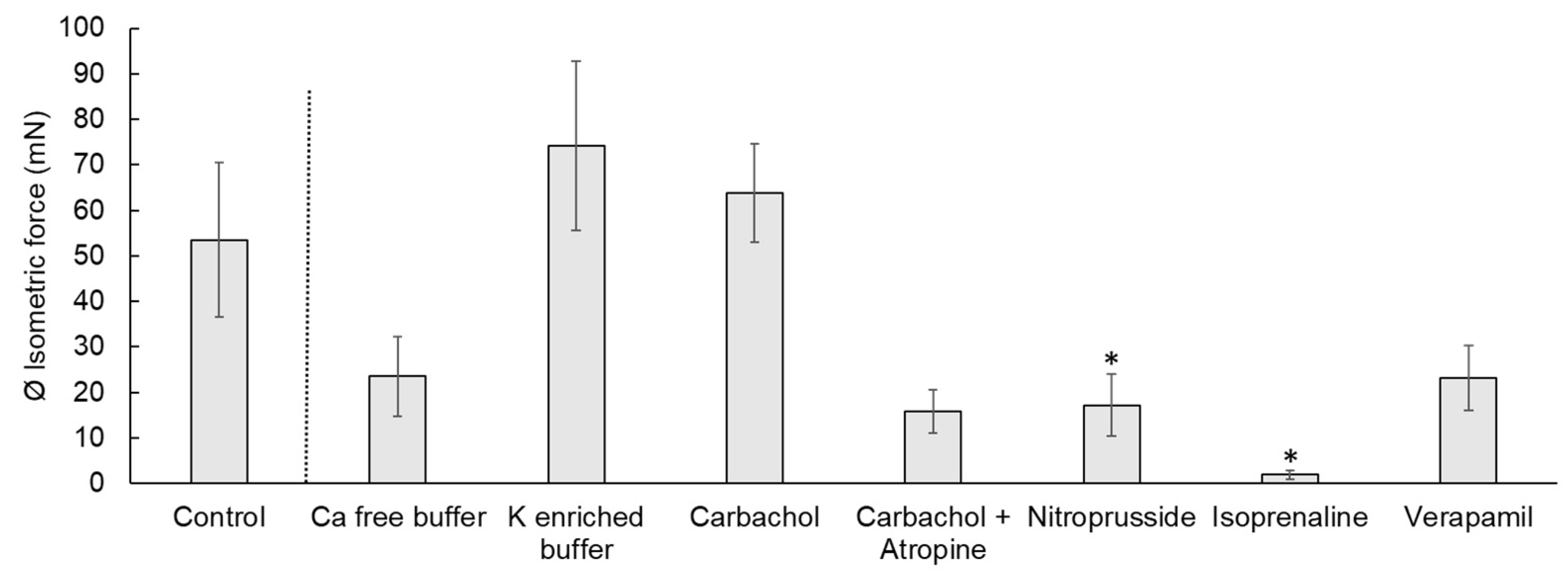
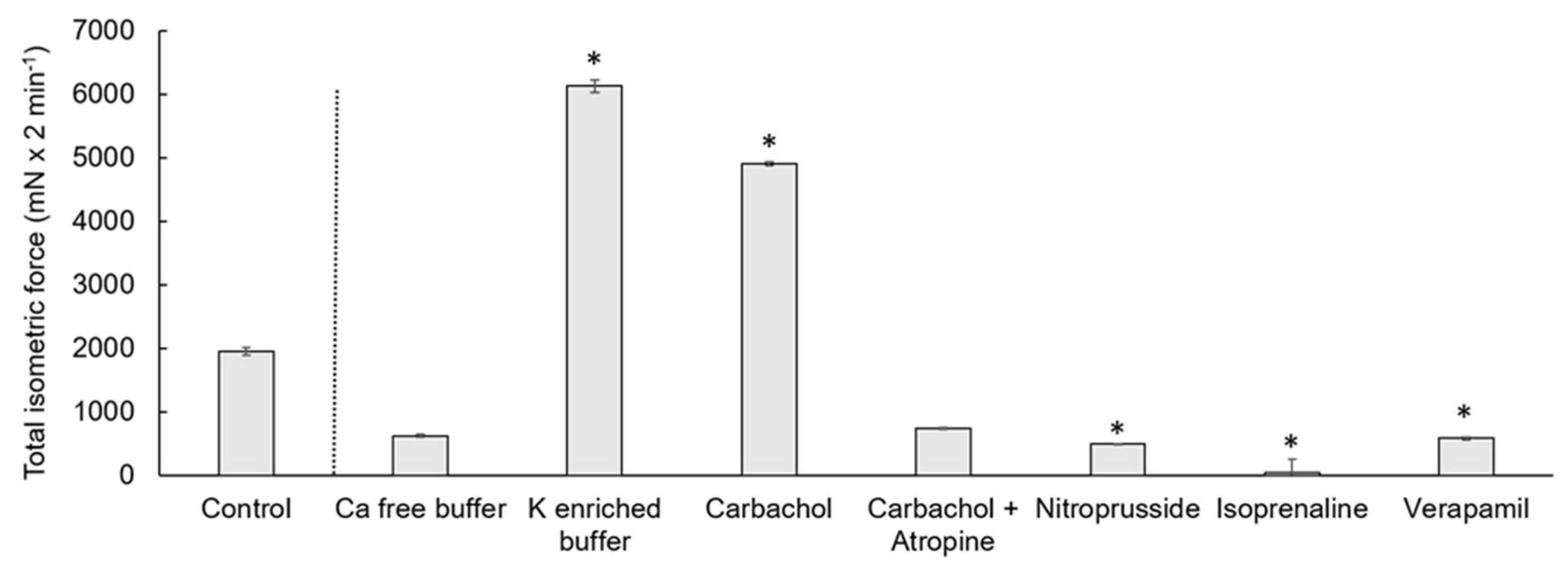
3.1. Weight of the Muscle Strips
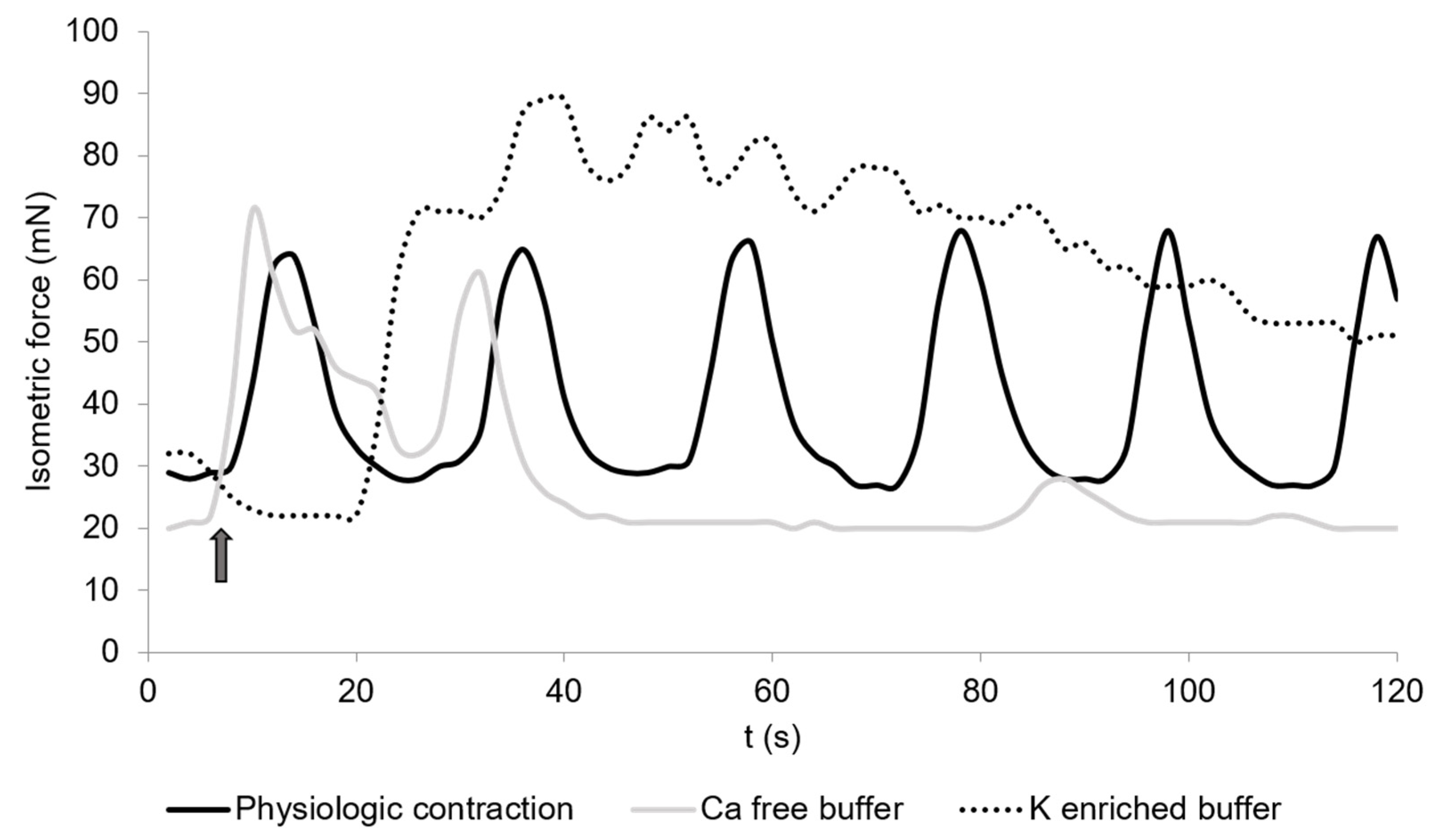
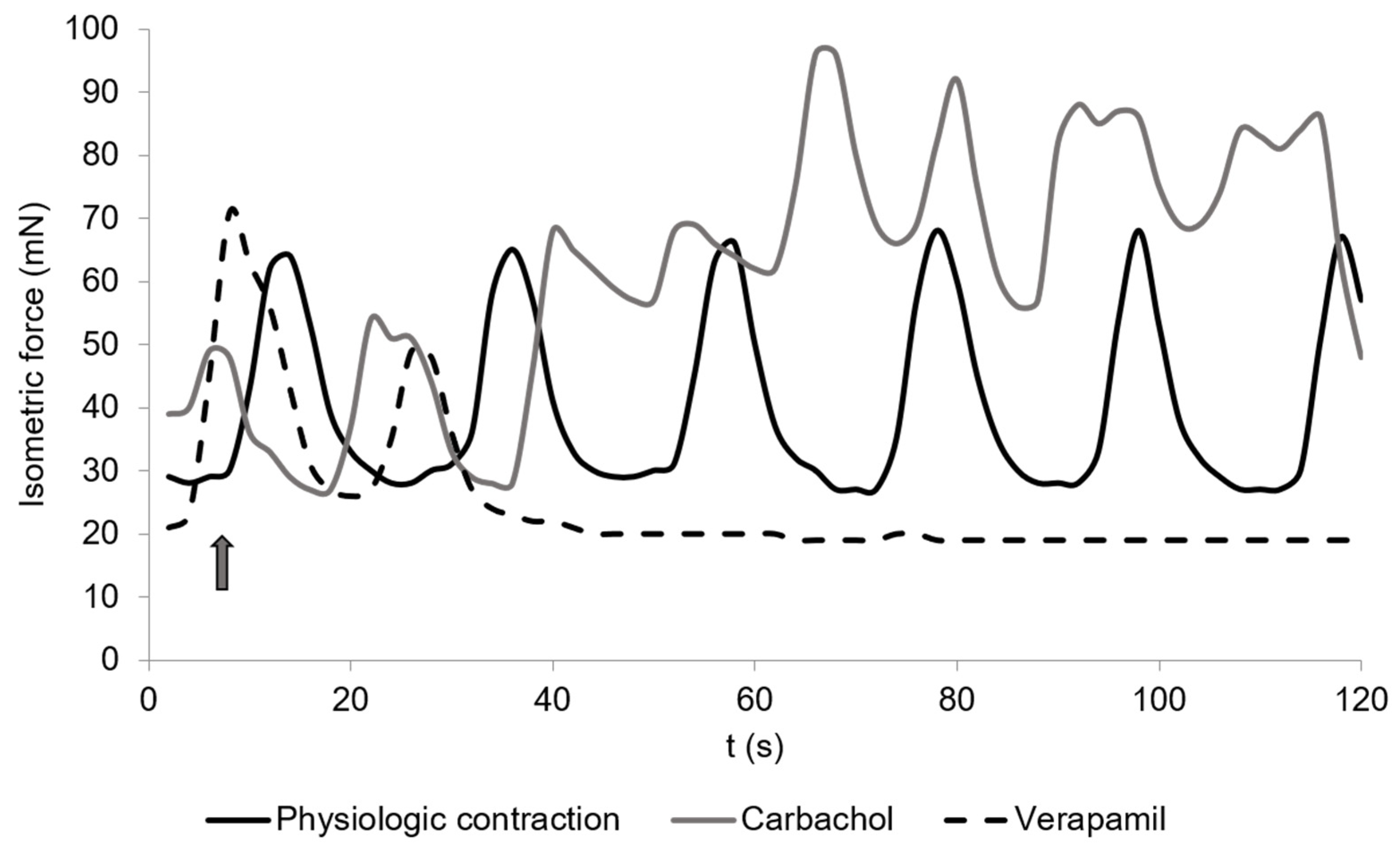
3.2. Spontaneous Contraction
3.3. Calcium-Free Buffer
3.4. Potassium-Enriched Buffer
3.5. Carbachol
3.6. Carbachol and Atropine
3.7. Nitroprusside
3.8. Isoprenaline
3.9. Verapamil
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scanes, C.G.; Pierzchala-Koziec, K. Biology of the Gastrointestinal Tract in Poultry. Avian Biol. Res. 2014, 7, 193–222. [Google Scholar] [CrossRef]
- Gong, J.; Forster, R.J.; Yu, H.; Chambers, J.R.; Sabour, P.M.; Wheatcroft, R.; Chen, S. Diversity and phylogenetic analysis of bacteria in the mucosa of chicken ceca and comparison with bacteria in the cecal lumen. FEMS Microbiol. Lett. 2002, 208, 1–7. [Google Scholar] [CrossRef]
- Yeoman, C.J.; Chia, N.; Jeraldo, P.; Sipos, M.; Goldenfeld, N.D.; White, B.A. The microbiome of the chicken gastrointestinal tract. Anim. Health Res. Rev. 2012, 13, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Son, J.H.; Ragland, D.; Adeola, O. Quantification of digesta flow into the caeca. Br. Poult. Sci. 2002, 43, 322–324. [Google Scholar] [CrossRef]
- Thomas, D.H. Salt and water excretion by birds: The lower intestine as an integrator of renal and intestinal excretion. Comp. Biochem. Physiol. Part A Physiol. 1982, 71, 527–535. [Google Scholar] [CrossRef]
- Thomas, D.H.; Skadhauge, E. Transport function and control in bird caeca. Comp. Biochem. Physiol. Part A Physiol. 1988, 90, 591–596. [Google Scholar] [CrossRef]
