
Multiplexed Genome Editing for Efficient Phenotypic Screening in Zebrafish

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Synthesis of Cas9 mRNA
2.3. CRISPR-Cas9 Target Selection, Design, and Synthesis
2.4. Embryo Injection
2.5. Genotype Identification
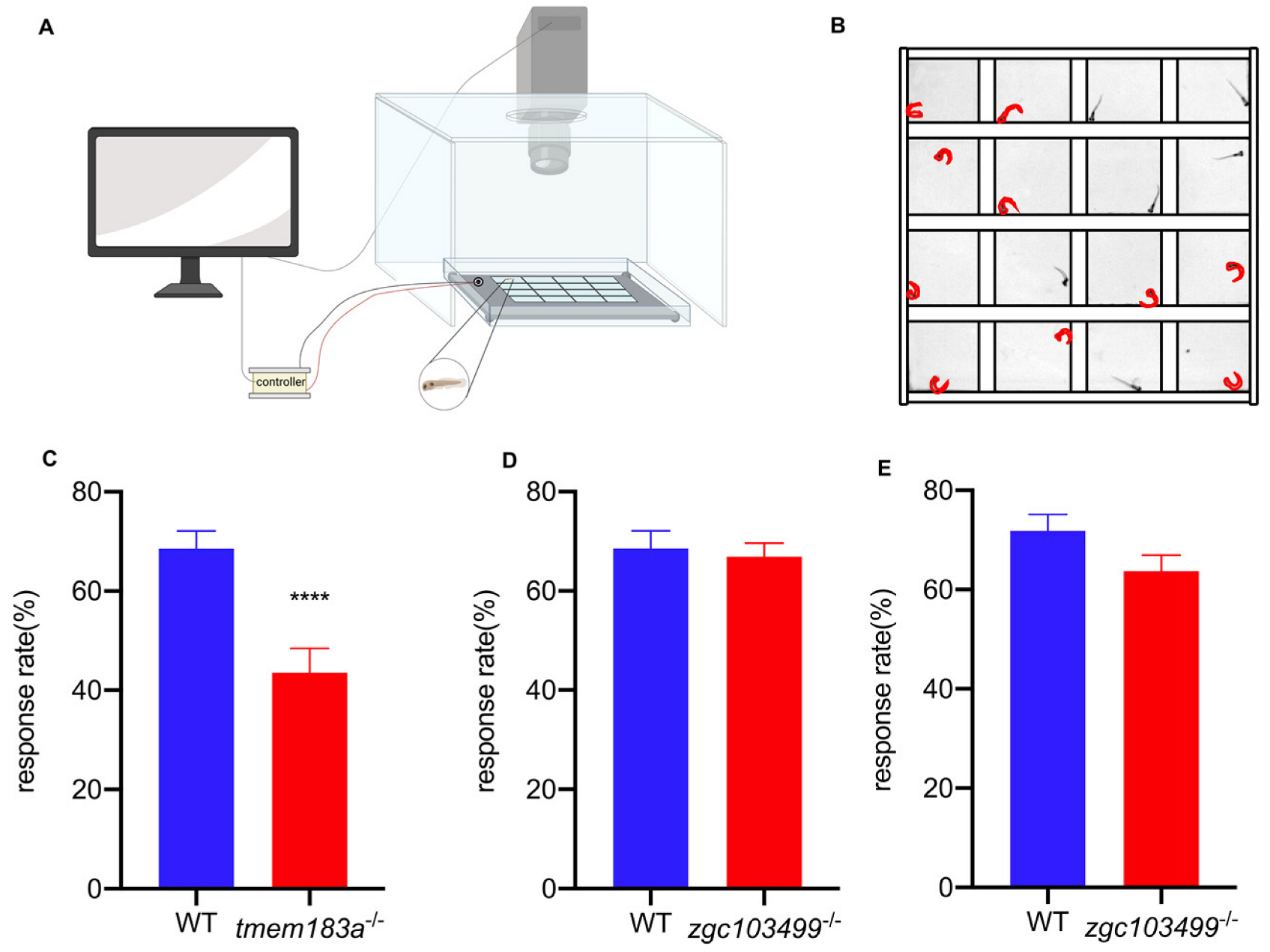
2.6. C-start Response
2.7. Immunohistochemistry
2.8. Statistical Analysis
3. Results
3.1. Design of CRISPR-Cas9 Gene Editing Systems in Candidate Genes
3.2. Multiplexed Genome Editing Led to Five Gene Disruptions in F0 Zebrafish
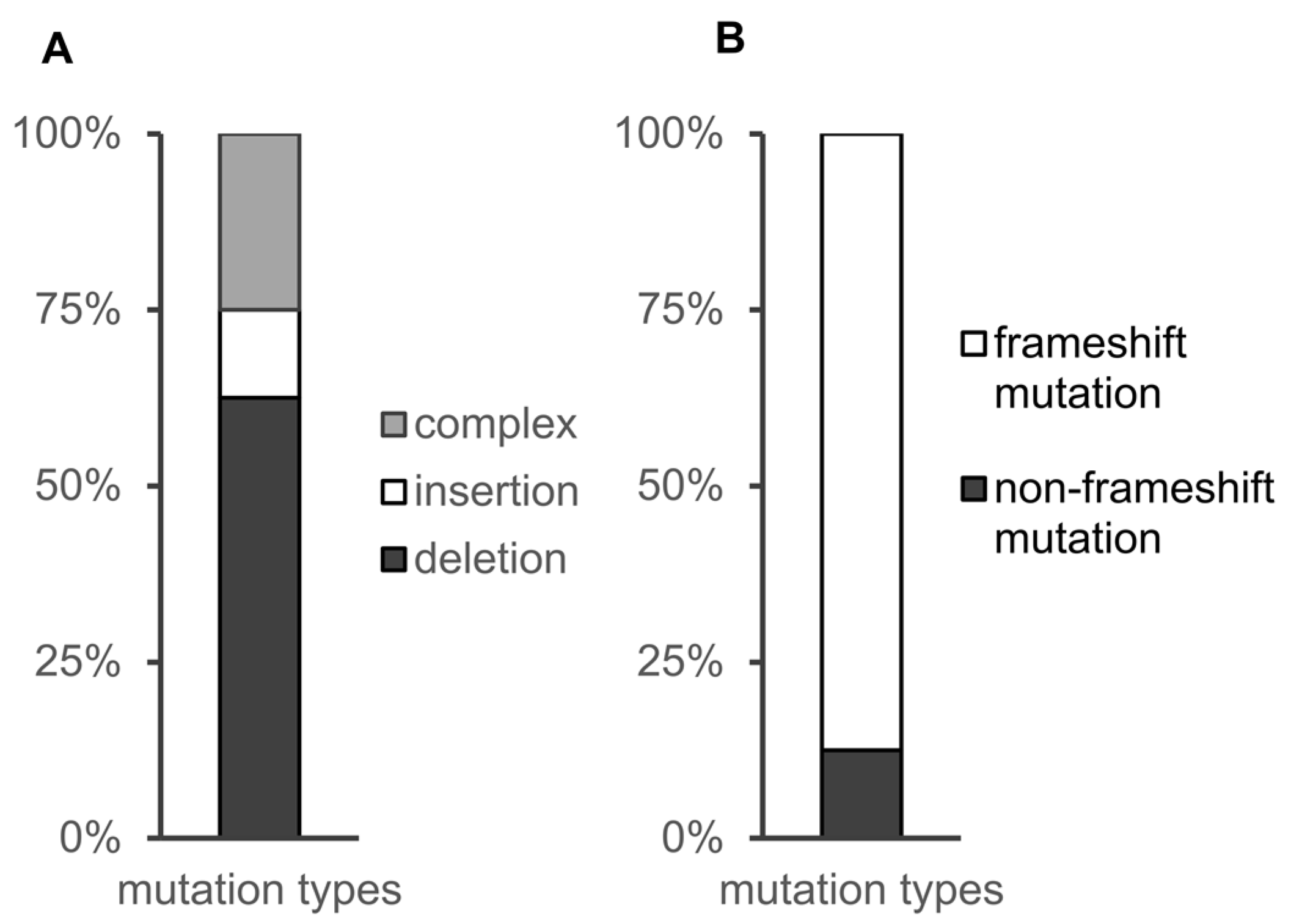
3.3. Distribution of CRISPR-Cas9-Induced Mutations in the Study
3.4. Phenotypic Analysis in Individual Homozygous Mutants
3.4.1. C-start Test
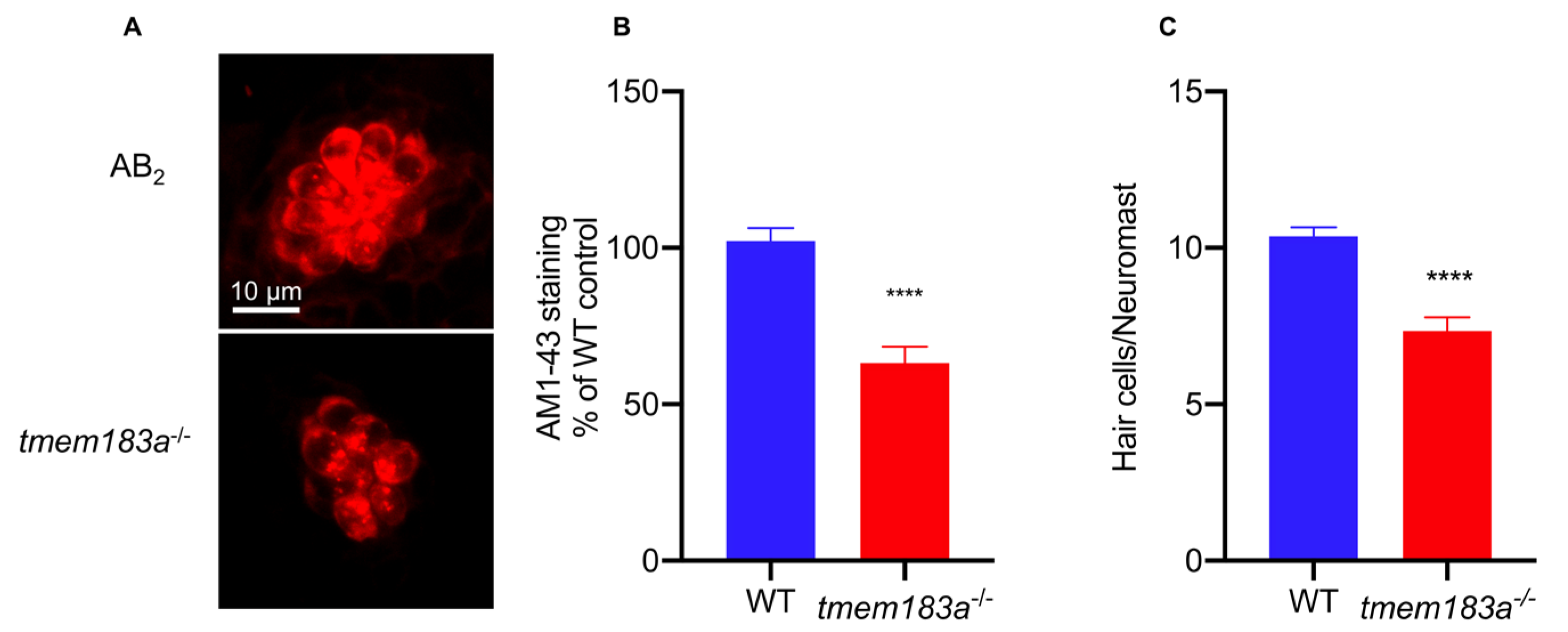
3.4.2. AM1-43 Staining
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Welter, D.; MacArthur, J.; Morales, J.; Burdett, T.; Hall, P.; Junkins, H.; Klemm, A.; Flicek, P.; Manolio, T.; Hindorff, L.; et al. The NHGRI GWAS Catalog, a curated resource of SNP-trait associations. Nucleic Acids Res. 2014, 42, D1001–D1006. [Google Scholar] [CrossRef]
- Choukair, D.; Hauck, F.; Bettendorf, M.; Krude, H.; Klein, C.; Bäumer, T.; Berner, R.; Lee-Kirsch, M.A.; Grasemann, C.; Burgard, P.; et al. An Integrated clinical pathway for diagnosis, treatment and care of rare diseases: Model, operating procedures, and results of the project TRANSLATE-NAMSE funded by the German Federal Joint Committee. Orphanet J. Rare Dis. 2021, 16, 474. [Google Scholar] [CrossRef] [PubMed]
- Antonarakis, S.E. Carrier screening for recessive disorders. Nat. Rev. Genet. 2019, 20, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Stoeger, T.; Gerlach, M.; Morimoto, R.I.; Nunes Amaral, L.A. Large-scale investigation of the reasons why potentially important genes are ignored. PLoS Biol. 2018, 16, e2006643. [Google Scholar] [CrossRef]
