
Histological Study of Glandular Variability in the Skin of the Natterjack Toad—Epidalea calamita (Laurenti, 1768)—Used in Spanish Historical Ethnoveterinary Medicine and Ethnomedicine

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Medicinal and Veterinary Practices Based on Epidalea Calamita
1.2. Bioactive Constituents and Candidate Drugs
1.3. A New Ethnopharmacological View through the Integument
2. Materials and Methods
3. Results and Discussion
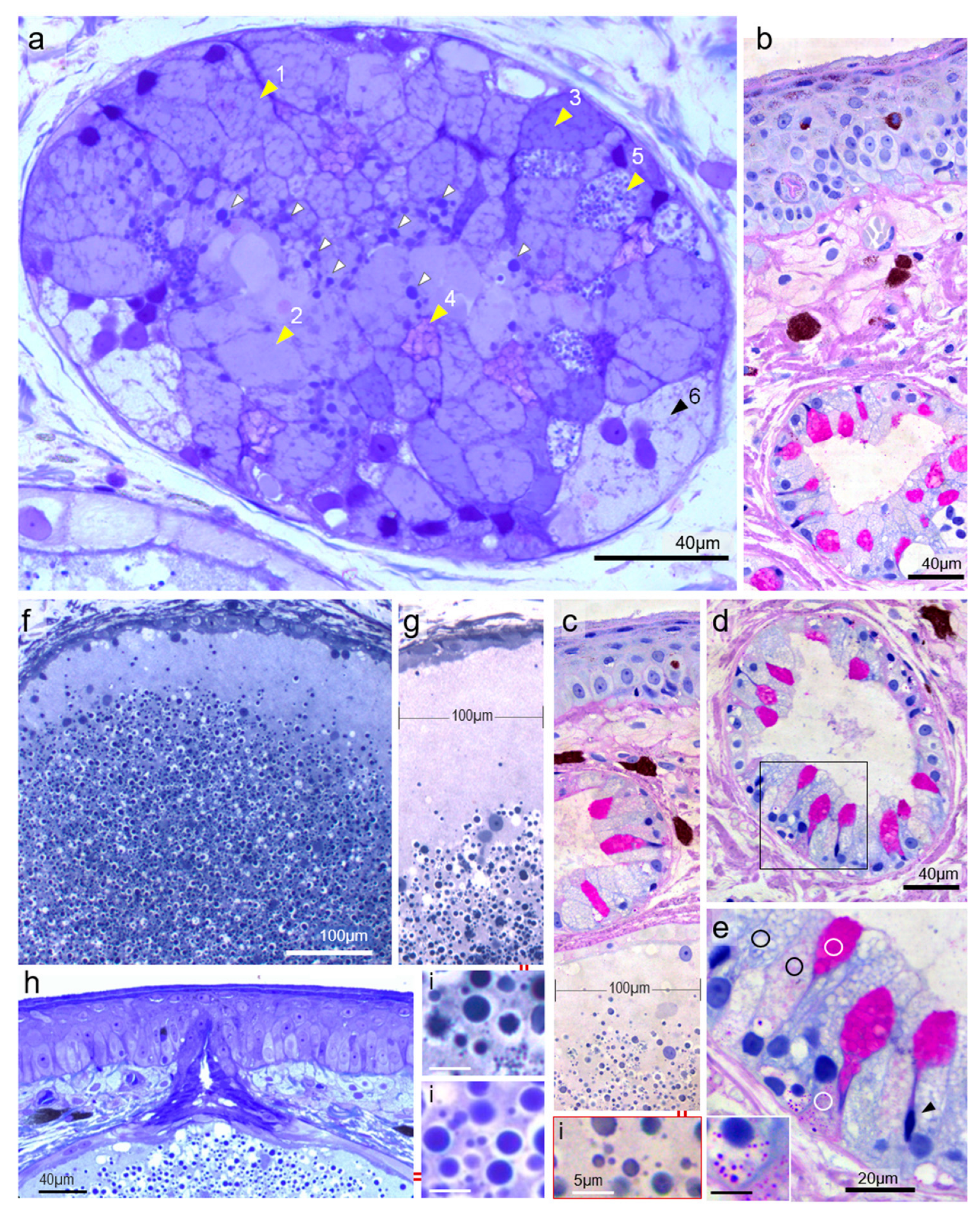
3.1. Tegumentary Architecture
3.2. Mucous Gland Architecture and Cytology
3.3. Variability and Functional Secretory Cycles
3.4. Venom Gland Architecture
3.5. Histogenetic and Evolutionary Basis of Variability
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adams, F. The Genuine Works of Hippocrates; Sydenham Society: London, UK, 1849; Volume 1. [Google Scholar]
- Schawe, C.W. Veterinary Medicine and Human Health; Bailliere, Tindall and Cassell: London, UK, 1969. [Google Scholar]
- Wilkenson, L. Animals and Disease: An Introduction to the History of Comparative Medicine; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Cam, M.-T. (Ed.) La Médecine Vétérinaire Antique: Sources Écrites, Archéologiques, Iconographiques; Presses Universitaires: Rennes, France, 2007. [Google Scholar]
- Teixidó Gómez, F.; Teixidó Gómez, J. Las Obras de Albeytería de Martín Arredondo. Asclepio 2002, 54, 165–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mañé Seró, M.C. Martín Arredondo (1598–16??). In Semblanzas Veterinarias Volume III; Dehesa, F.L., Castaño, M., Etxaniz, J.M., Moreno, L.A., Pumarola, M., Sánchez de Lollano, J., Eds.; Consejo General de Colegios Veterinarios de España: Bilbao, Spain, 2011; pp. 17–28. [Google Scholar]
- Kahn, L.H.; Kaplan, B.; Steele, J.H. Confronting zoonoses through closer collaboration between medicine and veterinary medicine (as ‘one medicine’). Vet. Ital. 2007, 43, 5–19. [Google Scholar] [PubMed]
- Mantovani, A. Human and veterinary medicine: The priority for public health synergies. Vet. Ital. 2008, 44, 577–582. [Google Scholar] [PubMed]
- Alves, R.R.N.; Policarpo, I.S. Animals and Human Health: Where Do They Meet? In Ethnozoology: Animals in Our Lives; Alves, R.R.N., Albuquerque, U.P., Eds.; Academic Press: London, UK, 2018; pp. 233–259. [Google Scholar]
- Erkyihun, G.A.; Gari, F.R.; Edao, B.M.; Kassa, G.M. A review on One Health approach in Ethiopia. One Health Outlook 2022, 4, 8. [Google Scholar] [CrossRef]
- Mwangi, D.K. Institutional one health and animal-human health connections in Nthongoni, Eastern Kenya. Health Place 2022, 9, 102818. [Google Scholar] [CrossRef]
