Live Performance and Microbial Load Modulation of Broilers Fed a Direct-Fed Microbials (DFM) and Xylanase Combination


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Experimental Diets
2.3. Data Collection
2.3.1. Live Performance
2.3.2. Apparent Metabolizable Energy Digestibility
2.3.3. Intestinal Lesion Score
2.3.4. E. tenella Enumeration, Pathogen Load in the Digesta, and Salmonella Incidence
2.3.5. Litter Moisture
2.3.6. Environmental Pathogen Load
2.4. Statistical Methods
3. Results
3.1. Live Performance and Apparent Metabolizable Energy Digestibility
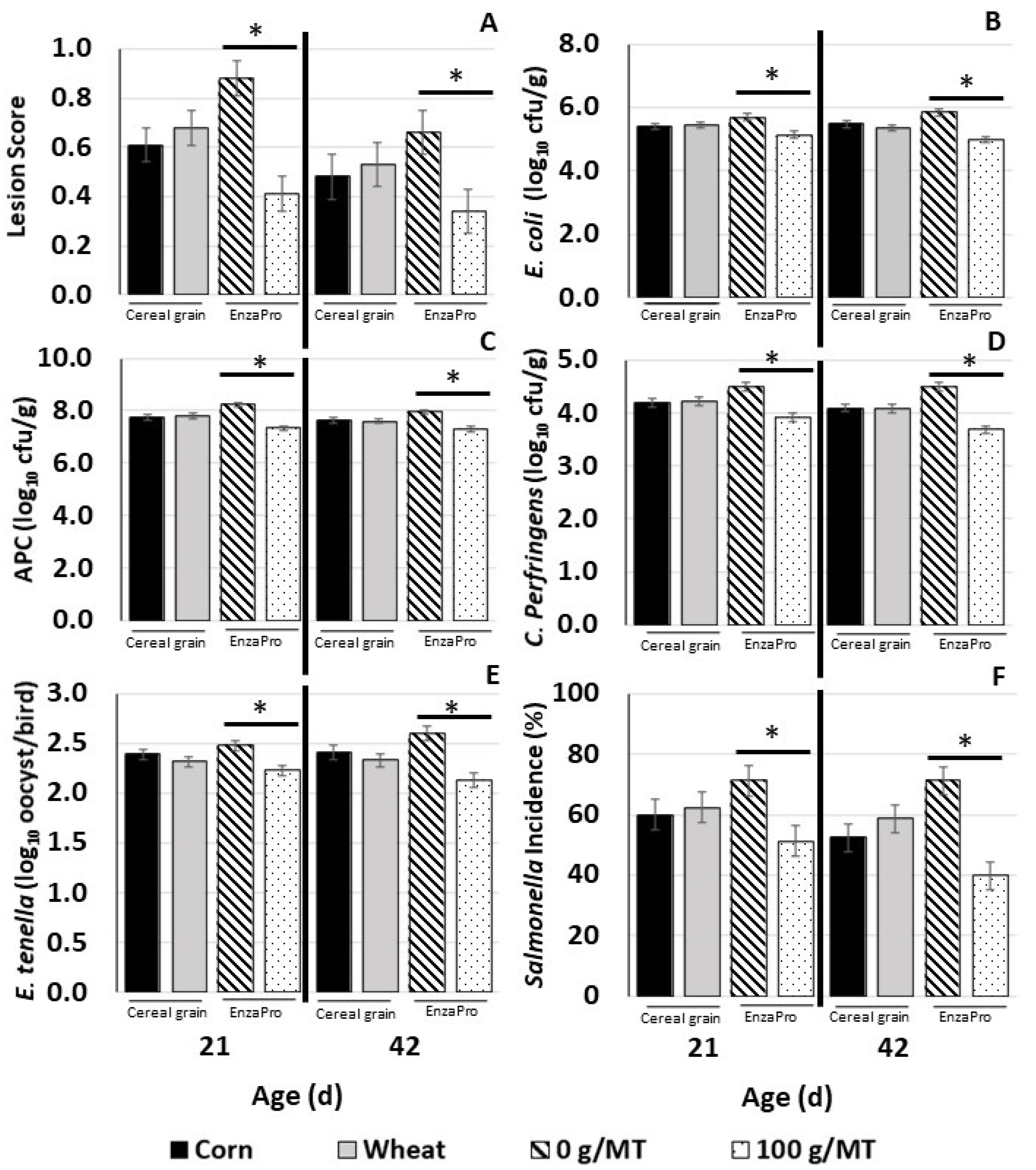
3.2. Intestinal Lesion Score, Pathogen Load in the Digesta, E. tenella Enumeration, and Salmonella Incidence
3.3. Litter Moisture and Environmental Pathogen Load
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ravindran, V.; Abdollahi, M.R. Nutrition and Digestive Physiology of the Broiler Chick: State of the Art and Outlook. Animals 2021, 11, 2795. [Google Scholar] [CrossRef] [PubMed]
- Bedford, M. Mechanism of action and potential environmental benefits from the use of feed enzymes. Anim. Feed Sci. Technol. 1995, 53, 145–155. [Google Scholar] [CrossRef]
- Bedford, M.R.; Schulze, H. Exogenous enzymes for pigs and poultry. Nutr. Res. Rev. 1998, 11, 91–114. [Google Scholar] [CrossRef] [PubMed]
- Choct, M. Enzymes for the feed industry: Past, present and future. World’s Poult. Sci. J. 2006, 62, 5–16. [Google Scholar] [CrossRef]
- Bedford, M.; Partridge, G. Enzymes in Farm Animal Nutrition, 2nd ed.; Wallingford: Oxforshire, UK; CABI: Cambridge, MA, USA, 2010. [Google Scholar]
- Cowieson, A.J. Strategic Selection of Exogenous Enzymes for Corn/soy-based Poultry Diets. J. Poult. Sci. 2010, 47, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Agboola, A.F.; Odu, O.; Omidiwura, B.R.O.; Iyayi, E.A. Effect of Probiotic, Carbohydrase Enzyme and Their Combination on the Performance, Histomorphology and Gut Microbiota in Broilers Fed Wheat-based Diets. Am. J. Exp. Agric. 2015, 8, 307–319. [Google Scholar] [CrossRef]