- Tang, W.; Davidson, J.D.; Zhang, G.; Conen, K.E.; Fang, J.; Serluca, F.; Li, J.; Xiong, X.; Coble, M.; Tsai, T.; et al. Genetic Control of Collective Behavior in Zebrafish. iScience 2020, 23, 100942–100942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, W.Y.; Fu, Y.; Reyon, D.; Maeder, M.L.; Tsai, S.Q.; Sander, J.D.; Peterson, R.T.; Yeh, J.R.; Joung, J.K. Efficient genome editing in zebrafish using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 227–229. [Google Scholar] [CrossRef]
- McCarty, N.S.; Graham, A.E.; Studená, L.; Ledesma-Amaro, R. Multiplexed CRISPR technologies for gene editing and transcriptional regulation. Nat. Commun. 2020, 11, 1281. [Google Scholar] [CrossRef]
- Chang, N.; Sun, C.; Gao, L.; Zhu, D.; Xu, X.; Zhu, X.; Xiong, J.W.; Xi, J.J. Genome editing with RNA-guided Cas9 nuclease in zebrafish embryos. Cell Res. 2013, 23, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyers, J.R.; MacDonald, R.B.; Duggan, A.; Lenzi, D.; Standaert, D.G.; Corwin, J.T.; Corey, D.P. Lighting up the senses: FM1-43 loading of sensory cells through nonselective ion channels. J. Neurosci. 2003, 23, 4054–4065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Sun, W.; Zhu, X.; Li, L.; Wu, X.; Lin, H.; Zhu, S.; Liu, A.; Du, T.; Liu, Y.; et al. Association between the gamma-aminobutyric acid type B receptor 1 and 2 gene polymorphisms and mesial temporal lobe epilepsy in a Han Chinese population. Epilepsy Res. 2008, 81, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Thuault, S.J.; Brown, J.T.; Sheardown, S.A.; Jourdain, S.; Fairfax, B.; Spencer, J.P.; Restituito, S.; Nation, J.H.; Topps, S.; Medhurst, A.D.; et al. The GABA(B2) subunit is critical for the trafficking and function of native GABA(B) receptors. Biochem. Pharmacol. 2004, 68, 1655–1666. [Google Scholar] [CrossRef] [PubMed]