- Souto, W.M.S.; Mourão, J.S.; Barboza, R.R.; Alves, R.R.N. Parallels between zootherapeutic practices in ethnoveterinary and human complementary medicine in northeastern Brazil. J. Ethnopharmacol. 2011, 134, 753–767. [Google Scholar] [CrossRef]
- Miara, M.D.; Bendif, H.; Ouabed, A.; Rebbas, K.; Ait Hammou, M.; Amirat, M.; Greene, A.; Teixidor-Toneu, I. Ethnoveterinary remedies used in the Algerian steppe: Exploring the relationship with traditional human herbal medicine. J. Ethnopharmacol. 2019, 244, 112164. [Google Scholar] [CrossRef]
- González, J.A.; Vallejo, J.R. Relics and historical uses of human zootherapeutic products in contemporary Spanish ethnoveterinary medicine. Vet. Sci. 2021, 8, 323. [Google Scholar] [CrossRef]
- Jensen-Jarolim, E. (Ed.) Comparative Medicine: Anatomy and Physiology; Springer: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Lahlou, M. The success of natural products in drug discovery. Pharmacol. Pharm. 2013, 4, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Kang, N.J.; Jin, H.S.; Lee, S.E.; Kim, H.J.; Koh, H.; Lee, D.W. New approaches towards the discovery and evaluation of bioactive peptides from natural resources. Crit. Rev. Environ. Sci. Technol. 2020, 50, 72–103. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T.; The International Natural Product Sciences Taskforce. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Seabrooks, L.; Hu, L. Insects: An underrepresented resource for the discovery of biologically active natural products. Acta Pharm. Sin. B 2017, 7, 409–426. [Google Scholar] [CrossRef] [PubMed]
- Herron, L.R.; Pridans, C.; Turnbull, M.L.; Smith, N.; Lillico, S.; Sherman, A.; Gilhooley, H.J.; Wear, M.; Kurian, D.; Papadakos, G.; et al. A chicken bioreactor for efficient production of functional cytokines. BMC Biotechnol. 2018, 18, 82. [Google Scholar] [CrossRef] [PubMed]
- Summer, K.; Browne, J.; Liu, L.; Benkendorff, K. Molluscan compounds provide drug leads for the treatment and prevention of respiratory disease. Mar. Drugs 2020, 18, 570. [Google Scholar]
- Chonco, L.; Landete-Castillejos, T.; Serrano-Heras, G.; Pérez Serrano, M.; Pérez-Barbería, F.J.; González-Armesto, C.; García, A.; de Cabo, C.; Lorenzo, J.M.; Li, C.; et al. Anti-tumour activity of deer growing antlers and its potential applications in the treatment of malignant gliomas. Sci. Rep. 2021, 11, 42. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.; Sun, M.; Liang, W.; He, M.; Schroën, Y.; Zou, W.; Pommerening, T.; Wang, M. An investigation of the pharmacological applications used for the Ancient Egyptian systemic model ‘ra-ib’ compared with modern Traditional Chinese Medicine. J. Ethnopharmacol. 2021, 265, 113115. [Google Scholar]
