


Diversity of Mycorrhizal Fungi in Temperate Orchid Species: Comparison of Culture-Dependent and Culture-Independent Methods

Abstract
:1. Introduction
2. Materials and Methods
2.1. Orchid Species and Root Collection
2.2. Root Preparation for Identification of Fungal-Root Associates
2.3. Culture-Dependent and -Independent Assessment of OMF
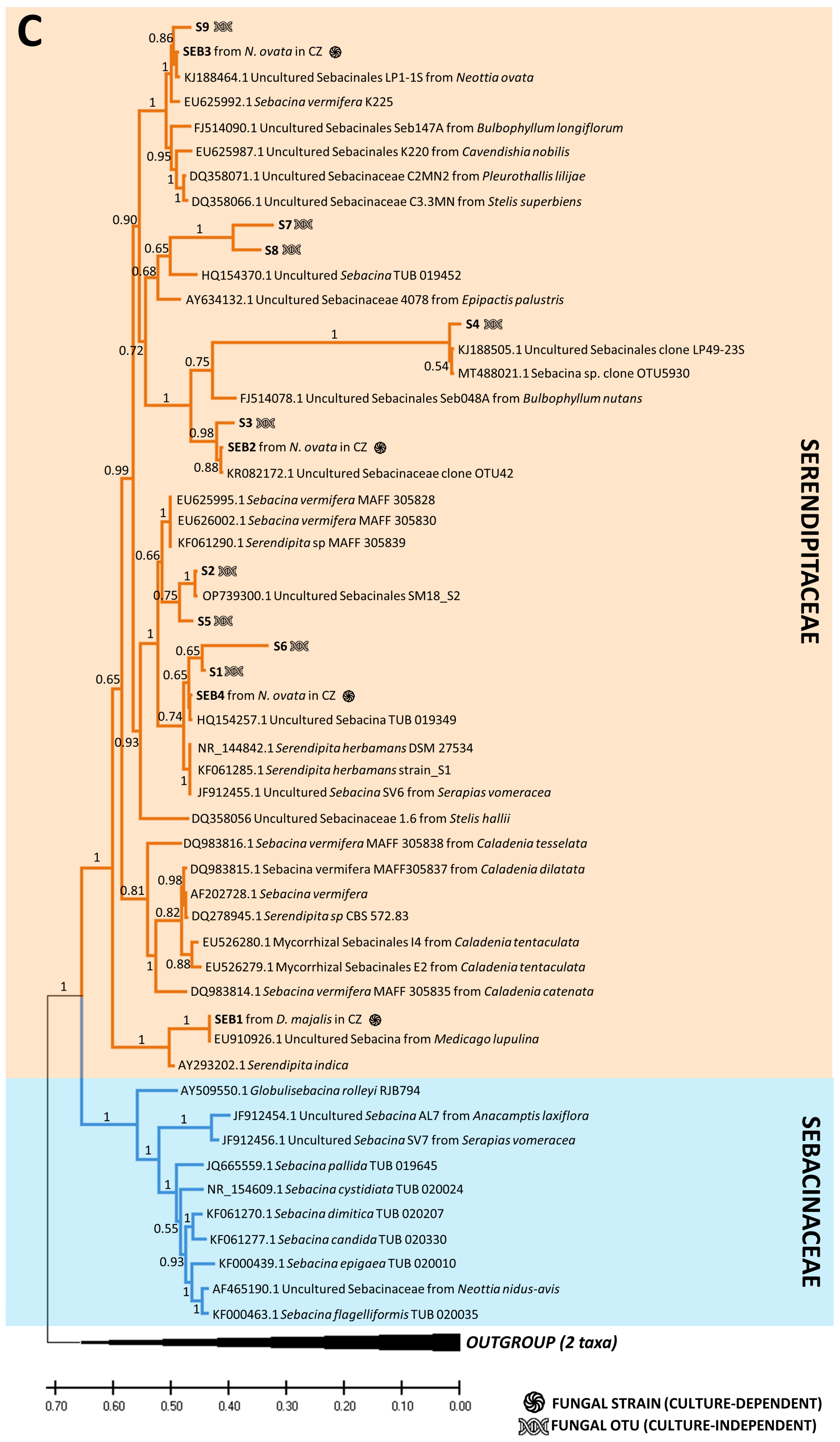
2.4. Phylogenetic Trees of Fungal OTUs Obtained from Both Methods
2.5. Data Analysis
3. Results
3.1. Culture-Independent Method: Diversity and Community Composition of Rhizoctonia across Orchid Species and Sites
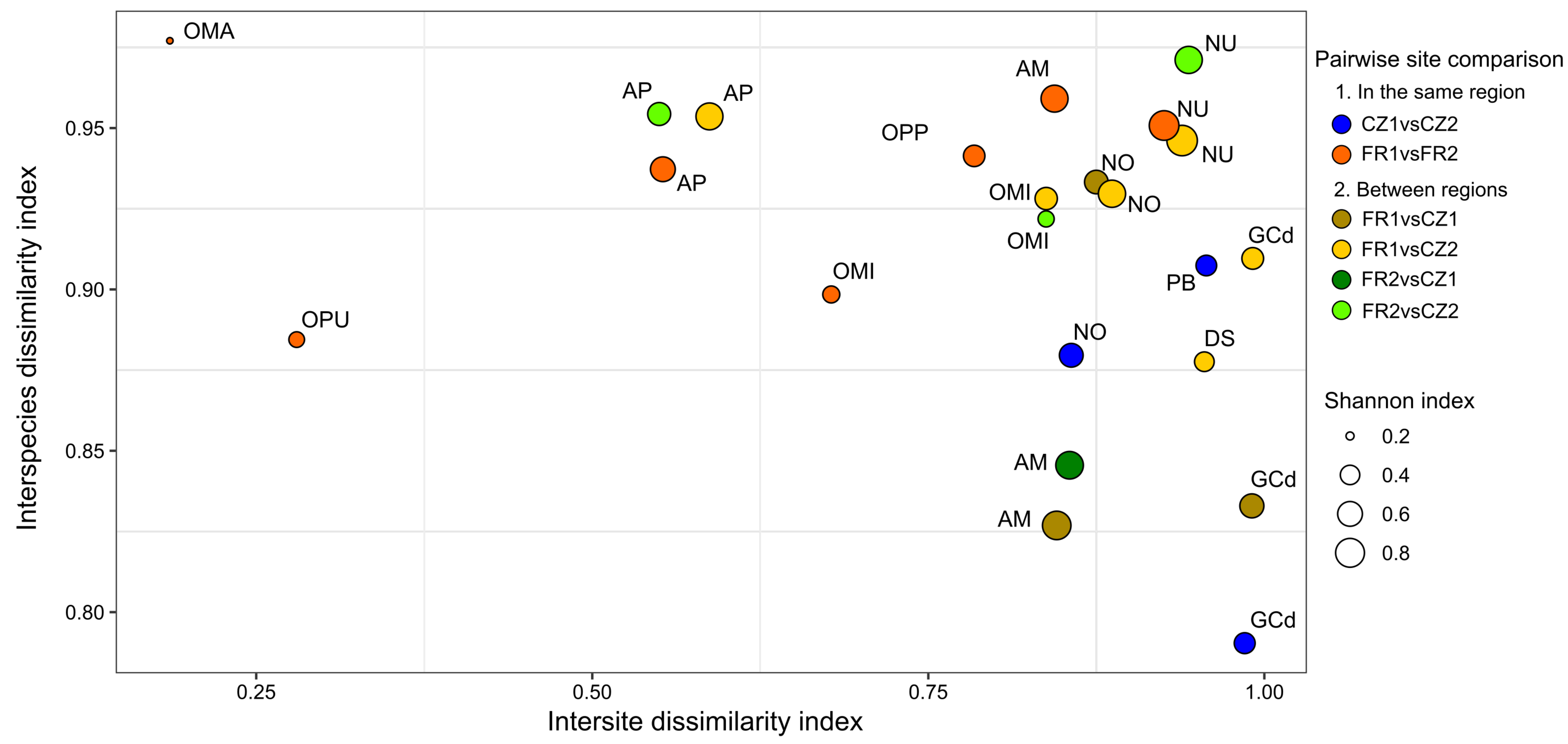
3.2. Comparison of OMF Diversity Using Culture-Dependent and -Independent Methods
4. Discussion
4.1. Orchid Mycorrhizal Fungi Diversity among Orchid Species
4.2. Effect of Locality and Host Species Identity on Fungal Composition
4.3. Combining Culture-Dependent and -Independent Methods
4.4. Perspectives for Future Research
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chase, M.W.; Cameron, K.M.; Freudenstein, J.V.; Pridgeon, A.M.; Salazar, G.; van den Berg, C.; Schuiteman, A. An updated classification of Orchidaceae: Updated Classification of Orchidaceae. Bot. J. Linn. Soc. 2015, 177, 151–174. [Google Scholar] [CrossRef]
- Swarts, N.D.; Dixon, K.W. Terrestrial orchid conservation in the age of extinction. Ann. Bot. 2009, 104, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Willis, K. State of the World’s Plants 2017; Royal Botanics Gardens Kew: Richmond, UK, 2017. [Google Scholar]
- Kull, T.; Hutchings, M.J. A comparative analysis of decline in the distribution ranges of orchid species in Estonia and the United Kingdom. Biol. Conserv. 2006, 129, 31–39. [Google Scholar] [CrossRef]
- Rasmussen, H.N.; Dixon, K.W.; Jersáková, J.; Těšitelová, T. Germination and seedling establishment in orchids: A complex of requirements. Ann. Bot. 2015, 116, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Fay, M.F. Orchid conservation: How can we meet the challenges in the twenty-first century? Bot. Stud. 2018, 59, 16. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- McCormick, M.K.; Jacquemyn, H. What constrains the distribution of orchid populations? New Phytol. 2014, 202, 392–400. [Google Scholar] [CrossRef]
- Dearnaley, J.D.W.; Martos, F.; Selosse, M.-A. 12 Orchid Mycorrhizas: Molecular Ecology, Physiology, Evolution and Conservation Aspects. In Fungal Associations; Hock, B., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 207–230. [Google Scholar]
- Rasmussen, H.N. Terrestrial Orchids: From Seed to Mycotrophic Plant, 1st ed.; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Rasmussen, H.N. Recent developments in the study of orchid mycorrhiza. Plant Soil 2002, 244, 149–163. [Google Scholar] [CrossRef]