- Alsahli, S.; Al-Twaijri, W.; Al Mutairi, F. Confirming the pathogenicity of NECAP1 in early onset epileptic encephalopathy. Epilepsia Open 2018, 3, 524–527. [Google Scholar] [CrossRef] [PubMed]
- Mizuguchi, T.; Nakashima, M.; Moey, L.H.; Ch’ng, G.S.; Khoo, T.B.; Mitsuhashi, S.; Miyatake, S.; Takata, A.; Miyake, N.; Saitsu, H.; et al. A novel homozygous truncating variant of NECAP1 in early infantile epileptic encephalopathy: The second case report of EIEE21. J. Hum. Genet. 2019, 64, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Elkon, R.; Milon, B.; Morrison, L.; Shah, M.; Vijayakumar, S.; Racherla, M.; Leitch, C.C.; Silipino, L.; Hadi, S.; Weiss-Gayet, M.; et al. RFX transcription factors are essential for hearing in mice. Nat. Commun. 2015, 6, 8549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popp, M.W.; Maquat, L.E. Leveraging Rules of Nonsense-Mediated mRNA Decay for Genome Engineering and Personalized Medicine. Cell 2016, 165, 1319–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- McCarty, N.S.; Shaw, W.M.; Ellis, T.; Ledesma-Amaro, R. Rapid Assembly of gRNA Arrays via Modular Cloning in Yeast. ACS Synth. Biol. 2019, 8, 906–910. [Google Scholar] [CrossRef] [Green Version]
- Breunig, C.T.; Durovic, T.; Neuner, A.M.; Baumann, V.; Wiesbeck, M.F.; Köferle, A.; Götz, M.; Ninkovic, J.; Stricker, S.H. One step generation of customizable gRNA vectors for multiplex CRISPR approaches through string assembly gRNA cloning (STAgR). PLoS ONE 2018, 13, e0196015. [Google Scholar] [CrossRef]
- Yin, L.; Maddison, L.A.; Li, M.; Kara, N.; LaFave, M.C.; Varshney, G.K.; Burgess, S.M.; Patton, J.G.; Chen, W. Multiplex Conditional Mutagenesis Using Transgenic Expression of Cas9 and sgRNAs. Genetics 2015, 200, 431–441. [Google Scholar] [CrossRef] [Green Version]