- Samy, R.P.; Stiles, B.G.; Franco, O.L.; Sethi, G.; Lim, L.H.K. Animal venoms as antimicrobial agents. Biochem. Pharmacol. 2017, 134, 127–138. [Google Scholar] [CrossRef]
- Chatterjee, B. Animal venoms have potential to treat cancer. Curr. Top. Med. Chem. 2018, 18, 2555–2566. [Google Scholar] [CrossRef]
- Li, F.J.; Hu, J.H.; Ren, X.; Zhou, C.M.; Liu, Q.; Zhang, Y.Q. Toad venom: A comprehensive review of chemical constituents, anticancer activities, and mechanisms. Arch. Pharm. 2021, 354, e2100060. [Google Scholar] [CrossRef]
- Song, M.J.; Kim, H. Analysis of ethnoveterinary treatments for cattle (Bos indicus) diseases referred in Sanrimgyeongje including twelve volumes of literature from the 7th to the 18th century. J. Ethnopharmacol. 2010, 133, 474–479. [Google Scholar] [CrossRef]
- Souto, W.M.S.; Pinto, L.C.; Mendoça, L.E.T.; Mourão, J.S.; Vieira, W.L.S.; Montenegro, P.F.G.P.; Alves, R.R.N. Medicinal animals in ethnoveterinary practices: A world overview. In Animals in Traditional Folk Medicine. Implications for Conservation; Alves, R.R.N., Rosa, I.L., Eds.; Springer: Heidelberg, Germany, 2013; pp. 43–66. [Google Scholar]
- Borges, A.K.M.; Barboza, R.R.D.; Souto, W.M.S.; Alves, R.R.N. Natural Remedies for Animal Health in Latin America. In Ethnoveterinary Medicine: Present and Future Concepts; McGaw, L.J., Abdalla, M.A., Eds.; Springer Nature: Cham, Switzerland, 2020; pp. 311–344. [Google Scholar]
- Vallejo, J.R.; González, J.A. Los anfibios en la medicina popular española, la farmacopea de Plinio y el Dioscórides. Hist. cienc. saude-Manguinhos 2015, 22, 1283–1319. [Google Scholar] [PubMed] [Green Version]
- Thomas, S.C. Differentiation of selected frog (Rana) and toad (Bufo) bones using morphological and osteometric traits. Can. Zooarchaeol. 1996, 10, 2–8. [Google Scholar]
- Hatfield, G. Encyclopedia of Folk Medicine: Old World and New World Traditions; ABC-CLIO: Santa Bárbara, CL, USA, 2004. [Google Scholar]
- Mohneke, M.; Onadeko, A.B.; Rödel, M.O. Medicinal and dietary uses of amphibians in Burkina Faso. Afr. J. Herpetol. 2011, 60, 78–83. [Google Scholar] [CrossRef]
- Rodríguez, C.; Rollins-Smith, L.; Ibáñez, R.; Durant-Archibold, A.A.; Gutiérrez, M. Toxins and pharmacologically active compounds from species of the family Bufonidae (Amphibia, Anura). J. Ethnopharmacol. 2017, 198, 235–254. [Google Scholar] [CrossRef] [Green Version]
- Bell, B. Sistema ó Curso Completo de Cirugía. v. 3; Imp. de los Señores García y Compañía: Madrid, Spain, 1813. [Google Scholar]
- AmphibiaWeb. University of California: Berkeley, CA, USA, 2022. Available online: https://amphibiaweb.org (accessed on 12 June 2022).
