
Developing a Novel Immune-Related Seven-Gene Signature and Immune Infiltration Pattern in Patients with COVID-19 and Cardiovascular Disease

Abstract
:1. Introduction
2. Materials and Methods
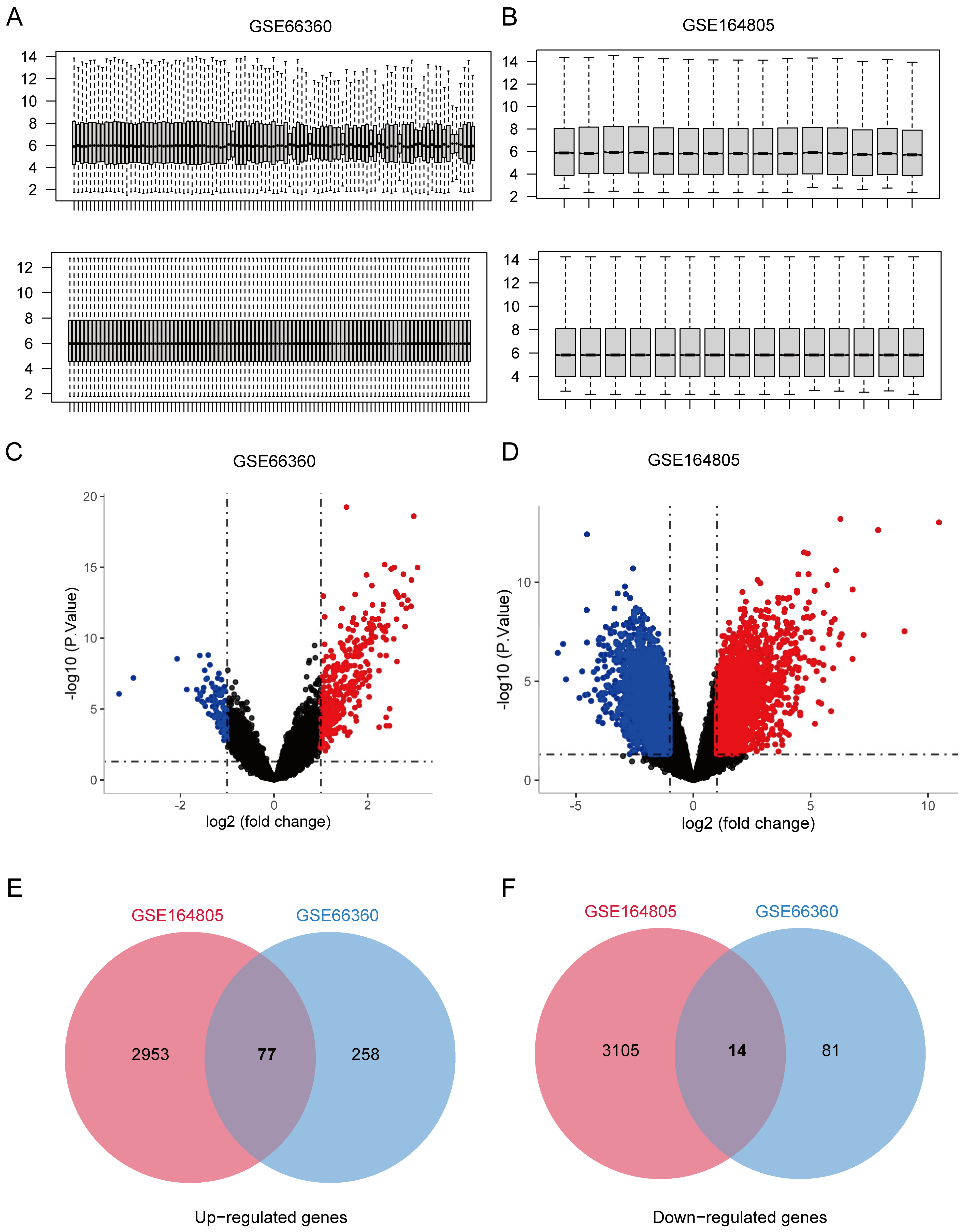
2.1. Microarray Datasets Collection and Preprocessing
2.2. KEGG and GO Enrichment Analyses