- Taylor, D.L.; Bruns, T.D.; Leake, J.R.; Read, D.J. Mycorrhizal Specificity and Function in Myco-heterotrophic Plants. In Mycorrhizal Ecology; Van Der Heijden, M.G.A., Sanders, I.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; Volume 157, pp. 375–413. [Google Scholar]
- Weiß, M.; Waller, F.; Zuccaro, A.; Selosse, M. Sebacinales—One thousand and one interactions with land plants. New Phytol. 2016, 211, 20–40. [Google Scholar] [CrossRef]
- Kohler, A.; Kuo, A.; Nagy, L.G.; Morin, E.; Barry, K.W.; Buscot, F.; Canbäck, B.; Choi, C.; Cichocki, N.; Clum, A.; et al. Convergent losses of decay mechanisms and rapid turnover of symbiosis genes in mycorrhizal mutualists. Nat. Genet. 2015, 47, 410–415. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Duffy, K.J.; Selosse, M.-A. Biogeography of Orchid Mycorrhizas. In Biogeography of Mycorrhizal Symbiosis; Tedersoo, L., Ed.; Springer International Publishing: New York, NY, USA, 2017; Volume 230, pp. 159–177. [Google Scholar]
- Girlanda, M.; Segreto, R.; Cafasso, D.; Liebel, H.T.; Rodda, M.; Ercole, E.; Cozzolino, S.; Gebauer, G.; Perotto, S. Photosynthetic Mediterranean meadow orchids feature partial mycoheterotrophy and specific mycorrhizal associations. Am. J. Bot. 2011, 98, 1148–1163. [Google Scholar] [CrossRef]
- Kohout, P.; Těšitelová, T.; Roy, M.; Vohník, M.; Jersáková, J. A diverse fungal community associated with Pseudorchis albida (Orchidaceae) roots. Fungal Ecol. 2013, 6, 50–64. [Google Scholar] [CrossRef]
- Selosse, M.-A.; Faccio, A.; Scappaticci, G.; Bonfante, P. Chlorophyllous and Achlorophyllous Specimens of Epipactis microphylla (Neottieae, Orchidaceae) Are Associated with Ectomycorrhizal Septomycetes, including Truffles. Microb. Ecol. 2004, 47, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Merckx, V.S.F.T. Mycoheterotrophy: An Introduction. In Mycoheterotrophy: The Biology of Plants Living on Fungi; Merckx, V., Ed.; Springer: New York, NY, USA, 2013; pp. 1–17. [Google Scholar]
- Wang, D.; Lerou, J.; Nuytinck, J.; Gomes, S.I.F.; Jacquemyn, H.; Merckx, V.S.F.T. Root-associated Fungi in Orchidaceae: Diversity, Phylogeny, Ecology, and Outstanding Questions. bioRxiv, 2022; preprint. [Google Scholar]
- Selosse, M.-A.; Roy, M. Green plants that feed on fungi: Facts and questions about mixotrophy. Trends Plant Sci. 2009, 14, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Öpik, M.; Moora, M. Missing nodes and links in mycorrhizal networks. New Phytol. 2012, 194, 304–306. [Google Scholar] [CrossRef] [PubMed]