- Hwang, W.Y.; Fu, Y.; Reyon, D.; Maeder, M.L.; Kaini, P.; Sander, J.D.; Joung, J.K.; Peterson, R.T.; Yeh, J.R. Heritable and precise zebrafish genome editing using a CRISPR-Cas system. PLoS ONE 2013, 8, e68708. [Google Scholar]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.E.; Norville, J.E.; Church, G.M. RNA-guided human genome engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellard, S.; Shields, B.; Tysoe, C.; Treacy, R.; Yau, S.; Mattocks, C.; Wallace, A. Semi-Automated Unidirectional Sequence Analysis for Mutation Detection in a Clinical Diagnostic Setting. Genet. Test. Mol. Biomark. 2009, 13, 381–386. [Google Scholar] [CrossRef] [Green Version]
- Maier, E.C.; Saxena, A.; Alsina, B.; Bronner, M.E.; Whitfield, T.T. Sensational placodes: Neurogenesis in the otic and olfactory systems. Dev. Biol. 2014, 389, 50–67. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Gao, Y.; Zhang, R.; Sun, F.; Cheng, C.; Qian, F.; Duan, X.; Wei, G.; Sun, C.; Pang, X.; et al. THOC1 deficiency leads to late-onset nonsyndromic hearing loss through p53-mediated hair cell apoptosis. PLoS Genet. 2020, 16, e1008953. [Google Scholar] [CrossRef]
- Pacentine, I.V.; Nicolson, T. Subunits of the mechano-electrical transduction channel, Tmc1/2b, require Tmie to localize in zebrafish sensory hair cells. PLoS Genet. 2019, 15, e1007635. [Google Scholar] [CrossRef] [Green Version]
- Erickson, T.; Morgan, C.P.; Olt, J.; Hardy, K.; Busch-Nentwich, E.; Maeda, R.; Clemens, R.; Krey, J.F.; Nechiporuk, A.; Barr-Gillespie, P.G.; et al. Integration of Tmc1/2 into the mechanotransduction complex in zebrafish hair cells is regulated by Transmembrane O-methyltransferase (Tomt). Elife 2017, 6, e28474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, H.; Kurima, K.; Kawashima, Y.; Griffith, A.J. Mutations of TMC1 cause deafness by disrupting mechanoelectrical transduction. Auris Nasus Larynx 2014, 41, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Kindt, K.S.; Sheets, L. Transmission Disrupted: Modeling Auditory Synaptopathy in Zebrafish. Front. Cell Dev. Biol. 2018, 6, 114. [Google Scholar] [CrossRef]