2.3. Establishment of Protein–Protein Interaction (PPI) Networks
2.4. Candidate Small Molecular Agents (SMAs) Prediction
2.5. Immune Cell Infiltration Evaluation and Its Correlation with Hub IRGs (HIRGs)
2.6. Diagnostic Efficacy Evaluation of HIRGs and Their Expressional Correlation
2.7. Gene Set Enrichment Analysis (GSEA)-Based Pathway Confirmation Study
3. Results
3.1. Co-Expression Genes Shared in CVD and COVID-19
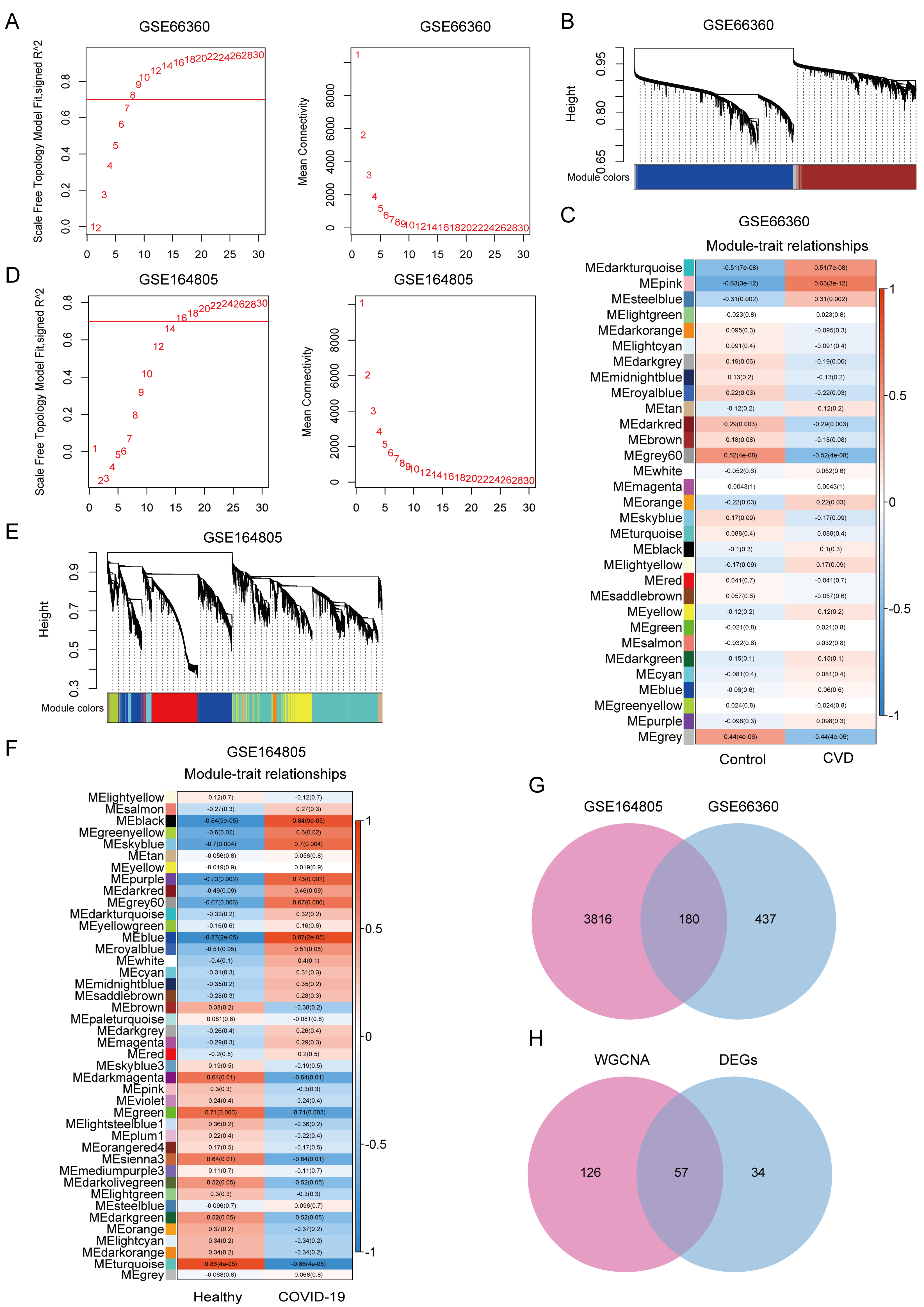
3.2. WGCNA Reveals Co-Expression Modules Associated with CVD and COVID-19
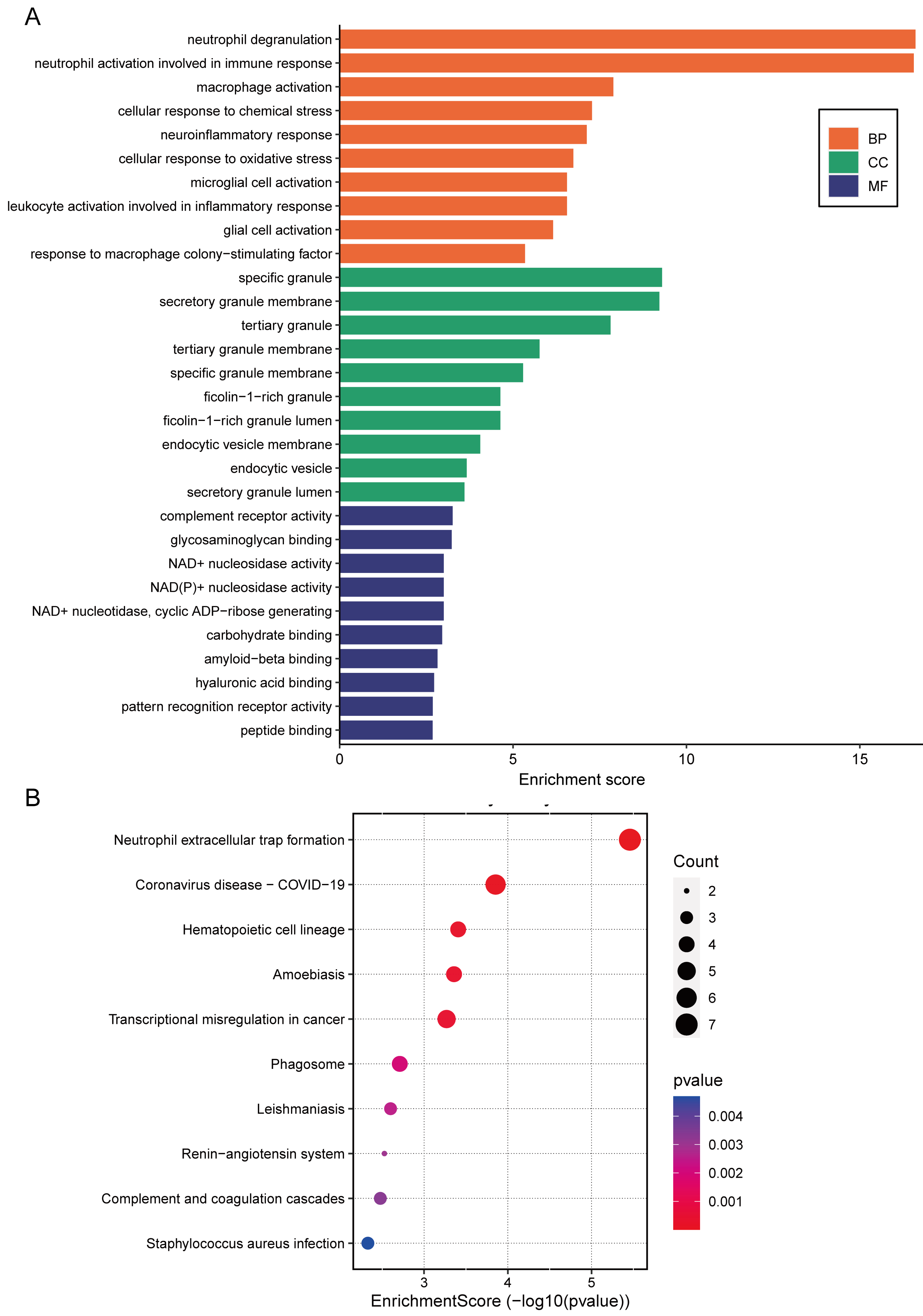
3.3. Functional Enrichment of the CGS
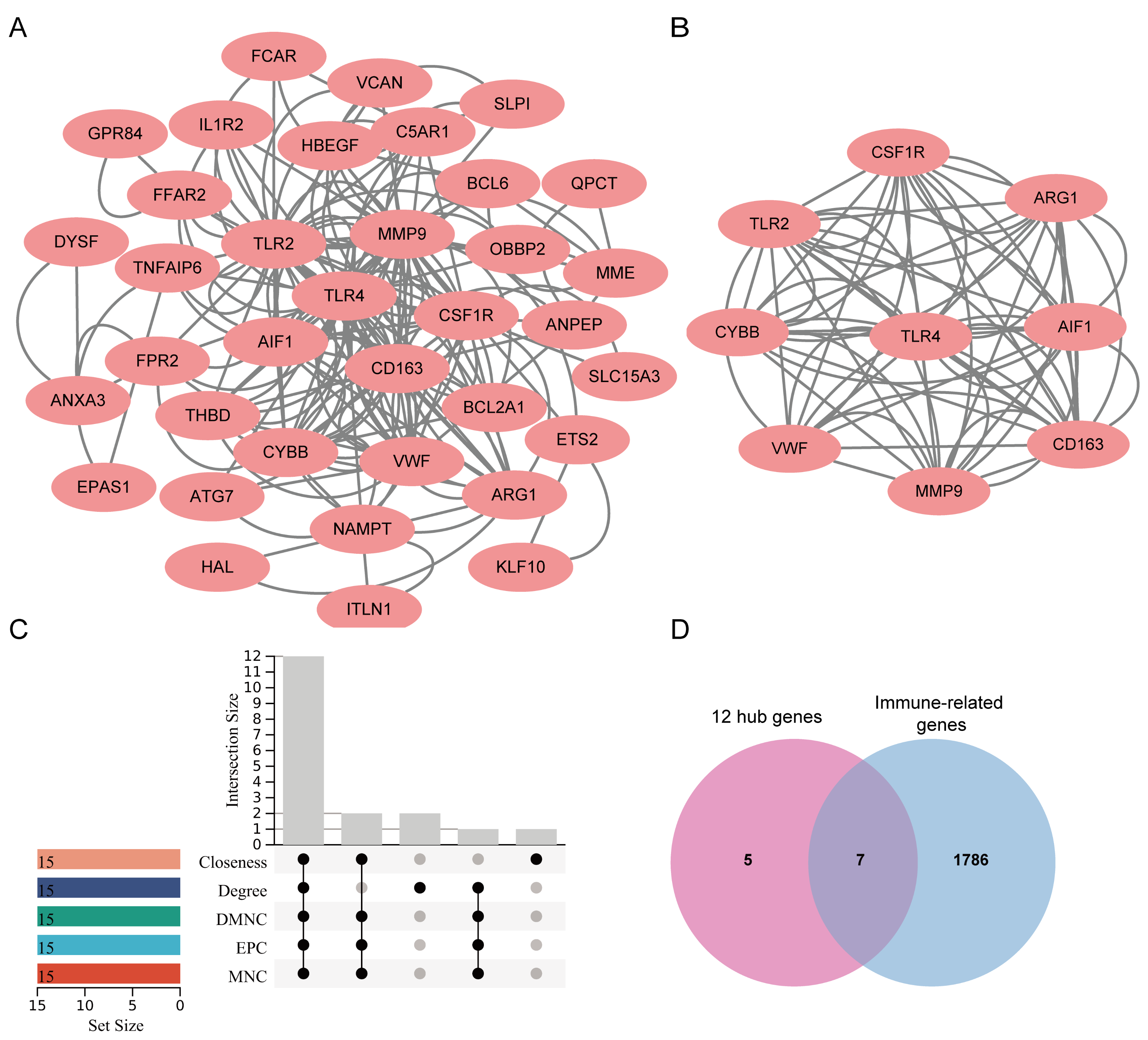
3.4. Identification and Modular Analysis of Hub Genes
3.5. Identification of Candidate Drugs