- Thomas, D.H.; Skadhauge, E. Functions of the flow of urine and digesta in the avian lower intestine. Acta Vet. Scand. Suppl. 1989, 86, 212–218. [Google Scholar] [PubMed]
- Karasawa, Y.; Son, J.H.; Koh, K. Ligation of caeca improves nitrogen utilisation and decreases urinary uric acid excretion in chickens fed on a low protein diet plus urea. Br. Poult. Sci. 1997, 38, 439–441. [Google Scholar] [CrossRef]
- Duke, G.E. Relationship of cecal and colonic motility to diet, habitat, and cecal anatomy in several avian species. J. Exp. Zool. 1989, 252, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Svihus, B.; Choct, M.; Classen, H. Function and nutritional roles of the avian caeca: A review. World’s Poult. Sci. J. 2013, 69, 249–264. [Google Scholar] [CrossRef]
- Olsson, C.; Holmgren, S. The control of gut motility. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 128, 479–501. [Google Scholar] [CrossRef]
- Sanders, K.M.; Koh, S.D.; Ro, S.; Ward, S.M. Regulation of gastrointestinal motility—Insights from smooth muscle biology. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 633–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vantrappen, G.; Janssens, J.; Hellemans, J.; Ghoos, Y. The interdigestive motor complex of normal subjects and patients with bacterial overgrowth of the small intestine. J. Clin. Investig. 1977, 59, 1158–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieuwenhuijs, V.B.; Verheem, A.; van Duijvenbode-Beumer, H.; Visser, M.R.; Verhoef, J.; Gooszen, H.G.; Akkermans, L.M. The role of interdigestive small bowel motility in the regulation of gut microflora, bacterial overgrowth, and bacterial translocation in rats. Ann. Surg. 1998, 228, 188–193. [Google Scholar] [CrossRef]