- Sun, Q.; Liu, D.; Guo, S.; Chen, Y.; Guo, Y. Effects of dietary essential oil and enzyme supplementation on growth performance and gut health of broilers challenged by Clostridium perfringens. Anim. Feed Sci. Technol. 2015, 207, 234–244. [Google Scholar] [CrossRef]
- Flores, C.; Williams, M.; Pieniazek, J.; Dersjant-Li, Y.; Awati, A.; Lee, J.T. Direct-fed microbial and its combination with xylanase, amylase, and protease enzymes in comparison with AGPs on broiler growth performance and foot-pad lesion development. J. Appl. Poult. Res. 2016, 25, 328–337. [Google Scholar] [CrossRef]
- Munyaka, P.M.; Nandha, N.K.; Kiarie, E.; Nyachoti, C.M.; Khafipour, E. Impact of combined β-glucanase and xylanase enzymes on growth performance, nutrients utilization and gut microbiota in broiler chickens fed corn or wheat-based diets. Poult. Sci. 2016, 95, 528–540. [Google Scholar] [CrossRef]
- Stefanello, C.; Vieira, S.L.; Carvalho, P.S.; Sorbara, J.O.B.; Cowieson, A.J. Energy and nutrient utilization of broiler chickens fed corn-soybean meal and corn-based diets supplemented with xylanase. Poult. Sci. 2016, 95, 1881–1887. [Google Scholar] [CrossRef]
- Teymouri, H.; Zarghi, H.; Golian, A. Evaluation of hull-less barley with or without enzyme cocktail in the finisher diets of broiler chickens. J. Agric. Sci. Technol. 2018, 20, 469–483. [Google Scholar]
- Nusairat, B.; Wang, J.-J. Xylanase and Direct-Fed Microbials (DFM) Potential for Improvement of Live Performance, Energy Digestibility, and Reduction of Environmental Microbial Load of Broilers. Front. Vet. Sci. 2020, 7, 606415. [Google Scholar] [CrossRef] [PubMed]
- Nusairat, B.; Wang, J.-J. The Effect of a Modified GH11 Xylanase on Live Performance, Gut Health, and Clostridium perfringens Excretion of Broilers Fed Corn-Soy Diets. Front. Vet. Sci. 2021, 8, 678536. [Google Scholar] [CrossRef]
- Van Hoeck, V.; Wu, D.; Somers, I.; Wealleans, A.; Vasanthakumari, B.; Sanchez, A.G.; Morisset, D. Xylanase impact beyond performance: A prebiotic approach in broiler chickens. J. Appl. Poult. Res. 2021, 30, 100193. [Google Scholar] [CrossRef]