- Gómez-Mestre, I. Sapo corredor—Epidalea calamita (Laurenti, 1768). In Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Martínez-Solano, I., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2014; Available online: https://www.vertebradosibericos.org/ (accessed on 12 June 2022).
- Díaz-Paniagua, C.; Gómez-Rodríguez, C.; Portheault, A.; De Vries, W. Los anfibios de Doñana; Organismo Autónomo de Parques Nacionales—Ministerio de Medio Ambiente: Madrid, Spain, 2005.
- Velasco, J.C.; Lizana, M.; Sancho, J.R.; Delibes de Castro, M.; Fernández Gutiérrez, J. Guía de Peces, Anfibios, Reptiles Y Mamíferos de Castilla y León; Náyade Editorial: Medina del Campo, Spain; Valladolid, Spain, 2005. [Google Scholar]
- Martínez-Solano, I. Atlas de distribución y estado de conservación de los anfibios de la Comunidad de Madrid. Graellsia 2006, 62, 253–291. [Google Scholar] [CrossRef] [Green Version]
- González-Miras, E.; Nevado, J.C. Atlas de distribución de los anfibios de la provincia de Almería (sudeste ibérico, España). Bol. Asoc. Herpetol. Esp. 2008, 19, 85–90. [Google Scholar]
- Diego-Rasilla, F.J.; Ortiz-Santaliestra, M.E. Naturaleza en Castilla y León: Los Anfibios; Caja de Burgos: Burgos, Spain, 2009. [Google Scholar]
- Reyne, M.; Dicks, K.; McFarlane, C.; Aubry, A.; Emmerson, M.; Marnell, F.; Reid, N.; Helyar, S. Population genetic structure of the Natterjack toad (Epidalea calamita) in Ireland: Implications for conservation management. Conserv. Genet. 2021, 23, 325–339. [Google Scholar] [CrossRef]
- Beja, P.; Kuzmin, S.; Beebee, T.; Denoel, M.; Schmidt, B.; Tarkhnishvili, D.; Ananjeva, N.B.; Orlov, N.L.; Nyström, P.; Ogrodowczyk, A.; et al. Epidalea Calamita (Errata Version Published in 2016). The IUCN Red List of Threatened Species 2009: E.T54598A86640094. Available online: https://doi.org/10.2305/IUCN.UK.2009.RLTS.T54598A11160828.en (accessed on 12 June 2022).
- González, J.A.; Amich, F.; Postigo-Mota, S.; Vallejo, J.R. The use of wild vertebrates in contemporary Spanish ethnoveterinary medicine. J. Ethnopharmacol. 2016, 191, 135–151. [Google Scholar] [CrossRef]
- Vallejo, J.R.; González, J.A. Fish-based remedies in Spanish ethnomedicine: A review from a historical perspective. J. Ethnobiol. Ethnomed. 2014, 10, 37. [Google Scholar] [CrossRef] [Green Version]
- González, J.A.; Amich, F.; Postigo-Mota, S.; Vallejo, J.R. Therapeutic and prophylactic uses of invertebrates in contemporary Spanish ethnoveterinary medicine. J. Ethnobiol. Ethnomed. 2016, 12, 36. [Google Scholar] [CrossRef] [Green Version]
- González, J.A.; Vallejo, J.R. The use of domestic animals and their derivative products in contemporary Spanish ethnoveterinary medicine. J. Ethnopharmacol. 2021, 271, 113900. [Google Scholar] [CrossRef] [PubMed]
- Ceríaco, L.M.P. A review of fauna used in zootherapeutic remedies in Portugal: Historical origins, current uses, and implications for conservation. In Animals in Traditional Folk Medicine. Implications for Conservation; Alves, R.R.N., Rosa, I.L., Eds.; Springer: Heidelberg, Germany, 2013; pp. 317–345. [Google Scholar]
- Charro Gorgojo, M.A. Sapos: Historia de una maldición. Rev. Folk. 2000, 235, 20–32. [Google Scholar]
- González, J.A.; Vallejo, J.R.; Aparicio, A.J. Etnozoología: Recursos Animales Para la Salud en la Tradición Salmantina; Diputación de Salamanca: Salamanca, Spain, 2015. [Google Scholar]
- Penco, A.D. Medicina Popular Veterinaria en la Comarca de Zafra. Ph.D. Thesis, Facultad de Medicina, Universidad de Extremadura, Badajoz, Spain, 2005. [Google Scholar]