- Těšitelová, T.; Jersáková, J.; Roy, M.; Kubátová, B.; Těšitel, J.; Urfus, T.; Trávníček, P.; Suda, J. Ploidy-specific symbiotic interactions: Divergence of mycorrhizal fungi between cytotypes of the Gymnadenia conopsea group (Orchidaceae). New Phytol. 2013, 199, 1022–1033. [Google Scholar] [CrossRef] [PubMed]
- Vogt-Schilb, H.; Těšitelová, T.; Kotilínek, M.; Sucháček, P.; Kohout, P.; Jersáková, J. Altered rhizoctonia assemblages in grasslands on ex-arable land support germination of mycorrhizal generalist, not specialist orchids. New Phytol. 2020, 227, 1200–1212. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Honnay, O.; Cammue, B.P.A.; Brys, R.; Lievens, B. Low specificity and nested subset structure characterize mycorrhizal associations in five closely related species of the genus. Orchis. Mol. Ecol. 2010, 19, 4086–4095. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Waud, M.; Lievens, B.; Brys, R. Differences in mycorrhizal communities between Epipactis palustris, E. helleborine and its presumed sister species E. neerlandica. Ann. Bot. 2016, 118, 105–114. [Google Scholar] [CrossRef]
- Perez-Lamarque, B.; Öpik, M.; Maliet, O.; Afonso Silva, A.C.; Selosse, M.-A.; Martos, F.; Morlon, H. Analysing diversification dynamics using barcoding data: The case of an obligate mycorrhizal symbiont. Mol. Ecol. 2022, 31, 3496–3512. [Google Scholar] [CrossRef]
- Yun, W.; Hall, I.R. Edible ectomycorrhizal mushrooms: Challenges and achievements. Can. J. Bot. 2004, 82, 1063–1073. [Google Scholar] [CrossRef]
- Zettler, L.W.; Corey, L.L. Orchid Mycorrhizal Fungi: Isolation and Identification Techniques. In Orchid Propagation: From Laboratories to Greenhouses—Methods and Protocols; Lee, Y.-I., Yeung, E.C.-T., Eds.; Springer: New York, NY, USA, 2018; pp. 27–59. [Google Scholar]
- Nilsson, R.H.; Anslan, S.; Bahram, M.; Wurzbacher, C.; Baldrian, P.; Tedersoo, L. Mycobiome diversity: High-throughput sequencing and identification of fungi. Nat. Rev. Microbiol. 2019, 17, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Bahram, M.; Zinger, L.; Nilsson, R.H.; Kennedy, P.G.; Yang, T.; Anslan, S.; Mikryukov, V. Best practices in metabarcoding of fungi: From experimental design to results. Mol. Ecol. 2022, 31, 2769–2795. [Google Scholar] [CrossRef] [PubMed]
- Alomía, Y.A.; Otero, J.T.; Jersáková, J.; Stevenson, P.R. Cultivable fungal community associated with the tropical orchid Dichaea andina. Fungal Ecol. 2022, 57, 101158. [Google Scholar] [CrossRef]
- Schwartz, S.; Oren, R.; Ast, G. Detection and Removal of Biases in the Analysis of Next-Generation Sequencing Reads. PLoS ONE 2011, 6, e16685. [Google Scholar] [CrossRef] [PubMed]