- Ding, X.M.; Li, D.D.; Bai, S.P.; Wang, J.P.; Zeng, Q.F.; Su, Z.W.; Xuan, Y.; Zhang, K.Y. Effect of dietary xylooligosaccharides on intestinal characteristics, gut microbiota, cecal short-chain fatty acids, and plasma immune parameters of laying hens. Poult. Sci. 2018, 97, 874–881. [Google Scholar] [CrossRef]
- Schat, K.A.; Myers, T.J. Avian intestinal immunity. Crit. Rev. Poult. Biol. 1993, 3, 19–34. [Google Scholar]
- Harrow, S.A.; Ravindran, V.; Butler, R.C.; Marshall, J.W.; Tannock, G.W. Real-Time Quantitative PCR Measurement of Ileal Lactobacillus salivarius Populations from Broiler Chickens to Determine the Influence of Farming Practices. Appl. Environ. Microbiol. 2007, 73, 7123–7127. [Google Scholar] [CrossRef] [Green Version]
- Nurmi, E.; Rantala, M. New Aspects of Salmonella Infection in Broiler Production. Nature 1973, 241, 210–211. [Google Scholar] [CrossRef]
- Khaksefidi, A.; Ghoorchi, T. Effect of Probiotic on Performance and Immunocompetence in Broiler Chicks. J. Poult. Sci. 2006, 43, 296–300. [Google Scholar] [CrossRef] [Green Version]
- Willis, W.L.; Isikhuemhen, O.S.; Ibrahim, S.A. Performance Assessment of Broiler Chickens Given Mushroom Extract Alone or in Combination with Probiotics. Poult. Sci. 2007, 86, 1856–1860. [Google Scholar] [CrossRef]
- Mingmongkolchai, S.; Panbangred, W. Bacillusprobiotics: An alternative to antibiotics for livestock production. J. Appl. Microbiol. 2018, 124, 1334–1346. [Google Scholar] [CrossRef]
- Xu, S.; Lin, Y.; Zeng, D.; Zhou, M.; Zeng, Y.; Wang, H.; Zhou, Y.; Zhu, H.; Pan, K.; Jing, B.; et al. Bacillus licheniformis normalize the ileum microbiota of chickens infected with necrotic enteritis. Sci. Rep. 2018, 8, 1744. [Google Scholar] [CrossRef]
- FASS. Guide for the Care and Use of Agricultural Animals in Research and Teaching, 4th ed.; Poultry Science Association: Champaign, IL, USA, 2020. [Google Scholar]
- NRC. Nutrient Requirements of Poultry; The National Academies Press: Washington, DC, USA, 1994. [Google Scholar]
- Aviagen. Ross Broiler Nutrition Specifications. 2019. Available online: https://en.aviagen.com/assets/Tech_Center/Ross_Broiler/RossBroilerNutritionSpecs-EN.pdf (accessed on 29 December 2021).
