Studies on the Properties of the Sporulation Specific Protein Dit1 and Its Product Formyl Tyrosine

Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains
2.2. Plasmids
2.3. Yeast Culture and Sporulation
2.4. In vitro Assays for Dityrosine
2.5. High Performance Liquid Chromatography (HPLC) Analysis
2.6. Western Blot Analysis
2.7. Coimmunoprecipitation
2.8. Microscopy
2.9. Statistics
3. Results
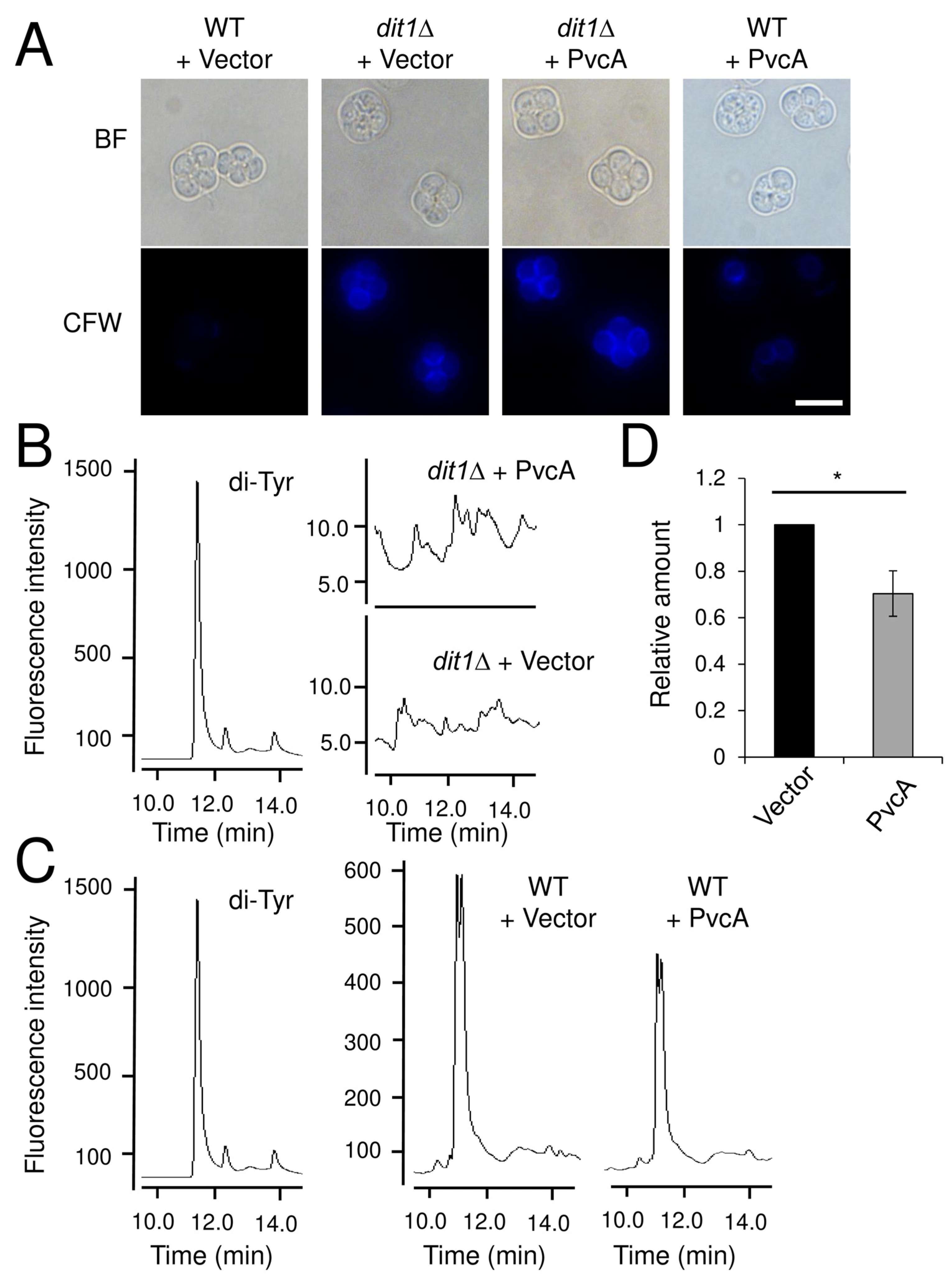
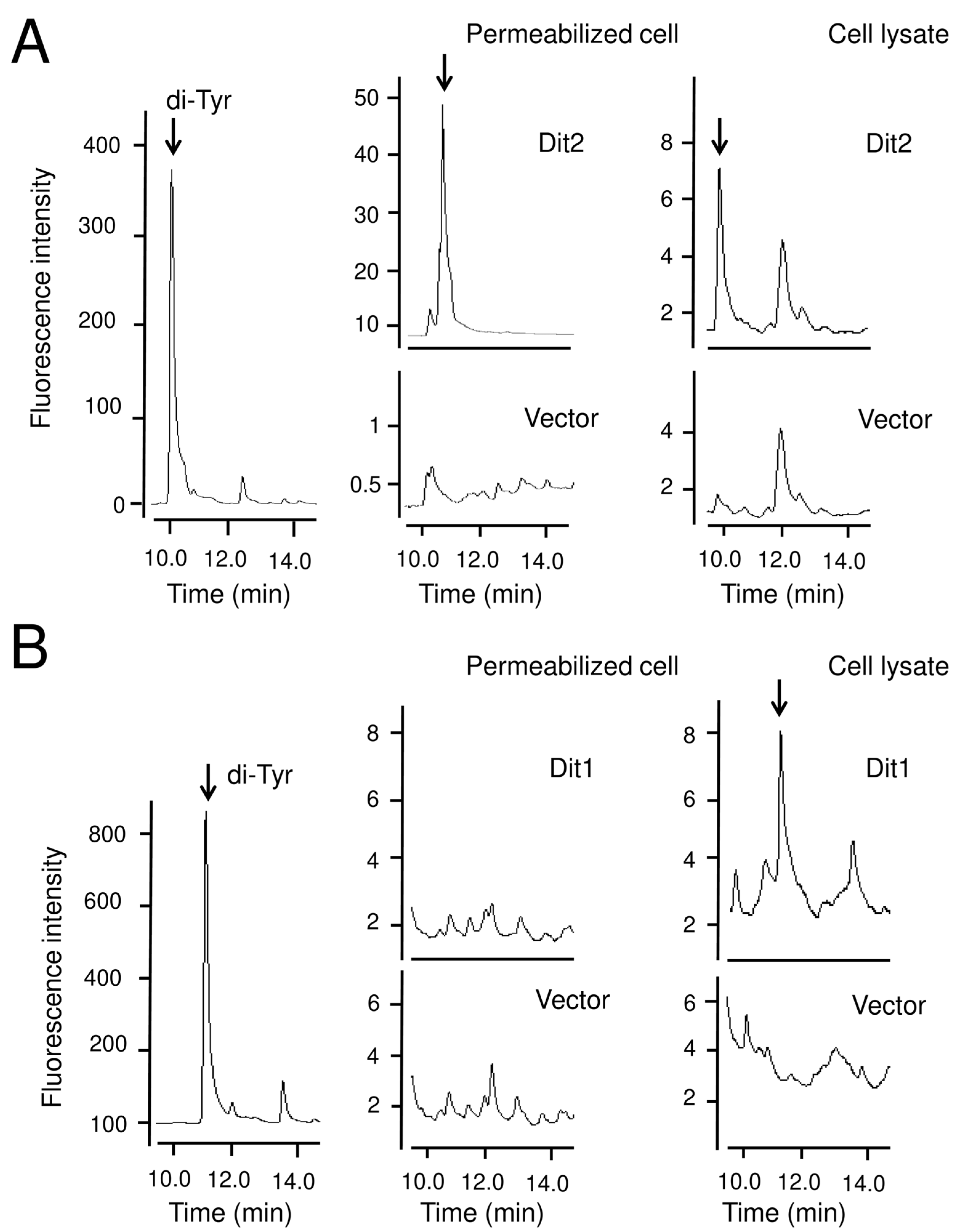
3.1. Dit1 Is not an Isocyanide Synthase
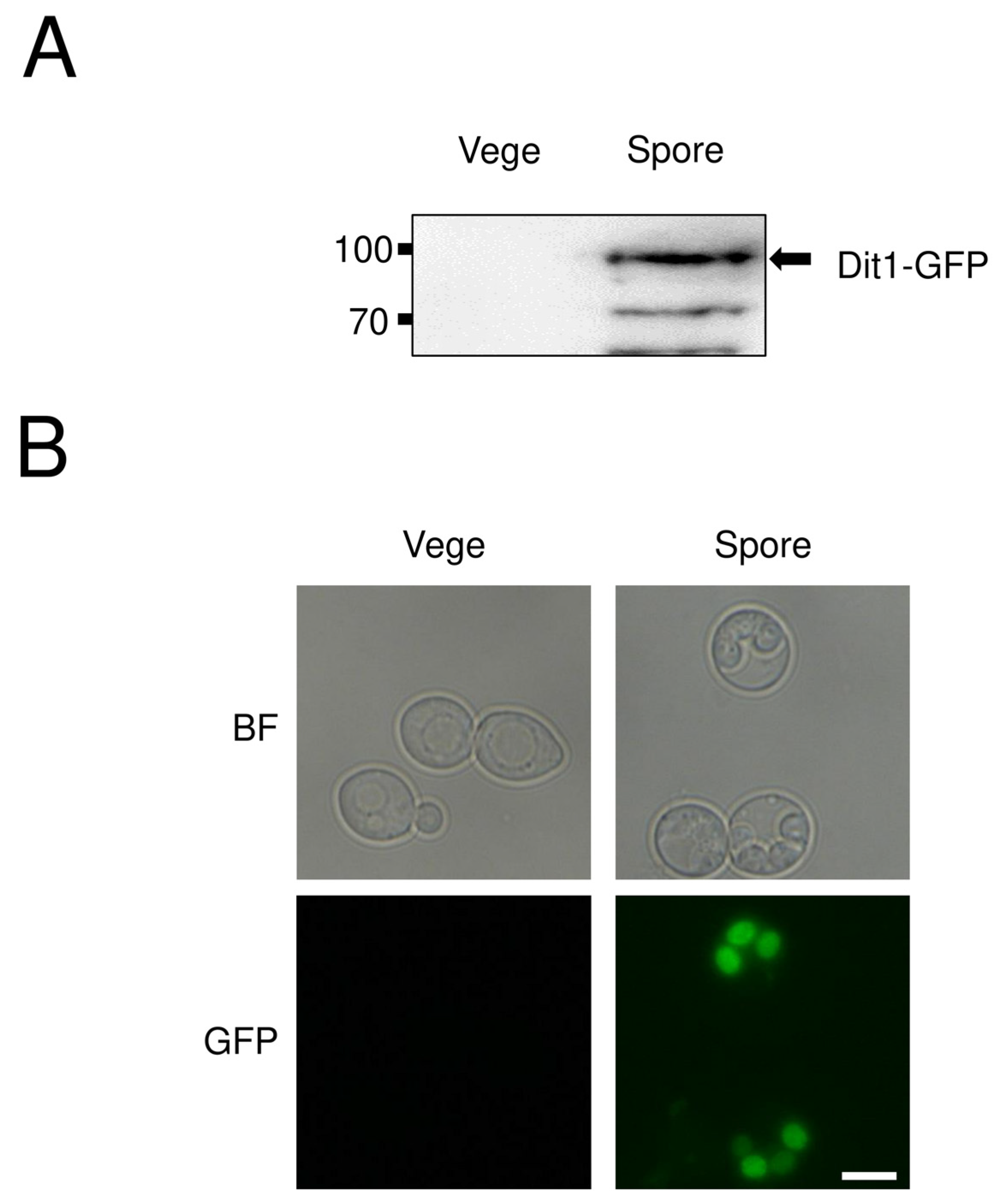
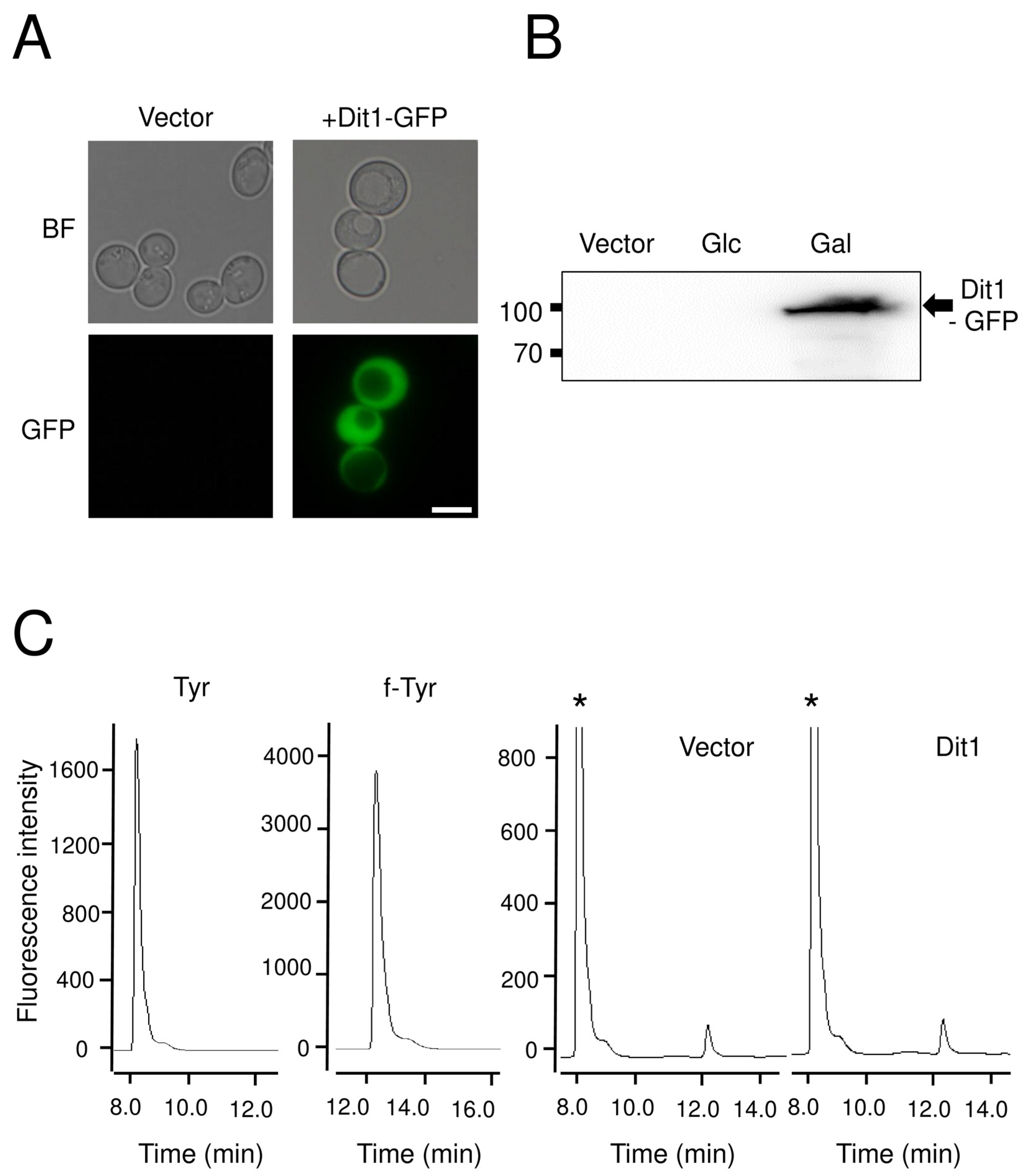
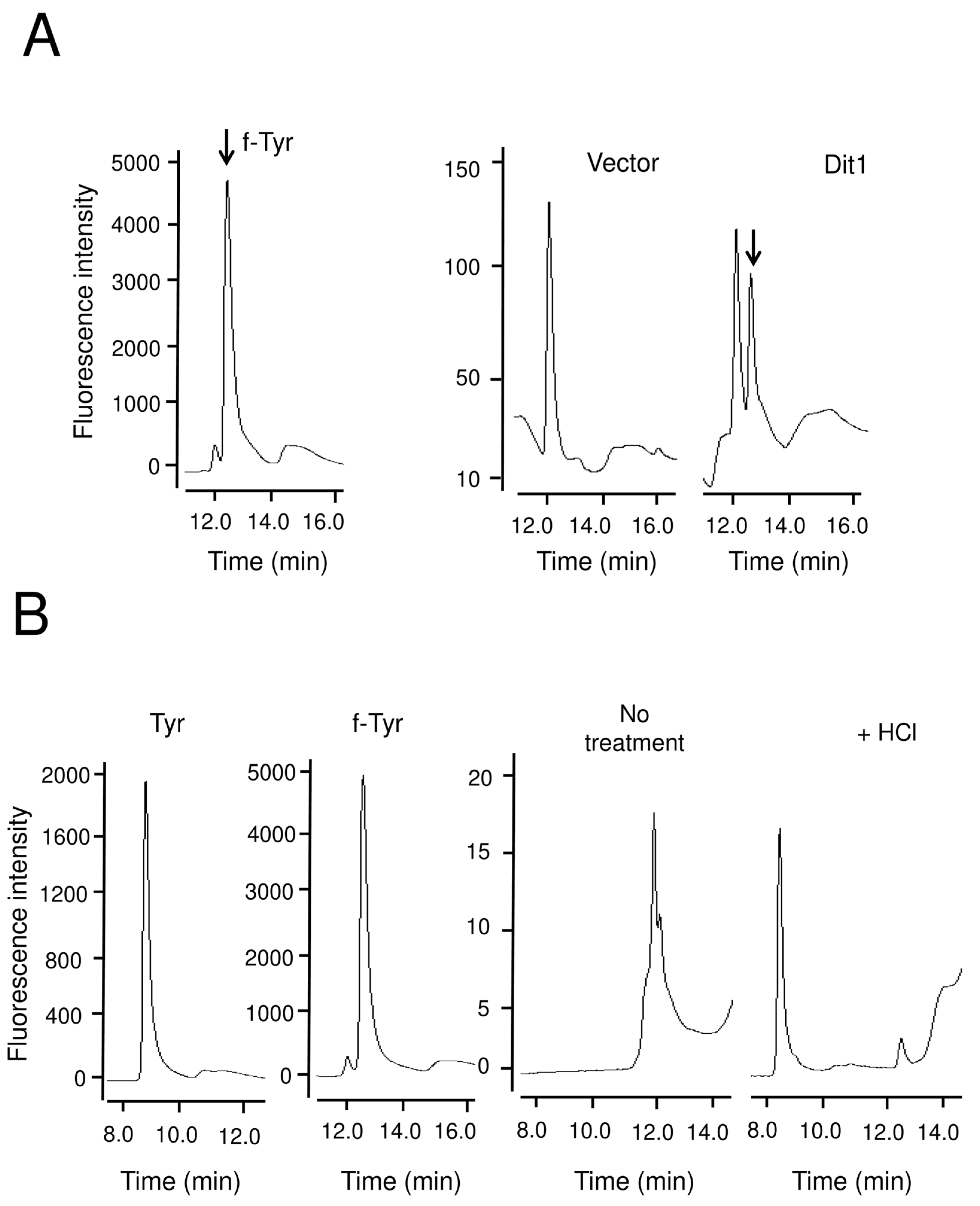
3.2. Dit1 Expressed in Vegetative Cells Was Active but Formyl Tyrosine Was not Detected in the Lysate
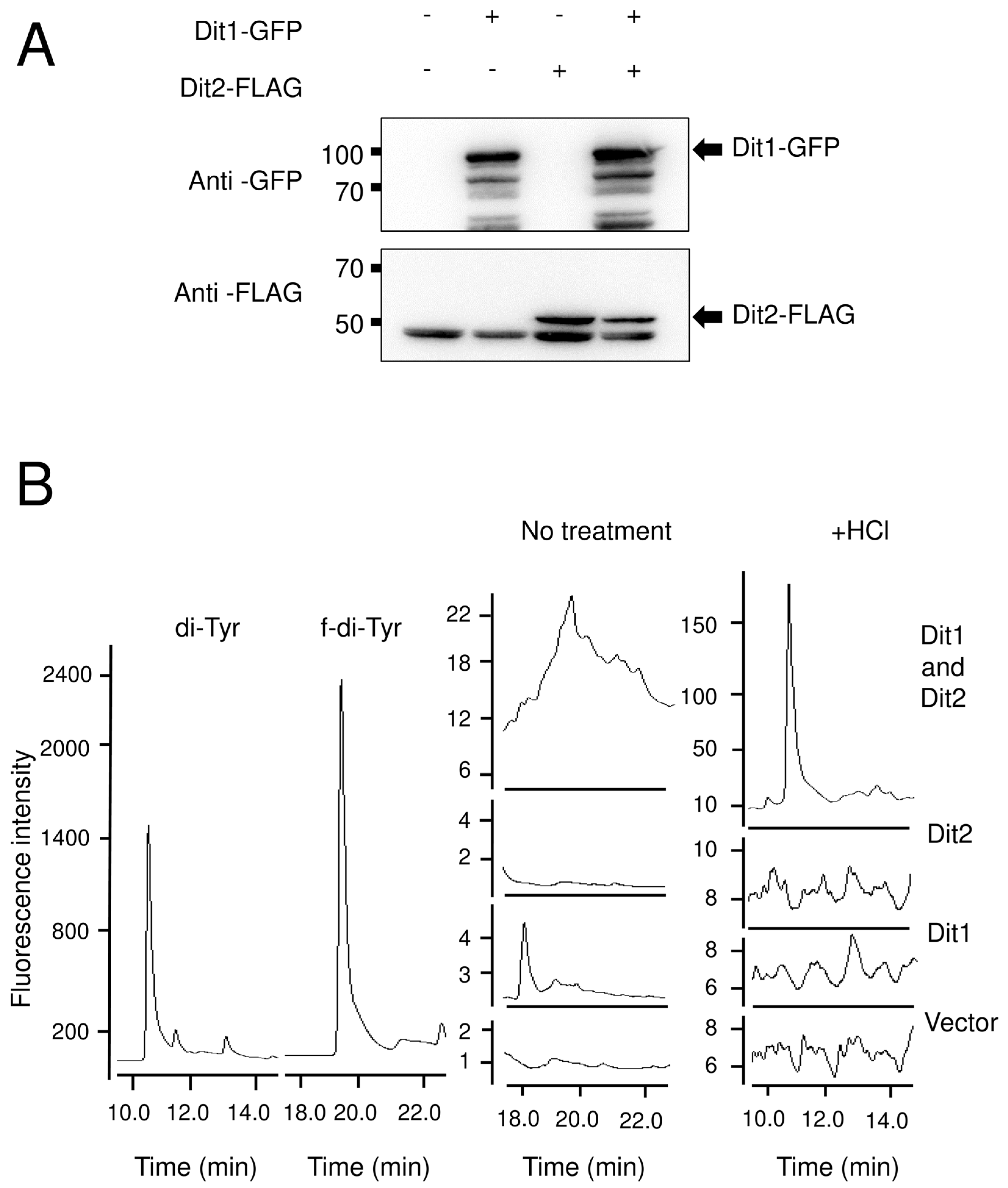
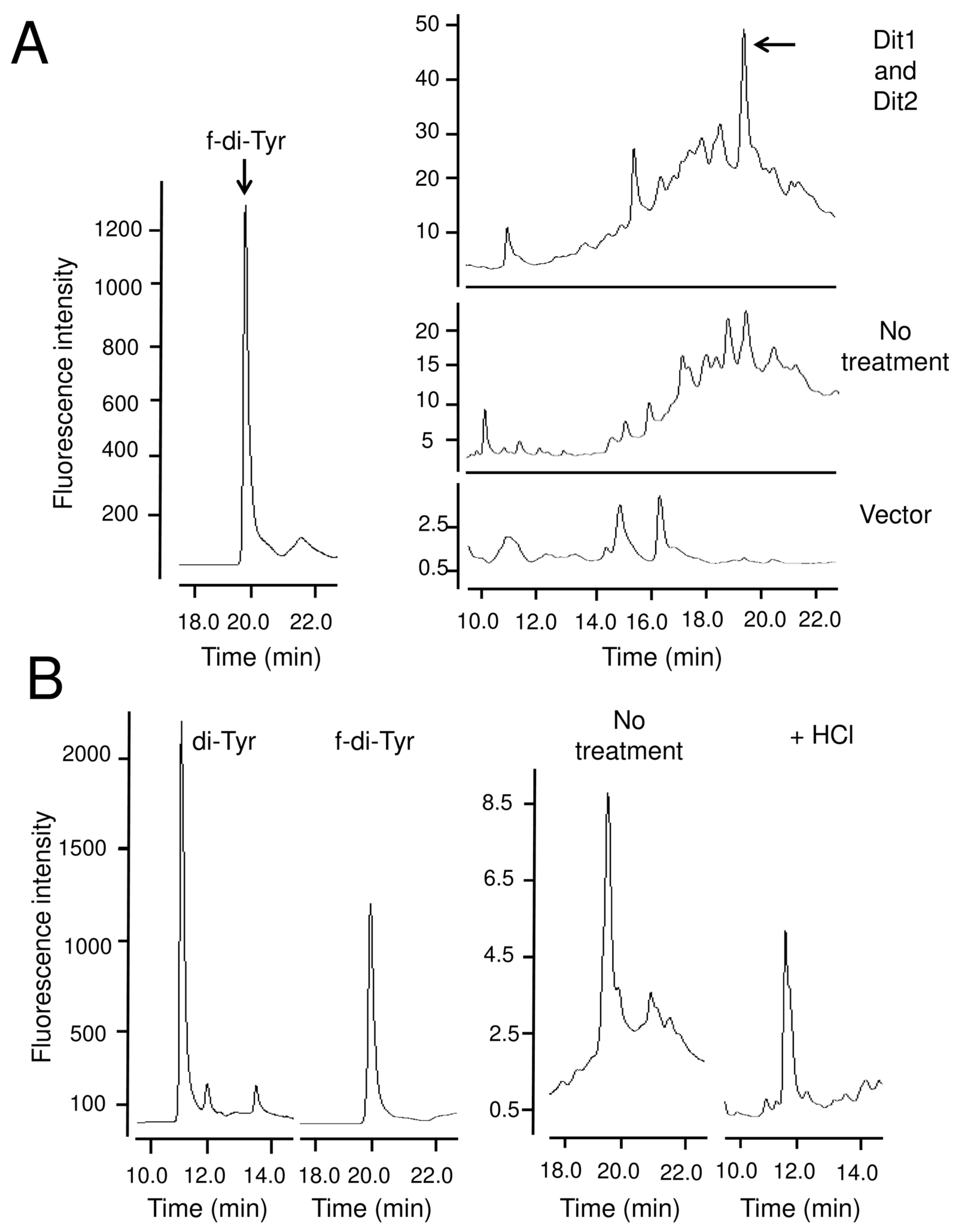
3.3. Dit1 Produces Formyl Tyrosine but It Is not Stable in Yeast Lysate
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Freese, E.B.; Chu, M.I.; Freese, E. Initiation of yeast sporulation of partial carbon, nitrogen, or phosphate deprivation. J. Bacteriol. 1982, 149, 840–851. [Google Scholar] [CrossRef] [Green Version]