- Duke, G.E.; Evanson, O.A.; Epstein, D.R. Coordination of cecal motility during cecal evacuation. Poult. Sci. 1983, 62, 545–550. [Google Scholar] [CrossRef]
- Duke, G.E.; Evanson, O.A.; Huberty, B.J. Electrical potential changes and contractile activity of the distal cecum of turkeys. Poult. Sci. 1980, 59, 1925–1934. [Google Scholar] [CrossRef]
- Hodgkiss, J.P. Peristalsis and antiperistalsis in the chicken caecum are myogenic. Q. J. Exp. Physiol. 1984, 69, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Janssen, P.W.; Lentle, R.G.; Hulls, C.; Ravindran, V.; Amerah, A.M. Spatiotemporal mapping of the motility of the isolated chicken caecum. J. Comp. Physiol. B 2009, 179, 593–604. [Google Scholar] [CrossRef]
- Gabella, G. Structure of the musculature of the chicken small intestine. Anat. Embryol. 1985, 171, 139–149. [Google Scholar] [CrossRef]
- Perrino, B.A. Calcium sensitization mechanisms in gastrointestinal smooth muscles. J. Neurogastroenterol. Motil. 2016, 22, 213–225. [Google Scholar] [CrossRef] [Green Version]
- Bolton, T.B.; Lang, R.J.; Takewaki, T. Mechanisms of action of noradrenaline and carbachol on smooth muscle of guinea-pig anterior mesenteric artery. J. Physiol. 1984, 351, 549–572. [Google Scholar] [CrossRef] [Green Version]
- Hirsbrunner, G.; Knutti, B.; Liu, I.; Küpfer, U.; Scholtysik, G.; Steiner, A. An in vitro study on spontaneous myometrial contractility in the cow during estrus and diestrus. Anim. Reprod. Sci. 2002, 70, 171–180. [Google Scholar] [CrossRef]