- Johnson, J.; Reid, W. Anticoccidial drugs: Lesion scoring techniques in battery and floor-pen experiments with chickens. Exp. Parasitol. 1970, 28, 30–36. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis, 18th ed.; AOAC Int.: Gaithersburg, MD, USA, 2005. [Google Scholar]
- BAM. BAM Chapter 16: Clostridium perfringens. In Bacteriological Analytical Manual, 8th ed.; FDA: GSilver Spring, ML, USA, 1980. [Google Scholar]
- Tang, D.F.; Liu, X.X.; Shi, X.G.; Aftab, U. Effect of cereal type and Xylanase supplementation on nutrient retention and growth performance of broilers. J. Appl. Poult. Res. 2017, 26, 529–535. [Google Scholar] [CrossRef]
- Farahat, M.; Ibrahim, D.; Kishawy, A.; Abdallah, H.; Hernandez-Santana, A.; Attia, G. Effect of cereal type and plant extract addition on the growth performance, intestinal morphology, caecal microflora, and gut barriers gene expression of broiler chickens. Animal 2021, 15, 100056. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.V.; Ravindran, V. Effect of Cereal Type on the Performance, Gastrointestinal Tract Development and Intestinal Morphology of the Newly Hatched Broiler Chick. J. Poult. Sci. 2008, 45, 46–50. [Google Scholar] [CrossRef] [Green Version]
- Teirlynck, E.; Bjerrum, L.; Eeckhaut, V.; Huygebaert, G.; Pasmans, F.; Haesebrouck, F.; Dewulf, J.; Ducatelle, R.; Van Immerseel, F. The cereal type in feed influences gut wall morphology and intestinal immune cell infiltration in broiler chickens. Br. J. Nutr. 2009, 102, 1453–1461. [Google Scholar] [CrossRef]
- Nusairat, B.; McNaughton, J.; Tyus, J.; Wang, J.-J. Combination of Xylanase and Bacillus Direct-fed Microbials, as an Alternative to Antibiotic Growth Promoters, Improves Live Performance and Gut Health in Subclinical Challenged Broilers. Int. J. Poult. Sci. 2018, 17, 362–366. [Google Scholar] [CrossRef]
- Vandeplas, S.; Dauphin, R.D.; Thiry, C.; Beckers, Y.; Welling, G.W.; Thonart, P.; Théwis, A. Efficiency of a Lactobacillus plantarum-xylanase combination on growth performances, microflora populations, and nutrient digestibilities of broilers infected with Salmonella Typhimurium. Poult. Sci. 2009, 88, 1643–1654. [Google Scholar] [CrossRef]
- De Oliveira, M.J.K.; Sakomura, N.K.; Dorigam, J.C.D.P.; Doranalli, K.; Soares, L.; Viana, G. Bacillus amyloliquefaciens CECT 5940 alone or in combination with antibiotic growth promoters improves performance in broilers under enteric pathogen challenge. Poult. Sci. 2019, 98, 4391–4400. [Google Scholar] [CrossRef]
- Singh, A.K.; Mishra, B.; Bedford, M.R.; Jha, R. Effects of supplemental xylanase and xylooligosaccharides on production performance and gut health variables of broiler chickens. J. Anim. Sci. Biotechnol. 2021, 12, 98. [Google Scholar] [CrossRef]
- Wecke, C.; Liebert, F. Age and Gender Dependent Deposition of Crude Nutrients and Energy in Fast Growing Meat-Type Chickens. Open J. Anim. Sci. 2019, 09, 35–50. [Google Scholar] [CrossRef] [Green Version]
- Choct, M.; Hughes, R.J.; Wang, J.; Bedford, M.R.; Morgan, A.J.; Annison, G. Increased small intestinal fermentation is partly responsible for the anti-nutritive activity of non-starch polysaccharides in chickens. Br. Poult. Sci. 1996, 37, 609–621. [Google Scholar] [CrossRef]