- Garmendia Larrañaga, J. Rito y Fórmula en la Medicina Popular Vasca: La Salud Por las Plantas Medicinales; Txertoa: San Sebastián, Spain, 1990. [Google Scholar]
- Vázquez Gallego, X. Tradiciones, Mitos, Creencias y Curanderismo en Medicina Popular de Galicia; Diputación Provincial de Lugo: Lugo, Spain, 1990. [Google Scholar]
- Sánchez Pérez, J.A. Supersticiones Españolas; Saeta: Madrid, Spain, 1948. [Google Scholar]
- Barandiaran, J.; Manterola, A. Medicina Popular en Vasconia; Atlas Etnográfico de Vasconia volume V; Etniker Euskalerria, Eusko Jaurlaritza y Gobierno de Navarra: Bilbao, Spain, 2004. [Google Scholar]
- Flores Arroyuelo, F.J. Diccionario de Supersticiones y Creencias Populares, 2nd ed.; Alianza: Madrid, Spain, 2005. [Google Scholar]
- García Ramos, J.A. La Medicina Popular en Almería. Ensayo de Antropología Cultural; Author’s Edition: Albox, Spain, 2010. [Google Scholar]
- Crnobrnja-Isailović, J.; Jovanović, B.; Čubrić, T.; Ćorović, J.; Gopčević, K. The benefits of active substances in amphibians and reptiles and the jeopardy of losing those species forever. In Biodiversity and Biomedicine: Our Future; Ozturk, M., Egamberdieva, D., Pešić, M., Eds.; Academic Press: London, UK, 2020; pp. 107–122. [Google Scholar]
- Kiss, G.; Michl, H. On the venomous skin secretion of the orange-speckled frog Bombina Variegata. Toxicon 1962, 1, 33–39. [Google Scholar] [CrossRef]
- Spande, T.F.; Garraffo, H.M.; Edwards, M.W.; Yeh, H.J.C.; Pannell, L.; Daly, J.W. Epibatidine: A novel (chloropyridyl) azabicycloheptane with potent analgesic activity from an Ecuadoran poison frog. J. Am. Chem. Soc. 1992, 114, 3475–3478. [Google Scholar] [CrossRef]
- Wilson, E.O.; Alfred, A. The Future of Life; Alfred A. Knopf.: New York, NY, USA, 2002. [Google Scholar]
- Alves, R.R.N.; Rosa, I.M.L. Biodiversity, traditional medicine and public health: Where do they meet? J. Ethnobiol. Ethnomed. 2007, 3, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampath, V.P.; Horesh, N.; Sasi, B.; Zannadeh, H.; Pogodin, I.; Singh, S.V.; Deutsch, J.; Lichtstein, D. Synthesis and biological evaluation of novel bufalin derivatives. Int. J. Mol. Sci. 2022, 23, 4007. [Google Scholar] [CrossRef] [PubMed]
- Sõukand, R.; Hrynevich, Y.; Prakofjewa, J.; Valodzina, T.; Vasilyeva, I.; Paciupa, J.; Shrubok, A.; Hlushko, A.; Knureva, Y.; Litvinava, Y.; et al. Use of cultivated plants and non-plant remedies for human and animal home-medication in Liubań district, Belarus. J. Ethnobiol. Ethnomed. 2017, 13, 54. [Google Scholar] [CrossRef] [Green Version]
- Bullitta, S.; Re, G.A.; Manunta, M.D.I.; Piluzza, G. Traditional knowledge about plant, animal, and mineral-based remedies to treat cattle, pigs, horses, and other domestic animals in the Mediterranean island of Sardinia. J. Ethnobiol. Ethnomed. 2018, 14, 50. [Google Scholar] [CrossRef] [Green Version]
- Quave, C.L.; Lohani, U.; Verde, A.; Fajardo, J.; Rivera, D.; Obón, C.; Valdés, A.; Pieroni, A. A comparative assessment of zootherapeutic remedies from selected areas in Albania, Italy, Spain and Nepal. J. Ethnobiol. 2010, 30, 92–125. [Google Scholar] [CrossRef] [Green Version]
- Alves, R.R.N.; Vieira, W.L.S.; Santana, G.G.; Vieira, K.S.; Montenegro, P.F.G.P. Herpetofauna used in traditional folk medicine: Conservation implications. In Animals in Traditional Folk Medicine. Implications for Conservation; Alves, R.R.N., Rosa, I.L., Eds.; Springer: Heidelberg, Germany, 2013; pp. 109–133. [Google Scholar]