- Neiman, A.M. Ascospore formation in the yeast Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2005, 69, 565–584. [Google Scholar] [CrossRef] [Green Version]
- Tachikawa, H.; Bloecher, A.; Tatchell, K.; Neiman, A.M. A Gip1p-Glc7p phosphatase complex regulates septin organization and spore wall formation. J. Cell Biol. 2001, 155, 797–808. [Google Scholar] [CrossRef] [Green Version]
- Briza, P.; Breitenbach, M.; Ellinger, A.; Segall, J. Isolation of two developmentally regulated genes involved in spore wall maturation in Saccharomyces cerevisiae. Genes Dev. 1990, 4, 1775–1789. [Google Scholar] [CrossRef] [Green Version]
- Briza, P.; Kalchhauser, H.; Pittenauer, E.; Allmaier, G.; Breitenbach, M. N,N’-Bisformyl dityrosine is an in vivo precursor of the yeast ascospore wall. Eur. J. Biochem. 1996, 239, 124–131. [Google Scholar] [CrossRef]
- Briza, P.; Eckerstorfer, M.; Breitenbach, M. The sporulation-specific enzymes encoded by the DIT1 and DIT2 genes catalyze a two-step reaction leading to a soluble LL-dityrosine-containing precursor of the yeast spore wall. Proc. Natl. Acad. Sci. USA 1994, 91, 4524–4528. [Google Scholar] [CrossRef] [Green Version]
- Felder, T.; Bogengruber, E.; Tenreiro, S.; Ellinger, A.; Sa-Correia, I.; Briza, P. Dtrlp, a multidrug resistance transporter of the major facilitator superfamily, plays an essential role in spore wall maturation in Saccharomyces cerevisiae. Eukaryot. Cell 2002, 1, 799–810. [Google Scholar] [CrossRef] [Green Version]
- Drake, E.J.; Gulick, A.M. Three-dimensional structures of Pseudomonas aeruginosa PvcA and PvcB, two proteins involved in the synthesis of 2-isocyano-6,7-dihydroxycoumarin. J. Mol. Biol. 2008, 384, 193–205. [Google Scholar] [CrossRef] [Green Version]
- Clarke-Pearson, M.F.; Brady, S.F. Paerucumarin, a new metabolite produced by the pvc gene cluster from Pseudomonas aeruginosa. J. Bacteriol. 2008, 190, 6927–6930. [Google Scholar] [CrossRef] [Green Version]