- Otto, B.; Steusloff, A.; Just, I.; Aktories, K.; Pfitzer, G. Role of Rho proteins in carbachol-induced contractions in intact and permeabilized guinea-pig intestinal smooth muscle. J. Physiol. 1996, 496, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Nieto, J.E.; Rakestraw, P.C.; Snyder, J.R.; Vatistas, N.J. In vitro effects of erythromycin, lidocaine, and metoclopramide on smooth muscle from the pyloric antrum, proximal portion of the duodenum, and middle portion of the jejunum of horses. Am. J. Vet. Res. 2000, 61, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Tappenbeck, K.; Hoppe, S.; Hopster, K.; Kietzmann, M.; Feige, K.; Huber, K. Lidocaine and structure-related mexiletine induce similar contractility-enhancing effects in ischaemia-reperfusion injured equine intestinal smooth muscle in vitro. Vet. J. 2013, 196, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Tappenbeck, K.; Hoppe, S.; Reichert, C.; Feige, K.; Huber, K. In vitro effects of lidocaine on contractility of circular and longitudinal equine intestinal smooth muscle. Vet. J. 2013, 198, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Ignarro, L.J.; Kadowitz, P.J. The pharmacological and physiological role of cyclic GMP in vascular smooth muscle relaxation. Annu. Rev. Pharmacol. Toxicol. 1985, 25, 171–191. [Google Scholar] [CrossRef]
- Sarna, S.K. Gastrointestinal longitudinal muscle contractions. Am. J. Physiol. Gastrointest. Liver Physiol. 1993, 265, G156–G164. [Google Scholar] [CrossRef] [PubMed]
- Evans, E.D.; Mangel, A.W. Depolarization-stimulated contractility of gastrointestinal smooth muscle in calcium-free solution: A review. ISRN Gastroenterol. 2011, 2011, 692528. [Google Scholar] [CrossRef] [PubMed]