- Cowieson, A.; O’Neill, H.M.; O’Neill, H.M. Effects of exogenous xylanase on performance, nutrient digestibility and caecal thermal profiles of broilers given wheat-based diets. Br. Poult. Sci. 2013, 54, 346–354. [Google Scholar] [CrossRef]
- Hosseini, S.M.; Afshar, M. Effects of feed form and xylanase supplementation on performance and ileal nutrients digestibility of heat-stressed broilers fed wheat–soybean diet. J. Appl. Anim. Res. 2017, 45, 550–556. [Google Scholar] [CrossRef]
- Lee, S.H.; Hosseindoust, A.; Ingale, S.L.; Rathi, P.C.; Yoon, S.Y.; Choi, J.W.; Kim, J.S. Thermostable xylanase derived from Trichoderma citrinoviride increases growth performance and non-starch polysaccharide degradation in broiler chickens. Br. Poult. Sci. 2020, 61, 57–62. [Google Scholar] [CrossRef]
- Attia, Y.A.; Al-Khalaifah, H.; El-Hamid, H.S.A.; Al-Harthi, M.A.; El-Shafey, A.A. Effect of Different Levels of Multienzymes on Immune Response, Blood Hematology and Biochemistry, Antioxidants Status and Organs Histology of Broiler Chicks Fed Standard and Low-Density Diets. Front. Vet. Sci. 2020, 6, 510. [Google Scholar] [CrossRef] [Green Version]
- Kouzounis, D.; Hageman, J.; Soares, N.; Michiels, J.; Schols, H. Impact of Xylanase and Glucanase on Oligosaccharide Formation, Carbohydrate Fermentation Patterns, and Nutrient Utilization in the Gastrointestinal Tract of Broilers. Animals 2021, 11, 1285. [Google Scholar] [CrossRef]
- Paraskeuas, V.; Mountzouris, K.C. Broiler gut microbiota and expressions of gut barrier genes affected by cereal type and phytogenic inclusion. Anim. Nutr. 2019, 5, 22–31. [Google Scholar] [CrossRef]
- Watkins, B.; Kratzer, F. Effect of Oral Dosing of Lactobacillus Strains on Gut Colonization and Liver Biotin in Broiler Chicks. Poult. Sci. 1983, 62, 2088–2094. [Google Scholar] [CrossRef]
- Francis, C.; Janky, D.; Arafa, A.; Harms, R. Interrelationship of Lactobacillus and Zinc Bacitracin in the Diets of Turkey Poults. Poult. Sci. 1978, 57, 1687–1689. [Google Scholar] [CrossRef]
- Kizerwetter-Swida, M.; Binek, M. Protective effect of potentially probiotic Lactobacillus strain on infection with pathogenic bacteria in chickens. Pol. J. Vet. Sci. 2009, 12, 15–20. [Google Scholar]
- Taira, K.; Nagai, T.; Obi, T.; Takase, K. Effect of Litter Moisture on the Development of Footpad Dermatitis in Broiler Chickens. J. Vet. Med Sci. 2014, 76, 583–586. [Google Scholar] [CrossRef] [Green Version]
- Dumas, M.D.; Polson, S.W.; Ritter, D.; Ravel, J.; Gelb, J.; Morgan, R.; Wommack, K.E. Impacts of Poultry House Environment on Poultry Litter Bacterial Community Composition. PLoS ONE 2011, 6, e24785. [Google Scholar] [CrossRef] [Green Version]
- Moran, E.T. Intestinal events and nutritional dynamics predispose Clostridium perfringens virulence in broilers. Poult. Sci. 2014, 93, 3028–3036. [Google Scholar] [CrossRef]
- Wigley, P. Blurred Lines: Pathogens, Commensals, and the Healthy Gut. Front. Vet. Sci. 2015, 2, 40. [Google Scholar] [CrossRef] [Green Version]
- Shehata, A.A.; Yalçın, S.; Latorre, J.D.; Basiouni, S.; Attia, Y.A.; El-Wahab, A.A.; Visscher, C.; El-Seedi, H.R.; Huber, C.; Hafez, H.M.; et al. Probiotics, Prebiotics, and Phytogenic Substances for Optimizing Gut Health in Poultry. Microorganisms 2022, 10, 395. [Google Scholar] [CrossRef]