- Rammitsu, K.; Kajita, T.; Imai, R.; Ogura-Tsujita, Y. Strong primer bias for Tulasnellaceae fungi in metabarcoding: Specific primers improve the characterization of the mycorrhizal communities of epiphytic orchids. Mycoscience 2021, 62, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Smith, D.; Peay, K.; Kennedy, P. Parsing ecological signal from noise in next generation amplicon sequencing. New Phytol. 2015, 205, 1389–1393. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Bayman, P. Mycorrhizal Fungi of Vanilla: Diversity, Specificity and Effects on Seed Germination and Plant Growth. Mycologia 2007, 99, 510–525. [Google Scholar] [CrossRef]
- Mennicken, S.; Vogt-Schilb, H.; Těšitelová, T.; Kotilínek, M.; Alomía, Y.A.; Schatz, B.; Jersáková, J. Orchid–mycorrhizal fungi interactions reveal a duality in their network structure in two European regions differing in climate. Mol. Ecol. 2023, 32, 3308–3321. [Google Scholar] [CrossRef]
- Wang, H.; Qi, M.; Cutler, A.J. A simple method of preparing plant samples for PCR. Nucl. Acids Res. 1993, 21, 4153–4154. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: New York, NY, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Taylor, D.L.; McCormick, M.K. Internal transcribed spacer primers and sequences for improved characterization of basidiomycetous orchid mycorrhizas. New Phytol. 2008, 177, 1020–1033. [Google Scholar] [CrossRef] [PubMed]
- Gabriadze, I.; Kutateladze, T.; Vishnepolsky, B.; Karseladze, M.; Datukishvili, N. Application of PCR-based methods for rapid detection of corn ingredients in processed foods. Int. J. Food Sci. Nutr. 2014, 3, 199–202. [Google Scholar] [CrossRef]
- Li, T.; Yang, W.; Wu, S.; Selosse, M.-A.; Gao, J. Progress and Prospects of Mycorrhizal Fungal Diversity in Orchids. Front. Plant Sci. 2021, 12, 646325. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucl. Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE: New York, NY, USA, 2010; pp. 1–8. [Google Scholar]
- Rambaut, A. FigTree, v1.3.1; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, UK, 2010.
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R package version 2.5-7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 9 September 2023).
- Jacquemyn, H.; Brys, R.; Waud, M.; Busschaert, P.; Lievens, B. Mycorrhizal networks and coexistence in species-rich orchid communities. New Phytol. 2015, 206, 1127–1134. [Google Scholar] [CrossRef]
- Těšitelová, T.; Kotilínek, M.; Jersáková, J.; Joly, F.; Košnar, J.; Tatarenko, I.; Selosse, M. Two widespread green Neottia species (Orchidaceae) show mycorrhizal preference for Sebacinales in various habitats and ontogenetic stages. Mol. Ecol. 2015, 24, 1122–1134. [Google Scholar] [CrossRef]