- Pan, B.; Géléoc, G.S.; Asai, Y.; Horwitz, G.C.; Kurima, K.; Ishikawa, K.; Kawashima, Y.; Griffith, A.J.; Holt, J.R. TMC1 and TMC2 are components of the mechanotransduction channel in hair cells of the mammalian inner ear. Neuron 2013, 79, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Simone, B.W.; Martinez-Galvez, G.; WareJoncas, Z.; Ekker, S.C. Fishing for understanding: Unlocking the zebrafish gene editor’s toolbox. Methods 2018, 150, 3–10. [Google Scholar] [CrossRef]
- Choi, T.Y.; Choi, T.I.; Lee, Y.R.; Choe, S.K.; Kim, C.H. Zebrafish as an animal model for biomedical research. Exp. Mol. Med. 2021, 53, 310–317. [Google Scholar] [CrossRef]
- Abdelrahman, M.; Wei, Z.; Rohila, J.S.; Zhao, K. Multiplex Genome-Editing Technologies for Revolutionizing Plant Biology and Crop Improvement. Front. Plant. Sci. 2021, 12, 721203. [Google Scholar] [CrossRef]
- Kurata, M.; Wolf, N.K.; Lahr, W.S.; Weg, M.T.; Kluesner, M.G.; Lee, S.; Hui, K.; Shiraiwa, M.; Webber, B.R.; Moriarity, B.S. Highly multiplexed genome engineering using CRISPR/Cas9 gRNA arrays. PLoS ONE 2018, 13, e0198714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, J.; Wang, Z.; Zhang, Y.; Shi, S.; Nielsen, J.; Liu, Z. A gRNA-tRNA array for CRISPR-Cas9 based rapid multiplexed genome editing in Saccharomyces cerevisiae. Nat. Commun. 2019, 10, 1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jao, L.-E.; Wente, S.R.; Chen, W. Efficient multiplex biallelic zebrafish genome editing using a CRISPR nuclease system. Proc. Natl. Acad. Sci. USA 2013, 110, 13904–13909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varshney, G.K.; Pei, W.; LaFave, M.C.; Idol, J.; Xu, L.; Gallardo, V.; Carrington, B.; Bishop, K.; Jones, M.; Li, M.; et al. High-throughput gene targeting and phenotyping in zebrafish using CRISPR/Cas9. Genome. Res. 2015, 25, 1030–1042. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Zhang, H.; Gao, J.; Chen, F.; Chen, S.; Zhang, C.; Peng, G. Rapid and accurate synthesis of TALE genes from synthetic oligonucleotides. BioTechniques 2016, 60, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Voigt, C.A. Engineered dCas9 with reduced toxicity in bacteria: Implications for genetic circuit design. Nucleic Acids Res. 2018, 46, 11115–11125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nema, S.; Bhargava, Y. Open-RAC: Open-Design, Recirculating and Auto-Cleaning Zebrafish Maintenance System. Zebrafish 2017, 14, 371–378. [Google Scholar] [CrossRef]
- Naeem, M.; Majeed, S.; Hoque, M.Z.; Ahmad, I. Latest Developed Strategies to Minimize the Off-Target Effects in CRISPR-Cas-Mediated Genome Editing. Cells 2020, 9, 1608. [Google Scholar] [CrossRef]
- Störtz, F.; Minary, P. crisprSQL: A novel database platform for CRISPR/Cas off-target cleavage assays. Nucleic Acids Res. 2021, 49, D855–D861. [Google Scholar] [CrossRef]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef]
- Quick, R.E.; Buck, L.D.; Parab, S.; Tolbert, Z.R.; Matsuoka, R.L. Highly Efficient Synthetic CRISPR RNA/Cas9-Based Mutagenesis for Rapid Cardiovascular Phenotypic Screening in F0 Zebrafish. Front. Cell Dev. Biol. 2021, 9, 735598. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| gabbr1a | CTTGGTGTTGTGGTAGTAGCATTACA | GGGATGTTATGTGTTTATGTCTTACC |
| gabbr2 | GTGGATCTCAGTTGTTCGGTAGC | GTAGATTTTGGTGATGAGTGAGAAGG |
| necap1 | ACCATCACCAAAAAGTGGTATGGCA | CCTGAAGACAGTTATGAAGCTCACGT |
| tmem183a | ACCTTCCTGATCGTCTTCAGCCT | ATTCCCACATCTGCTCTGACCTAC |
| zgc103499 | GAGACACAGAGAGAGAAATTGTTAGAAG | GAGTTTGAGAACCCCCGCTTTAGA |
| Gene | Forward (5′-3′) |
|---|---|
| gabbr1a | GGATGTCCCTTGAGAACGGGAGG |
| gabbr2 | GGCACGGCCTGGACAACAACTGG |
| necap1 | GGAAGTTGGACGCTCCTGACTGG |
| tmem183a | GGTGTAGATTCGGGGAGAGGAGG |
| zgc103499 | GGATCGGAAACCCACCAAGCAGG |