- Longerich, I.; Taraz, K.; Budzikiewicz, H.; Tsai, L.; Meyer, J.M. Pseudoverdin, a compound related to the pyoverdin chromophore from a Pseudomonas aeruginosa strain incapable to produce pyoverdins. Z. Naturforsch. C J. Biosci. 1993, 48, 425–429. [Google Scholar] [CrossRef]
- Bemena, L.D.; Mukama, O.; Wang, N.; Gao, X.D.; Nakanishi, H. Characterization of a yeast sporulation-specific P450 family protein, Dit2, using an in vitro assay to crosslink formyl tyrosine. J. Biochem. 2018, 163, 123–131. [Google Scholar] [CrossRef]
- Sikorski, R.S.; Hieter, P. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae. Genetics 1989, 122, 19–27. [Google Scholar]
- Neiman, A.M.; Katz, L.; Brennwald, P.J. Identification of domains required for developmentally regulated SNARE function in Saccharomyces cerevisiae. Genetics 2000, 155, 1643–1655. [Google Scholar]
- Zhang, H.; Tachikawa, H.; Gao, X.D.; Nakanishi, H. Applied usage of yeast spores as chitosan beads. Appl. Environ. Microbiol. 2014, 80, 5098–5105. [Google Scholar] [CrossRef] [Green Version]
- Kremers, G.J.; Goedhart, J.; van den Heuvel, D.J.; Gerritsen, H.C.; Gadella, T.W., Jr. Improved green and blue fluorescent proteins for expression in bacteria and mammalian cells. Biochemistry 2007, 46, 3775–3783. [Google Scholar] [CrossRef]