- Qi, J.; Tan, C.K.; Hashimi, S.M.; Zulfiker, A.H.M.; Good, D.; Wei, M.Q. Toad glandular secretions and skin extractions as anti-inflammatory and anticancer agents. Evid.-Based Complement. Altern. Med. 2014, 2014, 312684. [Google Scholar] [CrossRef] [Green Version]
- Zhan, X.; Wu, H.; Wu, H.; Wang, R.; Luo, C.; Gao, B.; Chen, Z.; Li, Q. Metabolites from Bufo gargarizans (Cantor, 1842): A review of traditional uses, pharmacological activity, toxicity and quality control. J. Ethnopharmacol. 2020, 246, 112178. [Google Scholar] [CrossRef]
- Chen, K.K.; Kovaríková, A. Pharmacology and toxicology of toad venom. J. Pharm. Sci. 1967, 56, 1535–1541. [Google Scholar] [CrossRef]
- Yoshida, S.; Kamano, Y.; Sakai, T. Studies on the surface anesthetic activity of bufadienolides isolated from ch’ an su. Chem. Pharm. Bull. 1976, 24, 1714–1717. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.D.; Narui, T.; Takatsuki, S.; Hashimoto, T.; Kobayashi, F.; Ekimoto, H.; Abuki, H.; Niijima, K.; Okuyama, T. Hematological studies on naturally occurring substances. VI. Effects of an animal crude drug “chan su” (bufonis venenum) on blood coagulation, platelet aggregation, fibrinolysis system and cytotoxicity. Chem. Pharm. Bull. 1991, 39, 2135–2137. [Google Scholar] [CrossRef] [Green Version]
- Wei, W.L.; Hou, J.J.; Wang, X.; Yu, Y.; Li, H.J.; Li, Z.W.; Feng, Z.J.; Qu, H.; Wu, W.Y.; Guo, D.A. Venenum bufonis: An overview of its traditional use, natural product chemistry, pharmacology, pharmacokinetics and toxicology. J. Ethnopharmacol. 2019, 237, 215–235. [Google Scholar] [CrossRef]
- Xu, R.; Xie, H.Q.; Deng, L.L.; Zhang, J.X.; Yang, F.M.; Liu, J.H.; Hao, X.J.; Zhang, Y.H. A new bufadienolide with cytotoxic activity from the Chinese traditional drug Ch’an Su. Chin. J. Nat. Med. 2014, 12, 623–627. [Google Scholar] [CrossRef]
- García París, M. Los Anfibios de España; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 1985.
- Gómez-Mestre, I.; Tejedo, M. Contrasting patterns of quantitative and neutral genetic variation in locally adapted populations of the natterjack toad, Bufo calamita. Evolution 2004, 58, 2343–2352. [Google Scholar] [CrossRef] [Green Version]
- Heus, F.; Otvos, R.A.; Aspers, R.L.; van Elk, R.; Halff, J.I.; Ehlers, A.W.; Dutertre, S.; Lewis, R.J.; Wijmenga, S.; Smit, A.B.; et al. Miniaturized bioaffinity assessment coupled to mass spectrometry for guided purification of bioactives from toad and cone snail. Biology 2014, 3, 139–156. [Google Scholar] [CrossRef] [Green Version]
- Santos, X.; Llorente, G.A.; Montori, A.; Carretero, M.A.; Franch, M.; Garriga, N.; Richter-Boix, A. Evaluating factors affecting amphibian mortality on roads: The case of the common toad Bufo bufo, near a breeding place. Anim. Biodivers. Conserv. 2007, 30, 97–104. [Google Scholar]
- Sillero, N. Amphibian mortality levels on Spanish country roads: Descriptive and spatial analysis. Amphib. -Reptil. 2008, 29, 337–347. [Google Scholar] [CrossRef]
- Toledo, R.C.; Jared, C.; Brunner Junior, A. Morphology of the large granular alveoli of the parotoid glands in toad (Bufo ictericus) before and after compression. Toxicon 1992, 30, 745–753. [Google Scholar] [CrossRef]
- Roelants, K.; Fry, B.G.; Ye, L.; Stijlemans, B.; Brys, L.; Kok, P.; Clynen, E.; Schoofs, L.; Cornelis, P.; Bossuyt, F. Origin and functional diversification of an amphibian defense peptide arsenal. PLoS Genet. 2013, 9, e1003662. [Google Scholar] [CrossRef] [Green Version]
- Daly, J.W. Ernest Guenther Award in Chemistry of Natural Products. Amphibian Skin: A Remarkable Source of Biologically Active Arthropod Alkaloids. J. Med. Chem. 2003, 46, 445–452. [Google Scholar] [CrossRef]