- Hara, Y.; Szurszewski, J.H. Effect of potassium and acetylcholine on canine intestinal smooth muscle. J. Physiol. 1986, 372, 521–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, K.G.; Szurszewski, J.H. Mechanisms of phasic and tonic actions of pentagastrin on canine gastric smooth muscle. J. Physiol. 1980, 301, 229–242. [Google Scholar] [CrossRef] [Green Version]
- Morgan, K.G.; Muir, T.C.; Szurszewski, J.H. The electrical basis for contraction and relaxation in canine fundal smooth muscle. J. Physiol. 1981, 311, 475–488. [Google Scholar] [CrossRef]
- Mitsui, M.; Karaki, H. Dual effects of carbachol on cytosolic Ca2+ and contraction in intestinal smooth muscle. Am. J. Physiol. Cell Physiol. 1990, 258, C787–C793. [Google Scholar] [CrossRef]
- Murthy, K.S. Signaling for contraction and relaxation in smooth muscle of the gut. An. Rev. Physiol. 2006, 68, 345–374. [Google Scholar] [CrossRef]
- Ehlert, F.J. Contractile role of M2 and M3 muscarinic receptors in gastrointestinal, airway and urinary bladder smooth muscle. Life Sci. 2003, 74, 355–366. [Google Scholar] [CrossRef]
- Sanders, K.M.; Ward, S.M. Nitric oxide and its role as a non-adrenergic, non-cholinergic inhibitory neurotransmitter in the gastrointestinal tract. Br. J. Pharmacol. 2019, 176, 212–227. [Google Scholar] [CrossRef] [Green Version]
- Carvajal, J.A.; Germain, A.M.; Huidobro-Toro, J.P.; Weiner, C.P. Molecular mechanism of cGMP-mediated smooth muscle relaxation. J. Cell Physiol. 2000, 184, 409–420. [Google Scholar] [CrossRef]
- Kanagy, N.L.; Charpie, J.R.; Dananberg, J.; Webb, R.C. Decreased sensitivity to vasoconstrictors in aortic rings after acute exposure to nitric oxide. Am. J. Physiol. 1996, 271, H253–H260. [Google Scholar] [CrossRef] [Green Version]