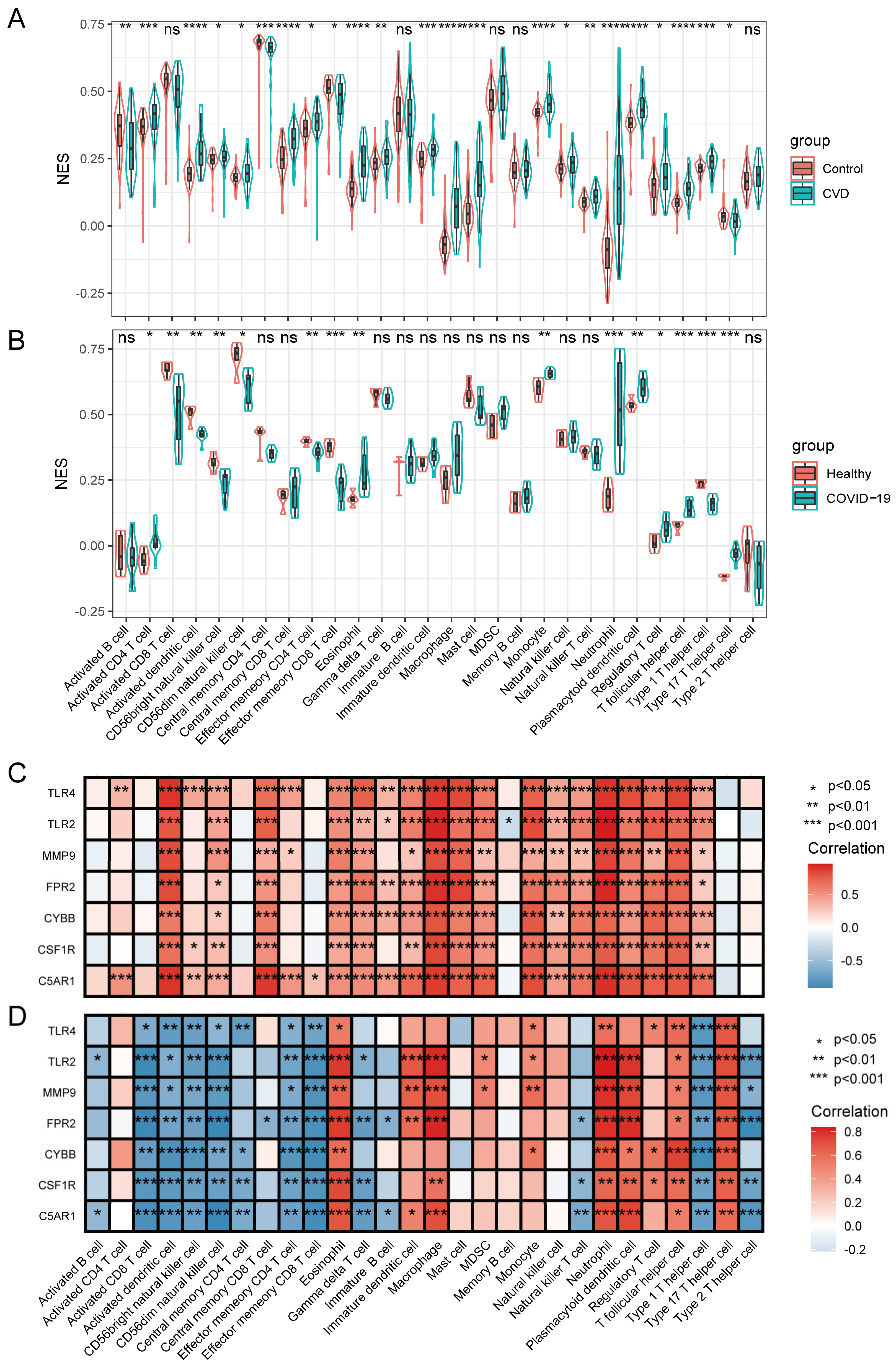
3.6. Immune Infiltrating Cell Analyses and Their Relationship with HIRGs
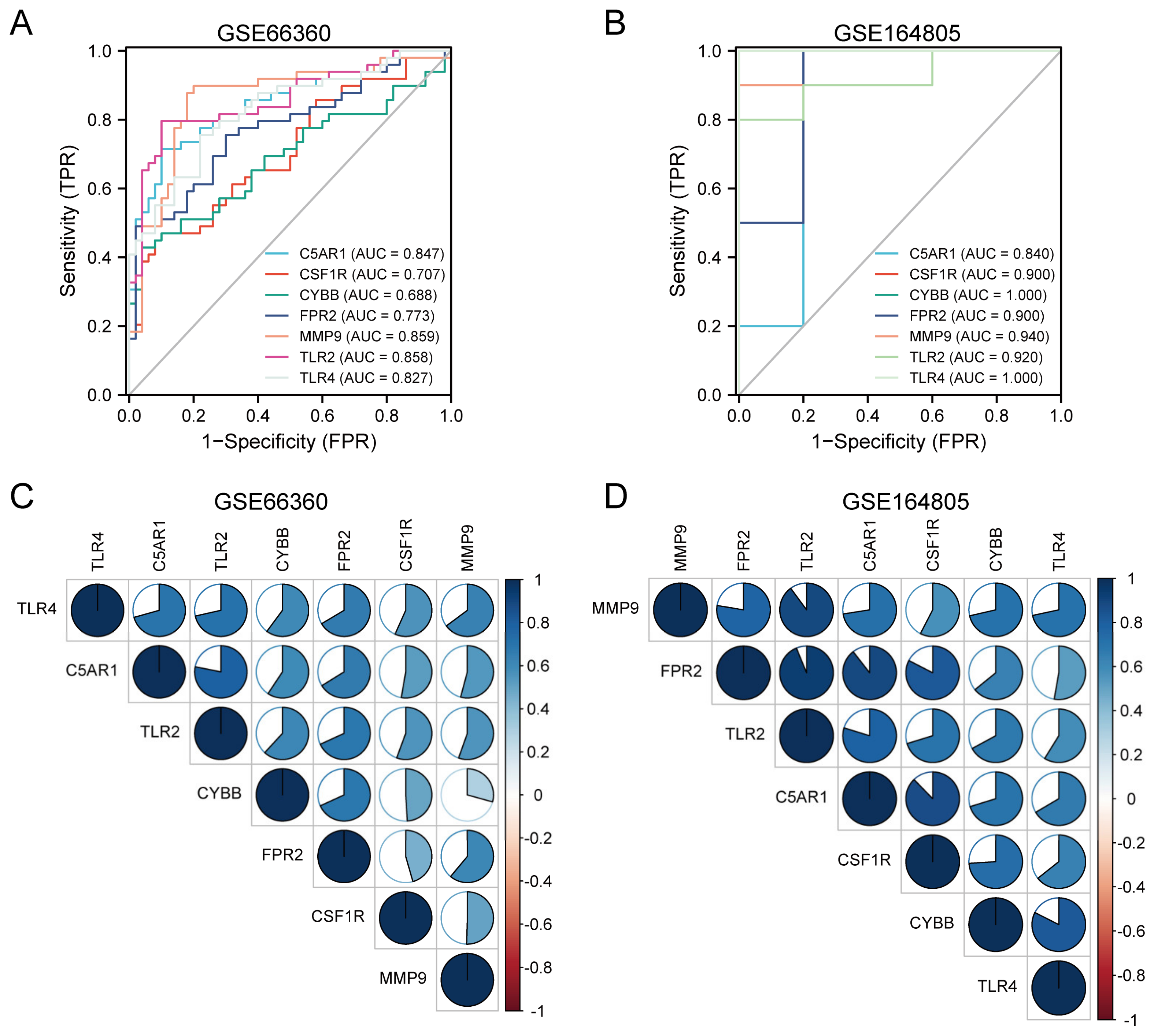
3.7. Diagnostic Performance and Correlation Analysis of the HIRGs
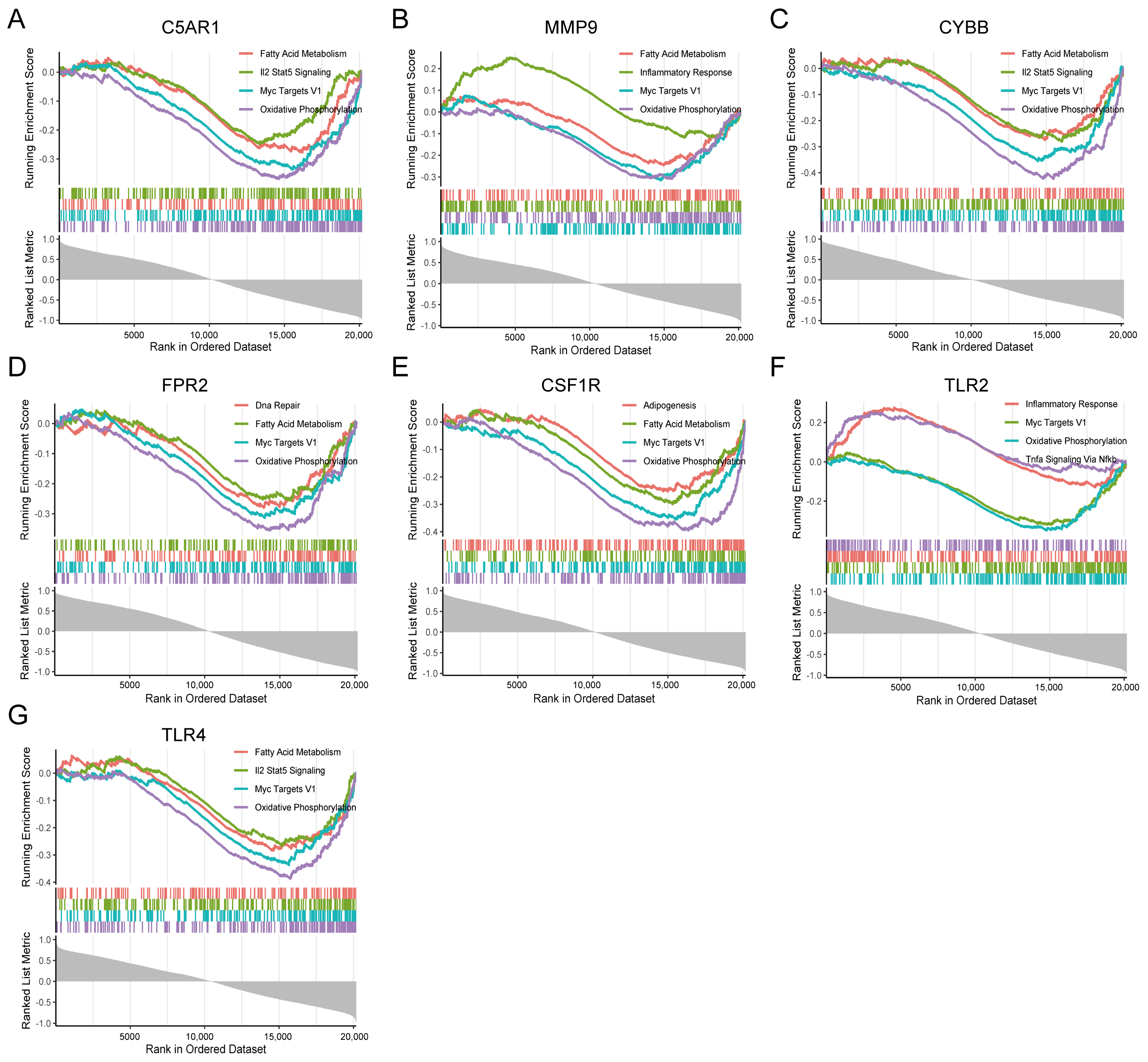
3.8. GSEA Identifies Seven HIRGs Associated Signaling Pathway
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z.; Benjamin, E.J.; Benziger, C.P.; et al. Global Burden of Cardiovascular Diseases and Risk Factors, 1990-2019: Update From the GBD 2019 Study. J. Am. Coll. Cardiol. 2020, 76, 2982–3021. [Google Scholar] [CrossRef]
- WHO. The Top 10 Causes of Death. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 10 October 2022).