| Gene | Penta KO #1 | Penta KO #2 | Penta KO #3 | Penta KO #4 | Penta KO #5 | Penta KO #6 | Mutation Efficiency |
|---|---|---|---|---|---|---|---|
| gabbr1a | +2 | × | +2 | × | × | −8 | 50% |
| (−2, +4) | (−2, +4) | ||||||
| gabbr2 | × | × | +1 | × | × | × | 16.67% |
| (−2, +3) | |||||||
| necap1 | × | −4 | × | −10 | × | × | 33% |
| tmem183a | −5 | × | × | × | −5 | × | 33% |
| zgc103499 | × | −33 | × | × | × | +1 | 33% |
| Gene | Specific Sequence | Frameshift Mutation Types |
|---|---|---|
| gabbr1a | GGATGTCCCTTGAGAACGGGAGGGTGTCGCTG | Wild-type |
| GGATGTCCCTTGAG--------GGTGTCGCTG | −8 | |
| GGATGTCCCTTGAGAA-----GGAGGGTGGGTGTC | +2 (−5, +7) | |
| gabbr2 | GGCACGGCCTGGACAACAACTGGTACGCG | Wild-type |
| GGCACGGCCTGGACA---CGCGACTGGTACG | +1 (−3, +4) | |
| necap1 | TCCAGTCAGGAGCGTCCAACTTCCAGTCCG | Wild-type |
| TCC----------GTCCAACTTCCAGTCCG | −10 | |
| TCC----AGGAGCGTCCAACTTCCAGTCCG | −4 | |
| tmem183a | CCTCCTCTCCCCGAATCTACACCCACT | Wild-type |
| CCTCC-----CCGAATCTACACCCACT | −5 | |
| zgc103499 | CCTGCTTGGTGGGTTTCCGATCCAGG | Wild-type |
| CCTGCTATGGTGGGTTTCCGATCCAGG | +1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, S.; Gao, G.; Zhang, C.; Peng, G. Multiplexed Genome Editing for Efficient Phenotypic Screening in Zebrafish. Vet. Sci. 2022, 9, 92. https://doi.org/10.3390/vetsci9020092
Guo S, Gao G, Zhang C, Peng G. Multiplexed Genome Editing for Efficient Phenotypic Screening in Zebrafish. Veterinary Sciences. 2022; 9(2):92. https://doi.org/10.3390/vetsci9020092
Chicago/Turabian StyleGuo, Shuyu, Ge Gao, Cuizhen Zhang, and Gang Peng. 2022. "Multiplexed Genome Editing for Efficient Phenotypic Screening in Zebrafish" Veterinary Sciences 9, no. 2: 92. https://doi.org/10.3390/vetsci9020092
APA StyleGuo, S., Gao, G., Zhang, C., & Peng, G. (2022). Multiplexed Genome Editing for Efficient Phenotypic Screening in Zebrafish. Veterinary Sciences, 9(2), 92. https://doi.org/10.3390/vetsci9020092