- Onoue, H.; Katusic, Z.S. The effect of 1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one (ODQ) and charybdotoxin (CTX) on relaxations of isolated cerebral arteries to nitric oxide. Brain Res. 1998, 785, 107–113. [Google Scholar] [CrossRef]
- Plane, F.; Hurrell, A.; Jeremy, J.Y.; Garland, C.J. Evidence that potassium channels make a major contribution to SIN-1-evoked relaxation of rat isolated mesenteric artery. Br. J. Pharmacol. 1996, 119, 1557–1562. [Google Scholar] [CrossRef] [Green Version]
- Stuart-Smith, K.; Warner, D.O.; Jones, K.A. The role of cGMP in the relaxation to nitric oxide donors in airway smooth muscle. Eur J. Pharmacol. 1998, 341, 225–233. [Google Scholar] [CrossRef]
- Wong, W.S.; Roman, C.R.; Fleisch, J.H. Differential relaxant responses of guinea-pig lung strips and bronchial rings to sodium nitroprusside: A mechanism independent of cGMP formation. J. Pharm. Pharmacol. 1995, 47, 757–761. [Google Scholar] [CrossRef] [PubMed]
- Jenkinson, D.H.; Morton, I.K.M. The role of α- and β-adrenergic receptors in some actions of catecholamines on intestinal smooth muscle. J. Physiol. 1967, 188, 387–402. [Google Scholar] [CrossRef] [PubMed]
- Beani, L.; Bianchi, C.; Crema, A. The effect of catecholamines and sympathetic stimulation on the release of acetylcholine from the guinea-pig colon. Br. J. Pharmacol. 1969, 36, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Paton, W.D.M.; Vizi, E.S. The inhibitory action of noradrenaline and adrenaline on acetylcholine output by guinea-pig ileum longitudinal muscle strip. Br. J. Pharmacol. 1969, 35, 10–28. [Google Scholar] [CrossRef] [Green Version]
- Bülbring, E.; den Hertog, A. The action of isoprenaline on the smooth muscle of the guinea-pig taenia coli. J. Physiol. 1980, 304, 277–296. [Google Scholar] [CrossRef]