- Omran, F.; Kyrou, I.; Osman, F.; Lim, V.G.; Randeva, H.S.; Chatha, K. Cardiovascular Biomarkers: Lessons of the Past and Prospects for the Future. Int. J. Mol. Sci. 2022, 23, 5680. [Google Scholar] [CrossRef]
- Abdar, M.; Salari, S.; Qahremani, S.; Lam, H.K.; Karray, F.; Hussain, S.; Khosravi, A.; Acharya, U.R.; Makarenkov, V.; Nahavandi, S. UncertaintyFuseNet: Robust uncertainty-aware Hierarchical Feature Fusion Model with Ensemble Monte Carlo Dropout for COVID-19 Detection. Inf. Fusion 2022, 90, 364–381. [Google Scholar] [CrossRef]
- Lenihan, D.; Carver, J.; Porter, C.; Liu, J.E.; Dent, S.; Thavendiranathan, P.; Mitchell, J.D.; Nohria, A.; Fradley, M.G.; Pusic, I.; et al. Cardio-oncology Care in the Era of the Coronavirus Disease 2019 (COVID-19) Pandemic: An International Cardio-Oncology Society (ICOS) Statement. CA Cancer J. Clin. 2020, 70, 480–504. [Google Scholar] [CrossRef]
- Modin, D.; Claggett, B.; Sindet-Pedersen, C.; Lassen, M.C.H.; Skaarup, K.G.; Jensen, J.U.S.; Fralick, M.; Schou, M.; Lamberts, M.; Gerds, T.; et al. Acute COVID-19 and the Incidence of Ischemic Stroke and Acute Myocardial Infarction. Circulation 2020, 142, 2080–2082. [Google Scholar] [CrossRef] [PubMed]
- Katsoularis, I.; Fonseca-Rodriguez, O.; Farrington, P.; Lindmark, K.; Fors Connolly, A.M. Risk of Acute Myocardial Infarction and Ischaemic Stroke Following COVID-19 in Sweden: A self-controlled Case Series and Matched Cohort Study. Lancet 2021, 398, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Knight, R.; Walker, V.; Ip, S.; Cooper, J.A.; Bolton, T.; Keene, S.; Denholm, R.; Akbari, A.; Abbasizanjani, H.; Torabi, F.; et al. Association of COVID-19 With Major Arterial and Venous Thrombotic Diseases: A Population-Wide Cohort Study of 48 Million Adults in England and Wales. Circulation 2022, 146, 892–906. [Google Scholar] [CrossRef] [PubMed]
- Kyriakoulis, K.G.; Dimakakos, E.; Kyriakoulis, I.G.; Catalano, M.; Spyropoulos, A.C.; Schulman, S.; Douketis, J.; Falanga, A.; Maraveyas, A.; Olinic, D.M.; et al. Practical Recommendations for Optimal Thromboprophylaxis in Patients with COVID-19: A Consensus Statement Based on Available Clinical Trials. J. Clin. Med. 2022, 11, 5997. [Google Scholar] [CrossRef]
- Dimakakos, E.; Gomatou, G.; Catalano, M.; Olinic, D.M.; Spyropoulos, A.C.; Falanga, A.; Maraveyas, A.; Liew, A.; Schulman, S.; Belch, J.; et al. Thromboembolic Disease in Patients With Cancer and COVID-19: Risk Factors, Prevention and Practical Thromboprophylaxis Recommendations-State-of-the-Art. Anticancer Res. 2022, 42, 3261–3274. [Google Scholar] [CrossRef]
- Hamming, I.; Timens, W.; Bulthuis, M.L.; Lely, A.T.; Navis, G.; van Goor, H. Tissue distribution of ACE2 protein, the Functional Receptor for SARS Coronavirus. A First Step in Understanding SARS Pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Woodruff, T.M.; Shukla, A.K. The Complement C5a-C5aR1 GPCR Axis in COVID-19 Therapeutics. Trends Immunol. 2020, 41, 965–967. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The Role of Pattern-Recognition Receptors in Innate Immunity: Update on Toll-like Receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Muse, E.D.; Kramer, E.R.; Wang, H.; Barrett, P.; Parviz, F.; Novotny, M.A.; Lasken, R.S.; Jatkoe, T.A.; Oliveira, G.; Peng, H.; et al. A Whole Blood Molecular Signature for Acute Myocardial Infarction. Sci. Rep. 2017, 7, 12268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Meng, Y.; Wang, K.; Zhang, X.; Chen, W.; Sheng, J.; Qiu, Y.; Diao, H.; Li, L. Inflammation and Antiviral Immune Response Associated with Severe Progression of COVID-19. Front. Immunol. 2021, 12, 631226. [Google Scholar] [CrossRef] [PubMed]
- Irizarry, R.A.; Hobbs, B.; Collin, F.; Beazer-Barclay, Y.D.; Antonellis, K.J.; Scherf, U.; Speed, T.P. Exploration, Normalization, and Summaries of High Density Oligonucleotide Array Probe Level Data. Biostatistics 2003, 4, 249–264. [Google Scholar] [CrossRef] [Green Version]
- Reimers, M.; Carey, V.J. Bioconductor: An Open Source Framework for Bioinformatics and Computational Biology. Methods Enzymol. 2006, 411, 119–134. [Google Scholar] [CrossRef]
- Ito, K.; Murphy, D. Application of ggplot2 to Pharmacometric Graphics. CPT Pharmacometrics Syst. Pharmacol. 2013, 2, e79. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R Package for Weighted Correlation Network Analysis. BMC Bioinformatics 2008, 9, 559. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING Database in 2021: Customizable Protein-Protein Networks, and Functional Characterization of User-Uploaded Gene/Measurement Sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Liu, X.; Lou, L.; Zhou, L. Molecular Mechanisms of Cardiac Injury Associated with Myocardial SARS-CoV-2 Infection. Front Cardiovasc. Med. 2021, 8, 643958. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Chin, C.H.; Chen, S.H.; Wu, H.H.; Ho, C.W.; Ko, M.T.; Lin, C.Y. Cytohubba: Identifying Hub Objects and Sub-Networks from Complex Interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, A.; Narayan, R.; Corsello, S.M.; Peck, D.D.; Natoli, T.E.; Lu, X.; Gould, J.; Davis, J.F.; Tubelli, A.A.; Asiedu, J.K.; et al. A Next Generation Connectivity Map: L1000 Platform and the First 1,000,000 Profiles. Cell 2017, 171, 1437–1452.e17. [Google Scholar] [CrossRef] [PubMed]
- Charoentong, P.; Finotello, F.; Angelova, M.; Mayer, C.; Efremova, M.; Rieder, D.; Hackl, H.; Trajanoski, Z. Pan-cancer Immunogenomic Analyses Reveal Genotype-Immunophenotype Relationships and Predictors of Response to Checkpoint Blockade. Cell Rep. 2017, 18, 248–262. [Google Scholar] [CrossRef] [Green Version]
- Hanzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene Set Variation Analysis for Microarray and RNA-seq Data. BMC Bioinformatics 2013, 14, 7. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.; Simko, V.; Levy, M.; Xie, Y.; Jin, Y.; Zemla, J. R Package ‘corrplot’: Visualization of a Correlation Matrix (Version 0.92). 2021. Available online: https://github.com/taiyun/corrplot (accessed on 10 October 2022).