- Djordjević, V.; Tsiftsis, S.; Lakušić, D.; Jovanović, S.; Stevanović, V. Factors affecting the distribution and abundance of orchids in grasslands and herbaceous wetlands. Syst. Biodivers. 2016, 14, 355–370. [Google Scholar] [CrossRef]
- Bailarote, B.C.; Lievens, B.; Jacquemyn, H. Does mycorrhizal specificity affect orchid decline and rarity? Am. J. Bot. 2012, 99, 1655–1665. [Google Scholar] [CrossRef] [PubMed]
- Kotilínek, M.; Těšitelová, T.; Jersáková, J. Biological Flora of the British Isles: Neottia ovata. J. Ecol. 2015, 103, 1354–1366. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Merckx, V.; Brys, R.; Tyteca, D.; Cammue, B.P.A.; Honnay, O.; Lievens, B. Analysis of network architecture reveals phylogenetic constraints on mycorrhizal specificity in the genus Orchis (Orchidaceae). New Phytol. 2011, 192, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, W.R. Diversity and Roles of Mycorrhizal Fungi in the Bee Orchid Ophrys apifera; The University of Liverpool: Liverpool, UK, 2018. [Google Scholar]
- Selosse, M.A.; Martos, F.; Perry, B.; Maj, P.; Roy, M.; Pailler, T. Saprotrophic fungal symbionts in tropical achlorophyllous orchids: Finding treasures among the ‘molecular scraps’? Plant Signal. Behav. 2010, 5, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Yagame, T.; Funabiki, E.; Nagasawa, E.; Fukiharu, T.; Iwase, K. Identification and symbiotic ability of Psathyrellaceae fungi isolated from a photosynthetic orchid, Cremastra appendiculata (Orchidaceae). Am. J. Bot. 2013, 100, 1823–1830. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, J.; Lv, Y.; Gao, C.; Guo, S. Mycena sp., a mycorrhizal fungus of the orchid Dendrobium officinale. Mycol. Progress 2012, 11, 395–401. [Google Scholar] [CrossRef]
- Looney, B.P.; Meidl, P.; Piatek, M.J.; Miettinen, O.; Martin, F.M.; Matheny, P.B.; Labbé, J.L. Russulaceae: A new genomic dataset to study ecosystem function and evolutionary diversification of ectomycorrhizal fungi with their tree associates. New Phytol. 2018, 218, 54–65. [Google Scholar] [CrossRef]
- Girlanda, M.; Selosse, M.A.; Cafasso, D.; Brilli, F.; Delfine, S.; Fabbian, R.; Ghignone, S.; Pinelli, P.; Segreto, R.; Loreto, F.; et al. Inefficient photosynthesis in the Mediterranean orchid Limodorum abortivum is mirrored by specific association to ectomycorrhizal Russulaceae. Mol. Ecol. 2006, 15, 491–504. [Google Scholar] [CrossRef]
- Xing, X.; Gao, Y.; Zhao, Z.; Waud, M.; Duffy, K.J.; Selosse, M.; Jakalski, M.; Liu, N.; Jacquemyn, H.; Guo, S. Similarity in mycorrhizal communities associating with two widespread terrestrial orchids decays with distance. J. Biogeogr. 2020, 47, 421–433. [Google Scholar] [CrossRef]