- Mohamed, L.A.; Tachikawa, H.; Gao, X.D.; Nakanishi, H. Yeast cell-based analysis of human lactate dehydrogenase isoforms. J. Biochem. 2015, 158, 467–476. [Google Scholar] [CrossRef]
- Mumberg, D.; Muller, R.; Funk, M. Yeast vectors for the controlled expression of heterologous proteins in different genetic backgrounds. Gene 1995, 156, 119–122. [Google Scholar] [CrossRef]
- Pan, H.P.; Wang, N.; Tachikawa, H.; Nakanishi, H.; Gao, X.D. Beta-1,6-glucan synthesis-associated genes are required for proper spore wall formation in Saccharomyces cerevisiae. Yeast 2017, 34, 431–446. [Google Scholar] [CrossRef] [Green Version]
- Amado, R.; Aeschbach, R.; Neukom, H. Dityrosine: In vitro production and characterization. Methods Enzymol. 1984, 107, 377–388. [Google Scholar] [CrossRef]
- Pan, H.P.; Wang, N.; Tachikawa, H.; Gao, X.D.; Nakanishi, H. Osw2 is required for proper assembly of glucan and/or mannan layers of the yeast spore wall. J. Biochem. 2018, 163, 293–304. [Google Scholar] [CrossRef]