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Kotlyar, M.; Pastrello, C.; Ahmed, Z.; Chee, J.; Varyova, Z.; Jurisica, I. IID 2021: Towards Context-Specific Protein Interaction Analyses by Increased Coverage, Enhanced Annotation and Enrichment Analysis. Nucleic Acids Res. 2022, 50, D640–D647. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Kuo, Y.H.; Zhong, M.; Zhang, J.; Zhou, X.X.; Xing, L.; Wells, J.A.; Wang, Y.; Arkin, M.R. Adaptor-Specific Antibody Fragment Inhibitors for the Intracellular Modulation of p97 (VCP) Protein-Protein Interactions. J. Am. Chem. Soc. 2022, 144, 13218–13225. [Google Scholar] [CrossRef]
- Dou, Q.; Wei, X.; Zhou, K.; Yang, S.; Jia, P. Cardiovascular Manifestations and Mechanisms in Patients with COVID-19. Trends Endocrinol. Metab. 2020, 31, 893–904. [Google Scholar] [CrossRef]
- Carvelli, J.; Demaria, O.; Vely, F.; Batista, L.; Chouaki Benmansour, N.; Fares, J.; Carpentier, S.; Thibult, M.L.; Morel, A.; Remark, R.; et al. Association of COVID-19 inflammation with activation of the C5a-C5aR1 axis. Nature 2020, 588, 146–150. [Google Scholar] [CrossRef]
- Niyonzima, N.; Rahman, J.; Kunz, N.; West, E.E.; Freiwald, T.; Desai, J.V.; Merle, N.S.; Gidon, A.; Sporsheim, B.; Lionakis, M.S.; et al. Mitochondrial C5ar1 Activity in Macrophages Controls IL-1beta Production Underlying Sterile Inflammation. Sci. Immunol. 2021, 6, eabf2489. [Google Scholar] [CrossRef] [PubMed]
- Nording, H.; Baron, L.; Haberthur, D.; Emschermann, F.; Mezger, M.; Sauter, M.; Sauter, R.; Patzelt, J.; Knoepp, K.; Nording, A.; et al. The C5a/C5a receptor 1 Axis Controls Tissue Neovascularization Through CXCL4 Release From Platelets. Nat. Commun. 2021, 12, 3352. [Google Scholar] [CrossRef] [PubMed]
- Abers, M.S.; Delmonte, O.M.; Ricotta, E.E.; Fintzi, J.; Fink, D.L.; de Jesus, A.A.A.; Zarember, K.A.; Alehashemi, S.; Oikonomou, V.; Desai, J.V.; et al. An Immune-Based Biomarker Signature is Associated with Mortality in COVID-19 Patients. JCI Insight 2021, 6, e144455. [Google Scholar] [CrossRef] [PubMed]
- Kroller-Schon, S.; Daiber, A.; Steven, S.; Oelze, M.; Frenis, K.; Kalinovic, S.; Heimann, A.; Schmidt, F.P.; Pinto, A.; Kvandova, M.; et al. Crucial Role for Nox2 and Sleep Deprivation in Aircraft Noise-Induced Vascular and Cerebral Oxidative Stress, Inflammation, and Gene Regulation. Eur. Heart J. 2018, 39, 3528–3539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Violi, F.; Oliva, A.; Cangemi, R.; Ceccarelli, G.; Pignatelli, P.; Carnevale, R.; Cammisotto, V.; Lichtner, M.; Alessandri, F.; De Angelis, M.; et al. Nox2 Activation in COVID-19. Redox. Biol. 2020, 36, 101655. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Kim, J.; Han, J.; Oh, D.; Kim, M.; Jeong, H.; Kim, T.J.; Kim, S.W.; Kim, J.N.; Seo, Y.S.; et al. Formyl Peptide Receptor 2 Determines Sex-Specific Differences in the Progression of Nonalcoholic Fatty Liver Disease and Steatohepatitis. Nat. Commun. 2022, 13, 578. [Google Scholar] [CrossRef] [PubMed]
- Baaten, C.; Meacham, S.; de Witt, S.M.; Feijge, M.A.H.; Adams, D.J.; Akkerman, J.N.; Cosemans, J.; Grassi, L.; Jupe, S.; Kostadima, M.; et al. A Synthesis Approach of Mouse Studies to Identify Genes and Proteins in Arterial Thrombosis and Bleeding. Blood 2018, 132, e35–e46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, M.G.; Nguyen, K.H.; Ball, J.B.; Hopkins, S.; Kelley, T.; Baratta, M.V.; Fleshner, M.; Maier, S.F. SARS-CoV-2 Spike S1 Subunit Induces Neuroinflammatory, Microglial And Behavioral Sickness Responses: Evidence of PAMP-like Properties. Brain Behav. Immun. 2022, 100, 267–277. [Google Scholar] [CrossRef]
- Johnson, M.G.; Puenpatom, A.; Moncada, P.A.; Burgess, L.; Duke, E.R.; Ohmagari, N.; Wolf, T.; Bassetti, M.; Bhagani, S.; Ghosn, J.; et al. Effect of Molnupiravir on Biomarkers, Respiratory Interventions, and Medical Services in COVID-19: A Randomized, Placebo-Controlled Trial. Ann. Intern. Med. 2022, 175, 1126–1134. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Y.; Flahault, A.; Chavarot, N.; Melenotte, C.; Cheminant, M.; Deschamps, P.; Carlier, N.; Lafont, E.; Thomas, M.; Flamarion, E.; et al. Pre-exposure Prophylaxis with Tixagevimab and Cilgavimab (Evusheld) for COVID-19 among 1112 Severely Immunocompromised Patients. Clin. Microbiol. Infect. 2022, 28, 1654. [Google Scholar] [CrossRef]
- Malden, D.E.; Hong, V.; Lewin, B.J.; Ackerson, B.K.; Lipsitch, M.; Lewnard, J.A.; Tartof, S.Y. Hospitalization and Emergency Department Encounters for COVID-19 After Paxlovid Treatment-California, December 2021–May 2022. MMWR Morb Mortal Wkly Rep 2022, 71, 830–833. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K. Molnupiravir Might Be Dangerous without Clarification of Its Indications. BMJ 2022, 377, o1030. [Google Scholar] [CrossRef] [PubMed]