- Juricova, H.; Matiasovicova, J.; Faldynova, M.; Sebkova, A.; Kubasova, T.; Prikrylova, H.; Karasova, D.; Crhanova, M.; Havlickova, H.; Rychlik, I. Probiotic Lactobacilli Do Not Protect Chickens against Salmonella Enteritidis Infection by Competitive Exclusion in the Intestinal Tract but in Feed, Outside the Chicken Host. Microorganisms 2022, 10, 219. [Google Scholar] [CrossRef]


{kind=link}
| Ingredient (%) | Starter (1–21 d) | Grower (22–35 d) | Finisher (36–42 d) | |||
|---|---|---|---|---|---|---|
| Corn | Wheat | Corn | Wheat | Corn | Wheat | |
| Corn | 58.16 | 46.32 | 64.50 | 52.67 | 68.94 | 57.14 |
| Wheat | - | 15.00 | - | 15.00 | - | 15.00 |
| Soybean meal 48% | 36.30 | 32.62 | 29.49 | 25.78 | 23.34 | 19.63 |
| Poultry meal | 0.71 | 1.15 | 1.73 | 2.17 | 3.83 | 4.26 |
| Poultry fat | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| DL-Methionine | 0.25 | 0.23 | 0.19 | 0.18 | 0.08 | 0.06 |
| Salt | 0.49 | 0.49 | 0.45 | 0.44 | 0.39 | 0.38 |
| Lysine | 0.00 | 0.09 | 0.04 | 0.14 | 0.03 | 0.13 |
| Limestone | 1.60 | 1.59 | 1.32 | 1.32 | 1.22 | 1.21 |
| Dicalcium phosphate | 1.89 | 1.91 | 1.68 | 1.70 | 1.57 | 1.59 |
| Vitamin and mineral premix 1 | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 |
| Sand filler 2 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| Calculated nutrients (%) | ||||||
| Metabolizable energy (kcal/kg) | 2900 | 2900 | 3000 | 3000 | 3100 | 3100 |
| Crude protein | 22 | 22 | 20 | 20 | 19 | 19 |
| Crude fat | 1.44 | 1.39 | 1.87 | 1.84 | 1.81 | 2.12 |
| Crude fiber | 2.62 | 2.68 | 2.59 | 2.64 | 2.55 | 2.60 |
| Ash | 7.84 | 8.81 | 7.46 | 7.55 | 8.77 | 7.31 |
| Calcium | 1.05 | 1.05 | 0.9 | 0.9 | 0.85 | 0.85 |
| Available phosphate | 0.50 | 0.50 | 0.45 | 0.45 | 0.42 | 0.42 |
| Sodium | 0.22 | 0.22 | 0.2 | 0.2 | 0.18 | 0.18 |
| Dig Lysine | 1.28 | 1.28 | 1.15 | 1.15 | 1.02 | 1.02 |
| Dig Methionine + cysteine | 0.947 | 0.947 | 0.851 | 0.851 | 0.755 | 0.755 |
| Analyzed nutrients (%) | ||||||
| Crude protein | 22.8 | 21.3 | 18.9 | 18.9 | 17.8 | 17.6 |
| Crude fat | 1.70 | 1.63 | 1.57 | 1.44 | 1.67 | 1.77 |
| Crude fiber | 3.01 | 2.84 | 2.87 | 2.99 | 2.72 | 2.58 |
| Ash | 6.55 | 6.58 | 6.10 | 6.54 | 7.95 | 7.35 |
| Dietary Treatments | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|
| Cereal Grain | EnzaPro (g/MT) | SEM 1 | Cereal grain | EnzaPro | Cereal grain x EnzaPro | |||
| Age Period, d | Corn | Wheat | 0 | 100 | ||||
| FI, g/bird | ||||||||
| 1–21 | 1042 | 1044 | 1039 | 1047 | 4.11 | 0.834 | 0.347 | 0.883 |
| 22–35 | 2466 | 2484 | 2471 | 2479 | 10.3 | 0.388 | 0.702 | 0.333 |
| 36–42 | 1383 | 1360 | 1363 | 1379 | 18.4 | 0.552 | 0.673 | 0.468 |
| 1–42 | 4890 | 4888 | 4873 | 4905 | 14.9 | 0.934 | 0.298 | 0.787 |
| BW, g/bird | ||||||||
| Day 0 | 47.7 | 47.5 | 47.8 | 47.4 | 0.162 | 0.698 | 0.241 | 0.435 |
| Day 21 | 838 | 837 | 832 b | 843 a | 2.85 | 0.843 | 0.0479 | 0.894 |
| Day 35 | 2137 | 2133 | 2117 B | 2153 A | 5.67 | 0.718 | 0.0008 | 0.916 |
| Day 42 | 2815 | 2813 | 2788 B | 2841 A | 7.86 | 0.841 | 0.0004 | 0.908 |
| BWG, g/bird | ||||||||
| 1–21 | 791 | 790 | 784 b | 796 a | 2.83 | 0.859 | 0.0395 | 0.856 |
| 22–35 | 1299 | 1296 | 1285 b | 1310 a | 5.75 | 0.822 | 0.0298 | 0.872 |
| 36–42 | 679 | 680 | 671 | 687 | 8.53 | 0.961 | 0.363 | 0.975 |