- Těšitelová, T.; Těšitel, J.; Jersáková, J.; Ríhová, G.; Selosse, M. Symbiotic germination capability of four Epipactis species (Orchidaceae) is broader than expected from adult ecology. Am. J. Bot. 2012, 99, 1020–1032. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.L.; Bruns, T.D. Independent, specialized invasions of ectomycorrhizal mutualism by two nonphotosynthetic orchids. Proc. Natl. Acad. Sci. USA 1997, 94, 4510–4515. [Google Scholar] [CrossRef] [PubMed]
- Jacquemyn, H.; Brys, R.; Merckx, V.S.F.T.; Waud, M.; Lievens, B.; Wiegand, T. Coexisting orchid species have distinct mycorrhizal communities and display strong spatial segregation. New Phytol. 2014, 202, 616–627. [Google Scholar] [CrossRef] [PubMed]
- Oja, J.; Kohout, P.; Tedersoo, L.; Kull, T.; Kõljalg, U. Temporal patterns of orchid mycorrhizal fungi in meadows and forests as revealed by 454 pyrosequencing. New Phytol. 2015, 205, 1608–1618. [Google Scholar] [CrossRef] [PubMed]
- Duffy, K.J.; Waud, M.; Schatz, B.; Petanidou, T.; Jacquemyn, H. Latitudinal variation in mycorrhizal diversity associated with a European orchid. J. Biogeogr. 2019, 46, 968–980. [Google Scholar] [CrossRef]
- Cevallos, S.; Herrera, P.; Sánchez-Rodríguez, A.; Declerck, S.; Suárez, J.P. Untangling factors that drive community composition of root associated fungal endophytes of Neotropical epiphytic orchids. Fungal Ecol. 2018, 34, 67–75. [Google Scholar] [CrossRef]
- Kretzchmar, H.; Eccarius, W.; Dietrich, H. The Orchid Genera Anacamptis, Orchis, Neotinea; EchinoMedia-Verlag: Albersdorf, Germany, 2007. [Google Scholar]
- Cruz, D.; Suárez, J.P.; Kottke, I.; Piepenbring, M.; Oberwinkler, F. Defining species in Tulasnella by correlating morphology and nrDNA ITS-5.8S sequence data of basidiomata from a tropical Andean forest. Mycol. Progress 2011, 10, 229–238. [Google Scholar] [CrossRef]
- Calevo, J.; Voyron, S.; Ercole, E.; Girlanda, M. Is the Distribution of Two Rare Orchis Sister Species Limited by Their Main Mycobiont? Diversity 2020, 12, 262. [Google Scholar] [CrossRef]
- Oberwinkler, F.; Cruz, D.; Suárez, J.P. Biogeography and Ecology of Tulasnellaceae. In Biogeography of Mycorrhizal Symbiosis; Tedersoo, L., Ed.; Springer International Publishing: New York, NY, USA, 2017; Volume 230, pp. 237–271. [Google Scholar]
- Nurfadilah, S.; Swarts, N.D.; Dixon, K.W.; Lambers, H.; Merritt, D.J. Variation in nutrient-acquisition patterns by mycorrhizal fungi of rare and common orchids explains diversification in a global biodiversity hotspot. Ann. Bot. 2013, 111, 1233–1241. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Brys, R.; Cammue BP, A.; Honnay, O.; Lievens, B. Mycorrhizal associations and reproductive isolation in three closely related Orchis species. Ann. Bot. 2011, 107, 347–356. [Google Scholar] [CrossRef]