- Charness, M.E.; Gupta, K.; Stack, G.; Strymish, J.; Adams, E.; Lindy, D.C.; Mohri, H.; Ho, D.D. Rebound of SARS-CoV-2 Infection after Nirmatrelvir-Ritonavir Treatment. N. Engl. J. Med. 2022, 387, 1045–1047. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Moller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045.e1039. [Google Scholar] [CrossRef] [PubMed]
- Varga, Z.; Flammer, A.J.; Steiger, P.; Haberecker, M.; Andermatt, R.; Zinkernagel, A.S.; Mehra, M.R.; Schuepbach, R.A.; Ruschitzka, F.; Moch, H. Endothelial Cell Infection and Endotheliitis in COVID-19. Lancet 2020, 395, 1417–1418. [Google Scholar] [CrossRef] [PubMed]
- Tomar, B.; Anders, H.J.; Desai, J.; Mulay, S.R. Neutrophils and Neutrophil Extracellular Traps Drive Necroinflammation in COVID-19. Cells 2020, 9, 1383. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MNC | EPC | DMNC | Degree | Closeness |
|---|---|---|---|---|
| TLR4 | TLR4 | ARG1 | TLR4 | TLR4 |
| TLR2 | TLR2 | AIF1 | TLR2 | TLR2 |
| MMP9 | MMP9 | THBD | MMP9 | MMP9 |
| CD163 | CD163 | CSF1R | CSF1R | CSF1R |
| CSF1R | CSF1R | CD163 | CD163 | CD163 |
| CYBB | AIF1 | CYBB | CYBB | CYBB |
| VWF | CYBB | VWF | VWF | VWF |
| AIF1 | VWF | FPR2 | AIF1 | AIF1 |
| ARG1 | ARG1 | BCL2A1 | ARG1 | ARG1 |
| THBD | THBD | IL1R2 | THBD | THBD |
| C5AR1 | C5AR1 | C5AR1 | C5AR1 | C5AR1 |
| FPR2 | FPR2 | MMP9 | MME | FPR2 |
| BCL2A1 | IL1R2 | TLR2 | FPR2 | IL1R2 |
| HBEGF | HBEGF | TLR4 | HBEGF | BCL2A1 |
| IL1R2 | BCL2A1 | HBEGF | NAMPT | BCL6 |
| Gene Symbol | Protein | CD Antigen Name | Function |
|---|---|---|---|
| C5AR1 | C5a anaphylatoxin chemotactic receptor 1 | CD88 | Receptor for the chemotactic and inflammatory peptide anaphylatoxin C5a |
| MMP9 | Matrix metalloproteinase-9 | Matrix metalloproteinase that plays an essential role in local proteolysis of the extracellular matrix and in leukocyte migration | |
| CYBB | Cytochrome b-245 heavy chain | Critical component of the membrane-bound oxidase of phagocytes that generates superoxide. | |
| FPR2 | N-fo-myl peptide receptor 2 | Low affinity receptor for N-formyl-methionyl peptides, which are powerful neutrophil chemotactic factors | |
| CSF1R | Macrophage colony-stimulating factor 1 receptor | CD115 | Tyrosine-protein kinase that acts as a cell-surface receptor for CSF1 and IL34 and plays an essential role in the regulation of survival, proliferation, and differentiation of hematopoietic precursor cells, especially mononuclear phagocytes, such as macrophages and monocytes. |
| TLR2 | Toll-like receptor 2 | CD282 | Cooperates with LY96 to mediate the innate immune response to bacterial lipoproteins and other microbial cell wall components. |
| TLR4 | Toll-like receptor 4 | CD284 | Cooperates with LY96 and CD14 to mediate the innate immune response to bacterial lipopolysaccharide (LPS) |
| Rank | Score | Name | Description |
|---|---|---|---|
| 1 | −99.93 | Solanine | Acetylcholinesterase inhibitor |
| 2 | −99.89 | Desoxypeganine | Acetylcholinesterase inhibitor |
| 3 | −99.86 | Alpha-linolenic-acid | Omega 3 fatty acid stimulant |
| 4 | −99.82 | CAY-10577 | Casein kinase inhibitor |
| 5 | −99.79 | Homochlorcyclizine | Antihistamine |
| 6 | −99.75 | Altretamine | DNA synthesis inhibitor |
| 7 | −99.71 | NU-1025 | PARP inhibitor |
| 8 | −99.65 | TG100-115 | PI3Kγ/PI3Kδ inhibitor |
| 9 | −99.62 | Raltegravir | HIV integrase inhibitor |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Y.; Zhang, J.; Xu, L.; Zhang, H.; Ma, S.; Gao, Y.; Jiang, Y. Developing a Novel Immune-Related Seven-Gene Signature and Immune Infiltration Pattern in Patients with COVID-19 and Cardiovascular Disease. J. Cardiovasc. Dev. Dis. 2022, 9, 450. https://doi.org/10.3390/jcdd9120450
Fu Y, Zhang J, Xu L, Zhang H, Ma S, Gao Y, Jiang Y. Developing a Novel Immune-Related Seven-Gene Signature and Immune Infiltration Pattern in Patients with COVID-19 and Cardiovascular Disease. Journal of Cardiovascular Development and Disease. 2022; 9(12):450. https://doi.org/10.3390/jcdd9120450
Chicago/Turabian StyleFu, Yajuan, Juan Zhang, Lingbo Xu, Hui Zhang, Shengchao Ma, Yujing Gao, and Yideng Jiang. 2022. "Developing a Novel Immune-Related Seven-Gene Signature and Immune Infiltration Pattern in Patients with COVID-19 and Cardiovascular Disease" Journal of Cardiovascular Development and Disease 9, no. 12: 450. https://doi.org/10.3390/jcdd9120450
APA StyleFu, Y., Zhang, J., Xu, L., Zhang, H., Ma, S., Gao, Y., & Jiang, Y. (2022). Developing a Novel Immune-Related Seven-Gene Signature and Immune Infiltration Pattern in Patients with COVID-19 and Cardiovascular Disease. Journal of Cardiovascular Development and Disease, 9(12), 450. https://doi.org/10.3390/jcdd9120450