- Lim, Y.Y.; Stein, A.R. Acid-catalyzed Solvolysis of Isonitriles. I. Can. J. Chem. 1971, 49, 2455–2459. [Google Scholar] [CrossRef]
- Briza, P.; Ellinger, A.; Winkler, G.; Breitenbach, M. Characterization of a DL-dityrosine-containing macromolecule from yeast ascospore walls. J. Biol. Chem. 1990, 265, 15118–15123. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Source |
|---|---|---|
| AN120 | MATα/MATa ARG4/arg4-NspI his3∆SK/his3∆SKho::LYS2/ho::LYS2 leu2/leu2 lys2/lys2 RME1/rme1::LEU2trp1::hisG/trp1::hisG ura3/ura3 | [13] |
| AN117-4B | MAT α ura3 leu2 trp1 his3∆SK arg4-NspI lys2 ho::LYS2 rme1::LEU2 | [13] |
| AN117-16D | MATa ura3 leu2 trp1 his3∆SK lys2 ho::LYS2 | [13] |
| YPH499 | MATa ura3-52 lys2-801_amber ade2-101_ochre trp1-Δ63 his3-Δ200 leu2-Δ1 | [12] |
| HW3 (dit1∆) | MATα/MATa ARG4/arg4-NspI his3∆SK/his3∆SK ho::LYS2/ho::LYS2 leu2/leu2 lys2/lys2RME1/rme1::LEU2 trp1::hisG/trp1::hisG ura3/ura3 dit1∆::his5+/dit1∆::his5+ | [14] |