- Lievens, B.; van Kerckhove, S.; Justé, A.; Cammue, B.P.; Honnay, O.; Jacquemyn, H. From extensive clone libraries to comprehensive DNA arrays for the efficient and simultaneous detection and identification of orchid mycorrhizal fungi. J. Microbiol. Methods 2010, 80, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Shefferson, R.P.; Kull, T.; Tali, K. Mycorrhizal interactions of orchids colonizing Estonian mine tailings hills. Am. J. Bot. 2008, 95, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Novotná, A.; Mennicken, S.; de Paula, C.C.P.; Vogt-Schilb, H.; Kotilínek, M.; Těšitelová, T.; Šmilauer, P.; Jersáková, J. Variability in Nutrient Use by Orchid Mycorrhizal Fungi in Two Medium Types. J. Fungi 2023, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.-Y.; Zhang, W.-L.; Selosse, M.-A.; Gao, J.-Y. Are fungi from adult orchid roots the best symbionts at germination? A case study. Mycorrhiza 2019, 29, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Cevallos, S.; Declerck, S.; Suárez, J.P. In situ Orchid Seedling-Trap Experiment Shows Few Keystone and Many Randomly Associated Mycorrhizal Fungal Species during Early Plant Colonization. Front. Plant Sci. 2018, 9, 1664. [Google Scholar] [CrossRef] [PubMed]
- Avis, P.G.; Branco, S.; Tang, Y.; Mueller, G.M. Pooled samples bias fungal community descriptions. Mol. Ecol. Resour. 2010, 10, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; May, T.W.; Smith, M.E. Ectomycorrhizal lifestyle in fungi: Global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 2010, 20, 217–263. [Google Scholar] [CrossRef]
- Wang, D.; Trimbos, K.B.; Gomes, S.I.F.; Jacquemyn, H.; Merckx, V.S.F.T. Metabarcoding read abundances of orchid mycorrhizal fungi are correlated to copy numbers estimated using ddPCR. New Phytol. 2023; Online Version of Record. [Google Scholar]
- Jansa, J.; Mozafar, A.; Frossard, E. Phosphorus acquisition strategies within arbuscular mycorrhizal fungal community of a single field site. Plant Soil. 2005, 276, 163–176. [Google Scholar] [CrossRef]
- Bukovská, P.; Rozmoš, M.; Kotianová, M.; Gančarčíková, K.; Dudáš, M.; Hršelová, H.; Jansa, J. Arbuscular mycorrhiza mediates efficient recycling from soil to plants of nitrogen bound in chitin. Front. Microbiol. 2021, 12, 574060. [Google Scholar] [CrossRef]
- Ventre Lespiaucq, A.; Jacquemyn, H.; Rasmussen, H.N.; Méndez, M. Temporal turnover in mycorrhizal interactions: A proof of concept with orchids. New Phytol. 2021, 230, 1690–1699. [Google Scholar] [CrossRef]