- Groisman, S.D.; Krasilschikov, K.B. Investigation of catecholamine effects on stomach and duodenum motor functions in unanaesthetized dogs. J. Auton. Nerv. Syst. 1986, 17, 33–44. [Google Scholar] [CrossRef]
- Wróbel, J.; Michalska, L. The effect of verapamil on intestinal calcium transport. Eur. J. Pharmacol. 1977, 45, 385–387. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Ca-Free Buffer | K-Enriched Buffer | Carbachol | Carbachol +Atropine | Nitroprusside | Isoprenaline | Verapamil | |
|---|---|---|---|---|---|---|---|---|
| Frequency | 6.2 | 1.3 | 1.5 | 2.2 | 5.6 | 3.5 | 0.8 | 3.9 |
| SEM | 0.7 | 0.3 | 0.2 | 0.9 | 1.1 | 0.8 | 0.4 | 0.8 |
| p–Value | - | 0.0010 | 0.0004 | 0.0049 | n.s. | 0.0349 | 0.0004 | n.s. |
| N | 11 | 7 | 9 | 8 | 4 | 11 | 8 | 9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Röhm, K.; Diener, M.; Huber, K.; Seifert, J. Characterization of Cecal Smooth Muscle Contraction in Laying Hens. Vet. Sci. 2021, 8, 91. https://doi.org/10.3390/vetsci8060091
Röhm K, Diener M, Huber K, Seifert J. Characterization of Cecal Smooth Muscle Contraction in Laying Hens. Veterinary Sciences. 2021; 8(6):91. https://doi.org/10.3390/vetsci8060091
Chicago/Turabian StyleRöhm, Katrin, Martin Diener, Korinna Huber, and Jana Seifert. 2021. "Characterization of Cecal Smooth Muscle Contraction in Laying Hens" Veterinary Sciences 8, no. 6: 91. https://doi.org/10.3390/vetsci8060091
APA StyleRöhm, K., Diener, M., Huber, K., & Seifert, J. (2021). Characterization of Cecal Smooth Muscle Contraction in Laying Hens. Veterinary Sciences, 8(6), 91. https://doi.org/10.3390/vetsci8060091