| HI391 | MATα/MATa ARG4/arg4-NspI his3∆SK/his3∆SKho::LYS2/ho::LYS2 leu2/leu2 lys2/lys2 RME1/rme1::LEU2trp1::hisG/trp1::hisG ura3/ura3 DIT1::GFP-HIS3 | this study |
| Name | Sequence |
|---|---|
| HP128 | GTGTAAGCTTATGGTGAGCAAGGGCGAGG |
| HP129 | GTGTCTCGAGTTACTTGTACAGCTCGTCCA |
| HXO683 | GTGTACTAGTAAAATGACATTTACTAGCAA |
| ECOR1-DIT1 -r | GTGTGAATTCAGAGATTTTCTTGATAACGA |
| HXO685 | GTGTACTAGTCAAATGGAGTTGTTAAAGCT |
| DIT2-FLAG-r | CCGGGGGATCCTTACTTATCGTCGTCATCCTTGTAATCTCCTGCTCCTGCTTCCATTATATTCTCGTTAA |
| DIT1-GFP-F-INT | CTCGTTTCGATATTGGAGAAGGAGGACATTTCGTTATCAAGAAAATCTCTCGGATCCCCGGGTTAATTAA |
| DIT1-GFP-R-INT | TGTTTAAGTAAAAGAACAAAAAGGTAGACCAATGTAGCGCTCTTACTTTAGAATTCGAGCTCGTTTAAAC |
| DIT1-HA-r | GTGTGAATTCTTATGCATAATCCGGAACATCATACGGATATCCTGCTCCTGCAGAGATTTTCTTGATAACGA |
| Name | Description | Source |
|---|---|---|
| pRS316TEF-PvcA | URA3; for expression of PvcA from The constitutive TEF2 promoter | this study |
| pRS424GAL1pr-DIT1-GFP | TRP1; for expression of Dit1-GFP from the GAL1 promoter | this study |
| pRS426GAL1pr-DIT2-FLAG | URA3; for expression of Dit2-FLAG from the GAL1 promoter | this study |