- Mitchell-Olds, T.; Willis, J.H.; Goldstein, D.B. Which evolutionary processes influence natural genetic variation for phenotypic traits? Nat. Rev. Genet. 2007, 8, 845–856. [Google Scholar] [CrossRef]
- Hallgrímmson, B.; Brown, J.J.Y.; Hall, B.K. The Study of Phenotypic Variability: An Emerging Research Agenda for Understanding the Developmental—Genetic Architecture Underlying Phenotypic Variation. In Variation: A Central Concept in Biology; Hallgrímsson, B., Hall, B.K., Eds.; Academic Press: London, UK, 2005; pp. 525–551. [Google Scholar]
- König, E.; Bininda-Emonds, O.R.P.; Shaw, C. The diversity and evolution of anuran skin peptides. Peptides 2015, 63, 96–117. [Google Scholar] [CrossRef]
- Jørgensen, C.B. 200 years of amphibian water economy: From Robert Townson to the present. Biol. Rev. Camb. Philos. Soc. 1997, 72, 153–237. [Google Scholar] [CrossRef]
- Delfino, G.; Brizzi, R.; Alvarez, B.B.; Taddei, L. Secretory polymorphism and serous cutaneous gland heterogeneity in Bufo granulosus (Amphibia, Anura). Toxicon 1999, 37, 1281–1296. [Google Scholar] [CrossRef]
- Delfino, G.; Nosi, D.; Giachi, F. Secretory granule-cytoplasm relationships in serous glands of anurans: Ultrastructural evidence and possible functional role. Toxicon 2001, 39, 1161–1171. [Google Scholar] [CrossRef]
- Bennett, D. The Pigmentary System. Pigment Cell Res. 2006, 19, 469. [Google Scholar] [CrossRef]
- Rollins-Smith, L.A.; Reinert, L.K.; O’Leary, C.J.; Houston, L.E.; Woodhams, D.C. Antimicrobial peptide defenses in amphibian skin. Integr. Comp. Biol. 2005, 45, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Rollins-Smith, L.A. The role of amphibian antimicrobial peptides in protection of amphibians from pathogens linked to global amphibian declines. Biochim. Biophys. Acta 2009, 1788, 1593–1599. [Google Scholar] [CrossRef] [Green Version]
- Hostetler, J.R.; Cannon, M.S. The anatomy of the parotoid gland in Bufonidae with some histochemical findings. I. Bufo marinus. J. Morphol. 1974, 142, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Cannon, M.S.; Hostetler, J.R. The anatomy of the parotoid gland in Bufonidae with some histochemical findings II. Bufo alvarius. J. Morphol. 1976, 148, 137–160. [Google Scholar] [CrossRef] [PubMed]
- Mailho-Fontana, P.L.; Antoniazzi, M.M.; Sciani, J.M.; Pimenta, D.C.; Barbaro, K.C.; Jared, C. Morphological and biochemical characterization of the cutaneous poison glands in toads (Rhinella marina group) from different environments. Front. Zool. 2018, 15, 46. [Google Scholar] [CrossRef] [Green Version]
- Nelsen, D.R.; Nisani, Z.; Cooper, A.M.; Fox, G.A.; Gren, E.C.K.; Corbit, A.G.; Hayes, W.K. Poisons, toxungens, and venoms: Redefining and classifying toxic biological secretions and the organisms that employ them. Biol. Rev. Camb. Philos. Soc. 2014, 89, 450–465. [Google Scholar] [CrossRef]
- Toledo, R.C.; Jared, C. Cutaneous granular glands and amphibian venoms. Comp. Biochem. Physiol. Part A Physiol. 1995, 111, 1–29. [Google Scholar] [CrossRef]
- Bucciarelli, G.M.; Shaffer, H.B.; Green, D.B.; Kats, L.B. An amphibian chemical defense phenotype is inducible across life history stages. Sci. Rep. 2017, 7, 8185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nalbantsoy, A.; Kariş, M.; Yalcin, H.T.; Göçmen, B. Biological activities of skin and parotoid gland secretions of bufonid toads (Bufo bufo, Bufo verrucosissimus and Bufotes variabilis) from Turkey. Biomed. Pharmacother. 2016, 80, 298–303. [Google Scholar] [CrossRef]