| 1–42 | 2768 | 2765 | 2740 B | 2793 A | 7.86 | 0.848 | 0.0004 | 0.923 |
| FCR, g:g | ||||||||
| 1–21 | 1.32 | 1.32 | 1.32 | 1.31 | 0.0024 | 0.354 | 0.0368 | 0.756 |
| 22–35 | 1.80 | 1.82 | 1.83 | 1.80 | 0.0079 | 0.198 | 0.101 | 0.436 |
| 36–42 | 1.87 | 1.84 | 1.87 | 1.85 | 0.0268 | 0.601 | 0.771 | 0.547 |
| 1–42 | 1.72 | 1.72 | 1.73 a | 1.71 b | 0.0044 | 0.642 | 0.0282 | 0.822 |
| Mortality, % | ||||||||
| 1–21 | 1.44 | 1.54 | 1.35 | 1.63 | 0.253 | 0.855 | 0.583 | 0.583 |
| 22–35 | 0.240 | 0.350 | 0.335 | 0.255 | 0.0865 | 0.541 | 0.656 | 0.656 |
| 36–42 | 0.210 | 0.105 | 0.210 | 0.105 | 0.0885 | 0.567 | 0.567 | 0.567 |
| 1–42 | 1.87 | 1.98 | 1.87 | 1.98 | 0.283 | 0.859 | 0.860 | 0.601 |
| BW Coefficient of variation, % | ||||||||
| 21 | 13.7 | 14.1 | 13.9 | 13.9 | 0.144 | 0.255 | 0.945 | 0.858 |
| 35 | 8.95 | 8.85 | 8.88 | 8.93 | 0.0885 | 0.585 | 0.788 | 0.564 |
| 42 | 9.30 | 9.12 | 9.48 | 9.04 | 0.0966 | 0.630 | 0.0251 | 0.934 |
| Dietary Treatments | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|
| Cereal Grain | EnzaPro (g/MT) | SEM 1 | Cereal Grain | EnzaPro | Cereal Grain x EnzaPro | |||
| Corn | Wheat | 0 | 100 | |||||
| AME (kcal/kg) | ||||||||
| Day 21 | 2923 | 2922 | 2898 B | 2947 A | 5.03 | 0.818 | 0.0001 | 0.951 |
| Day 42 | 2892 | 2890 | 2866 B | 2916 A | 4.72 | 0.801 | 0.0001 | 0.937 |
| AMEn (kcal/kg) | ||||||||
| Day 21 | 3121 | 3119 | 3096 B | 3143 A | 5.14 | 0.764 | 0.0001 | 0.989 |
| Day 42 | 3091 | 3089 | 3067 B | 3113 A | 4.70 | 0.679 | 0.0001 | 0.883 |
| Dietary Treatments | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|
| Cereal Grain | EnzaPro (g/MT) | SEM 1 | Cereal Grain | EnzaPro | Cereal Grain x EnzaPro | |||
| Corn | Wheat | 0 | 100 | |||||
| Litter moisture, % | ||||||||
| Day 0 | 18.5 | 18.8 | 18.7 | 18.6 | 0.132 | 0.237 | 0.501 | 0.885 |
| Day 21 | 22.9 | 22.9 | 22.9 | 23.0 | 0.129 | 0.916 | 0.896 | 0.697 |
| Day 42 | 26.9 | 27.3 | 27.5 a | 26.6 b | 0.211 | 0.311 | 0.0307 | 0.889 |
| Litter Clostridia perfringens, log10 CFU/g | ||||||||
| Day 0 | 4.36 | 4.18 | 4.19 | 4.35 | 0.114 | 0.424 | 0.496 | 0.967 |
| Day 21 | 4.17 | 4.04 | 4.31 | 3.90 | 0.107 | 0.542 | 0.0601 | 0.319 |
| Day 42 | 3.56 | 4.08 | 4.16 | 3.88 | 0.0939 | 0.544 | 0.147 | 0.811 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nusairat, B.; Odetallah, N.; Wang, J.-J. Live Performance and Microbial Load Modulation of Broilers Fed a Direct-Fed Microbials (DFM) and Xylanase Combination. Vet. Sci. 2022, 9, 142. https://doi.org/10.3390/vetsci9030142
Nusairat B, Odetallah N, Wang J-J. Live Performance and Microbial Load Modulation of Broilers Fed a Direct-Fed Microbials (DFM) and Xylanase Combination. Veterinary Sciences. 2022; 9(3):142. https://doi.org/10.3390/vetsci9030142
Chicago/Turabian StyleNusairat, Basheer, Nasser Odetallah, and Jeng-Jie Wang. 2022. "Live Performance and Microbial Load Modulation of Broilers Fed a Direct-Fed Microbials (DFM) and Xylanase Combination" Veterinary Sciences 9, no. 3: 142. https://doi.org/10.3390/vetsci9030142
APA StyleNusairat, B., Odetallah, N., & Wang, J. -J. (2022). Live Performance and Microbial Load Modulation of Broilers Fed a Direct-Fed Microbials (DFM) and Xylanase Combination. Veterinary Sciences, 9(3), 142. https://doi.org/10.3390/vetsci9030142