| pRS424GAL1pr-DIT1-HA | TRP1; for expression of Dit1-HA from the GAL1 promoter | this study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basiony, M.; Yang, Y.; Liu, G.; Gao, X.-D.; Nakanishi, H. Studies on the Properties of the Sporulation Specific Protein Dit1 and Its Product Formyl Tyrosine. J. Fungi 2020, 6, 77. https://doi.org/10.3390/jof6020077
Basiony M, Yang Y, Liu G, Gao X-D, Nakanishi H. Studies on the Properties of the Sporulation Specific Protein Dit1 and Its Product Formyl Tyrosine. Journal of Fungi. 2020; 6(2):77. https://doi.org/10.3390/jof6020077
Chicago/Turabian StyleBasiony, Mostafa, Yan Yang, Guoyu Liu, Xiao-Dong Gao, and Hideki Nakanishi. 2020. "Studies on the Properties of the Sporulation Specific Protein Dit1 and Its Product Formyl Tyrosine" Journal of Fungi 6, no. 2: 77. https://doi.org/10.3390/jof6020077
APA StyleBasiony, M., Yang, Y., Liu, G., Gao, X. -D., & Nakanishi, H. (2020). Studies on the Properties of the Sporulation Specific Protein Dit1 and Its Product Formyl Tyrosine. Journal of Fungi, 6(2), 77. https://doi.org/10.3390/jof6020077