- Demori, I.; El Rashed, Z.; Corradino, V.; Catalano, A.; Rovegno, L.; Queirolo, L.; Salvidio, S.; Biggi, E.; Zanotti-Russo, M.; Canesi, L.; et al. Peptides for skin protection and healing in amphibians. Molecules 2019, 24, 347. [Google Scholar] [CrossRef] [Green Version]
- Ribatti, D.; Santoiemma, M. Epithelial-mesenchymal interactions: A fundamental developmental biology mechanism. Int. J. Dev. Biol. 2014, 58, 303–306. [Google Scholar] [CrossRef] [Green Version]
- Willmore, K.E.; Hallgrímsson, B. Within Individual Variation: Developmental Noise versus Developmental Stability. In Variation: A Central Concept in Biology; Hallgrímsson, B., Hall, B.K., Eds.; Academic Press: London, UK, 2005; pp. 191–218. [Google Scholar]
- McKinney, M.L.; McNamara, K.J. (Eds.) Heterochrony: The Evolution of Ontogeny; Springer: Boston, MA, USA, 1991. [Google Scholar]
- McNamara, K.J. Heterochrony: The Evolution of Development. Evo. Edu. Outreach 2012, 5, 203–218. [Google Scholar] [CrossRef] [Green Version]
- Quagliata, S.; Malentacchi, C.; Delfino, C.; Brunasso, A.M.G.; Delfino, G. Adaptive evolution of secretory cell lines in vertebrate skin. Caryologia 2006, 59, 187–206. [Google Scholar] [CrossRef] [Green Version]
- Pigliucci, M. Phenotypic Plasticity: Beyond Nature and Nurture; Johns Hopkins University Press: Baltimore, MD, USA, 2001. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diseases or Troubles Treated | Geographical Location | References | |
|---|---|---|---|
| EVM | Scald, hoof rot | Unspecified | Charro Gorgojo [50] |
| Wounds | Fuenteguinaldo (Salamanca) | González et al. [51] | |
| Skin ulcerations | Comarca de Zafra (Badajoz) | Penco [52] | |
| ETHNOMED | Anthrax | Basque Country | Garmendia Larrañaga [53] |
| Brucellosis | Valencia | Charro Gorgojo [50] | |
| Tonsillitis | Lalín (Pontevedra) | Sánchez Pérez [54] | |
| Galicia | Vázquez Gallego [55] | ||
| Headache | Basque Country | Barandiaran & Manterola [56] | |
| Warts | Galicia | Charro Gorgojo [50], Flores Arroyuelo [57] | |
| Herpes zoster | Albox (Almería) | García Ramos [58] | |
| Venomous animal bites (spiders, snakes) | Unspecified | Flores Arroyuelo [57] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vallejo, J.R.; González, J.A.; Gómez-Navarro, M.E.; López-Cepero, J.M. Histological Study of Glandular Variability in the Skin of the Natterjack Toad—Epidalea calamita (Laurenti, 1768)—Used in Spanish Historical Ethnoveterinary Medicine and Ethnomedicine. Vet. Sci. 2022, 9, 423. https://doi.org/10.3390/vetsci9080423
Vallejo JR, González JA, Gómez-Navarro ME, López-Cepero JM. Histological Study of Glandular Variability in the Skin of the Natterjack Toad—Epidalea calamita (Laurenti, 1768)—Used in Spanish Historical Ethnoveterinary Medicine and Ethnomedicine. Veterinary Sciences. 2022; 9(8):423. https://doi.org/10.3390/vetsci9080423
Chicago/Turabian StyleVallejo, José Ramón, José A. González, María Eugenia Gómez-Navarro, and José María López-Cepero. 2022. "Histological Study of Glandular Variability in the Skin of the Natterjack Toad—Epidalea calamita (Laurenti, 1768)—Used in Spanish Historical Ethnoveterinary Medicine and Ethnomedicine" Veterinary Sciences 9, no. 8: 423. https://doi.org/10.3390/vetsci9080423
APA StyleVallejo, J. R., González, J. A., Gómez-Navarro, M. E., & López-Cepero, J. M. (2022). Histological Study of Glandular Variability in the Skin of the Natterjack Toad—Epidalea calamita (Laurenti, 1768)—Used in Spanish Historical Ethnoveterinary Medicine and Ethnomedicine. Veterinary Sciences, 9(8), 423. https://doi.org/10.3390/vetsci9080423