Saccharomyces boulardii: What Makes It Tick as Successful Probiotic?



Abstract
:1. Introduction
2. S. boulardii and S. cerevisiae: Similar but Different
3. S. boulardii Genomic Variations Provide Hints for Its Physiological Properties
4. Adaptation to Host Environment
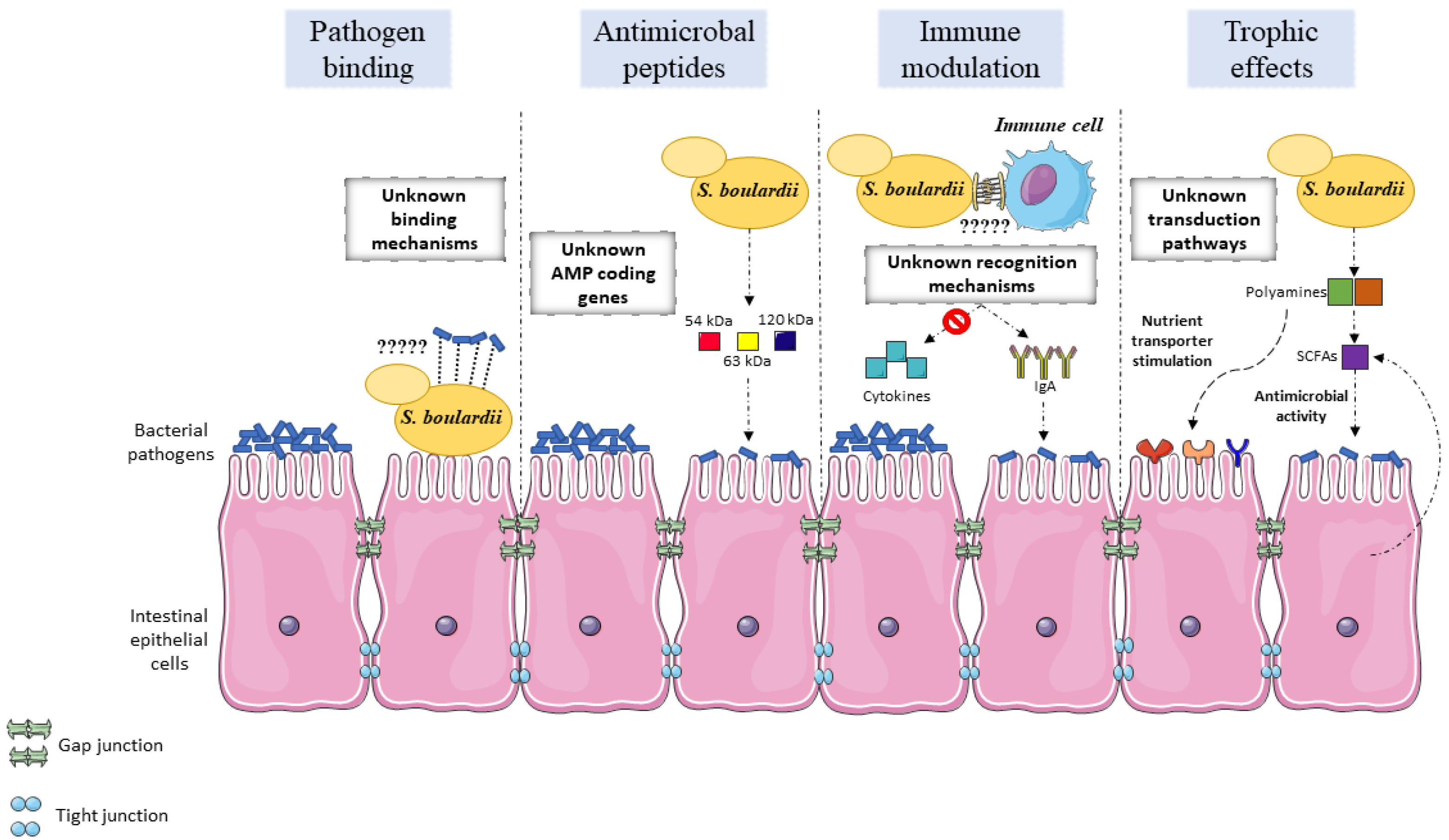
5. Mechanisms of Action
5.1. Modulation of The Normal Microbiome
5.2. Antimicrobial Activity
5.3. Adhesion
5.4. Immune Modulation
5.5. Trophic Effects
6. S. boulardii Safety and Clinical Efficacy
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- McFarland, L.V. Common Organisms and Probiotics: Saccharomyces boulardii. In The Microbiota in Gastrointestinal Pathophysiology; Academic Press: Cambridge, MA, USA, 2017; pp. 145–164. [Google Scholar]
- Schrezenmeir, J.; de Vrese, M. Probiotics, prebiotics, and synbiotics—approaching a definition. Am. J. Clin. Nutr. 2001, 73, 361s–364s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moradi, R.; Nosrati, R.; Zare, H.; Tahmasebi, T.; Saderi, H.; Owlia, P. Screening and characterization of in-vitro probiotic criteria of saccharomyces and kluyveromyces strains. Iran. J. Microbiol. 2018, 10, 123–131. [Google Scholar] [PubMed]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of Action of Probiotics. Adv. Nutr. 2019, 10, S49–S66. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Rajput, I.R.; Arain, M.A.; Li, Y.; Baloch, D.M. Oral administration of Saccharomyces boulardii alters duodenal morphology, enzymatic activity and cytokine production response in broiler chickens. Anim. Sci. J. 2017, 88, 1204–1211. [Google Scholar] [CrossRef] [PubMed]
- Kabluchko, T.V.; Bomko, T.V.; Nosalskaya, T.N.; Martynov, A.V.; Osolodchenko, T.P. In the gastrointestinal tract exist the protective mechanisms which prevent overgrowth of pathogenic bacterial and its incorporation. Ann. Mechnikov Inst. 2017, 1, 28–33. [Google Scholar]
- Kelesidis, T.; Pothoulakis, C. Efficacy and safety of the probiotic Saccharomyces boulardii for the prevention and therapy of gastrointestinal disorders. Therap. Adv. Gastroenterol. 2012, 5, 111–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, S.; Mansell, T.J. Yeasts as probiotics: Mechanisms, outcomes, and future potential. Fungal Genet. Biol. 2020, 137, 103333. [Google Scholar] [CrossRef]
- Van Der Aa Kühle, A.; Jespersen, L. The Taxonomic Position of Saccharomyces boulardii as Evaluated by Sequence Analysis of the D1/D2 Domain of 26S rDNA, the ITS1-5.8S rDNA-ITS2 Region and the Mitochondrial Cytochrome-c Oxidase II Gene. Syst. Appl. Microbiol. 2003, 26, 564–571. [Google Scholar] [CrossRef]
- Mitterdorfer, G.; Mayer, H.K.; Kneifel, W.; Viernstein, H. Clustering of Saccharomyces boulardii strains within the species S. cerevisiae using molecular typing techniques. J. Appl. Microbiol. 2002, 93, 521–530. [Google Scholar] [CrossRef]