- Lin, G.; McCormack, M.L.; Guo, D. Arbuscular mycorrhizal fungal effects on plant competition and community structure. J. Ecol. 2015, 103, 1224–1232. [Google Scholar] [CrossRef]
- Liu, N.; Jacquemyn, H.; Liu, Q.; Shao, S.C.; Ding, G.; Xing, X. Effects of a dark septate fungal endophyte on the growth and physiological response of seedlings to drought in an epiphytic orchid. Front. Microbiol. 2022, 13, 961172. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Orchid Genera | Orchid Species (Code) | No. of Sampled Plants | No. of Fungal OTUs | Sites |
|---|---|---|---|---|
| Anacamptis | A. morio (AM) * | 27 | 22 | CZ1, FR1, FR2 |
| A. pyramidalis (AP) * | 33 | 18 | CZ2, FR1, FR2 | |
| Dactylorhiza | D. sambucina (DS) * | 14 | 10 | CZ1, FR1 |
| D. viridis (DV) | 5 | 7 | FR1 | |
| Gymnadenia | G. conopsea, diploid (GCd) * | 28 | 23 | CZ1, CZ2, FR1 |
| G. conopsea, tetraploid (GCt) | 9 | 8 | CZ2 | |
| G. densiflora (GD) | 4 | 1 | CZ2 | |
| Neottia | N. ovata (NO) * | 20 | 22 | CZ1, CZ2, FR1 |
| Neotinea | N. ustulata (NU) * | 22 | 24 | CZ2, FR1, FR2 |
| Ophrys | Op. apifera (OPA) | 6 | 4 | CZ2 |
| Op. holubyana (OPH) | 9 | 5 | CZ2 | |
| Op. sphegodes subsp. passionis (OPP) | 17 | 10 | FR1, FR2 | |
| Orchis | O. anthropophora (OA) | 9 | 7 | FR2 |
| O. mascula (OMA) | 16 | 5 | FR1, FR2 | |
| O. militaris (OMI) * | 33 | 16 | CZ2, FR1, FR2 | |
| O. purpurea (OPU) | 18 | 7 | FR1, FR2 | |
| O. simia (OSI) | 8 | 7 | FR2 | |
| Platanthera | P. bifolia (PB) | 15 | 10 | CZ1, CZ2 |
| P. spp. (PL) † | 16 | 7 | FR1, FR2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mennicken, S.; Paula, C.C.P.d.; Vogt-Schilb, H.; Jersáková, J. Diversity of Mycorrhizal Fungi in Temperate Orchid Species: Comparison of Culture-Dependent and Culture-Independent Methods. J. Fungi 2024, 10, 92. https://doi.org/10.3390/jof10020092
Mennicken S, Paula CCPd, Vogt-Schilb H, Jersáková J. Diversity of Mycorrhizal Fungi in Temperate Orchid Species: Comparison of Culture-Dependent and Culture-Independent Methods. Journal of Fungi. 2024; 10(2):92. https://doi.org/10.3390/jof10020092
Chicago/Turabian StyleMennicken, Sophie, Caio César Pires de Paula, Hélène Vogt-Schilb, and Jana Jersáková. 2024. "Diversity of Mycorrhizal Fungi in Temperate Orchid Species: Comparison of Culture-Dependent and Culture-Independent Methods" Journal of Fungi 10, no. 2: 92. https://doi.org/10.3390/jof10020092
APA StyleMennicken, S., Paula, C. C. P. d., Vogt-Schilb, H., & Jersáková, J. (2024). Diversity of Mycorrhizal Fungi in Temperate Orchid Species: Comparison of Culture-Dependent and Culture-Independent Methods. Journal of Fungi, 10(2), 92. https://doi.org/10.3390/jof10020092