- Khatri, I.; Tomar, R.; Ganesan, K.; Prasad, G.S.; Subramanian, S. Complete genome sequence and comparative genomics of the probiotic yeast Saccharomyces boulardii. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Fietto, J.L.; Araújo, R.S.; Valadão, F.N.; Fietto, L.G.; Brandão, R.L.; Neves, M.J.; Gomes, F.C.; Nicoli, J.R.; Castro, I.M. Molecular and physiological comparisons between Saccharomyces cerevisiae and Saccharomyces boulardii. Can. J. Microbiol. 2004, 50, 615–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards-ingram, L.C.; Gent, M.E.; Hoyle, D.C.; Hayes, A.; Stateva, L.I.; Oliver, S.G. Comparative Genomic Hybridization Provides New Insights Into the Molecular Taxonomy of the Saccharomyces Sensu Stricto Complex. Genome Res. 2004, 14, 1043–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennequin, C.; Thierry, A.; Richard, G.F.; Lecointre, G.; Nguyen, H.V.; Gaillardin, C.; Dujon, B. Microsatellite Typing as a New Tool for Identification of Saccharomyces cerevisiae Strains. J. Clin. Microbiol. 2001, 39, 551–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFarland, L.V. Systematic review and meta-analysis of saccharomyces boulardii in adult patients. World J. Gastroenterol. 2010, 16, 2202–2222. [Google Scholar] [CrossRef]
- McFarland, L.V. Saccharomyces boulardii Is Not Saccharomyces cerevisiae. Clin. Infect. Dis. 1996, 22, 200–201. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.J.; Zhang, G.C.; Kong, I.I.; Yun, E.J.; Zheng, J.Q.; Kweon, D.H.; Jin, Y.S. A mutation in PGM2 causing inefficient galactose metabolism in the probiotic yeast Saccharomyces boulardii. Appl. Environ. Microbiol. 2018, 84, e02858-17. [Google Scholar] [CrossRef] [Green Version]
- Edwards-ingram, L.; Gitsham, P.; Burton, N.; Warhurst, G.; Clarke, I.; Hoyle, D.; Oliver, S.G.; Stateva, L. Genotypic and Physiological Characterization of Saccharomyces boulardii, the Probiotic Strain of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2007, 73, 2458–2467. [Google Scholar] [CrossRef] [Green Version]
- GMitterdorfer; WKneifel; HViernstein Utilization of prebiotic carbohydrates by yeasts of therapeutic relevance. Lett. Appl. Microbiol. 2001, 34, 251–255.
- Klein, S.M.; Elmer, G.W.; McFarland, L.V.; Surawicz, C.M.; Levy, R.H. Recovery and Elimination of the Biotherapeutic Agent, Saccharomyces boulardii, in Healthy Human Volunteers. Pharm. Res. An Off. J. Am. Assoc. Pharm. Sci. 1993, 10, 1615–1619. [Google Scholar]
- Rodrigues, A.C.P.; Mardi, R.M.; Bambirra, E.A.; Vieira, E.G.; Nicoli, U.R. Effect of Saccharomyces boulardii against experimental oral infection with Salmonella typhimurium and Shigella flexnerim conventional and gnotobiotic mice. J. Appl. Bacteriol. 1996, 81, 251–256. [Google Scholar] [CrossRef]
- Offei, B.; Vandecruys, P.; De Graeve, S.; Foulquié-moreno, M.R.; Thevelein, J.M. Unique genetic basis of the distinct antibiotic potency of high acetic acid production in the probiotic yeast Saccharomyces cerevisiae var. boulardii. Genome Res. 2019, 1478–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Styles, C.A.; Fink, G.R. Saccharomyces cerevisiae S288C has a mutation in FLO8, a gene required for filamentous growth. Genetics 1996, 144, 967–978. [Google Scholar] [PubMed]
- van den Brink, J.; Akeroyd, M.; van der Hoeven, R.; Ponk, J.T.; de Winde, J.H.; Daran-Lapujade, P.A.S. Energetic limits to metabolic flexibility: Responses of Saccharomyces cerevisiae to glucose-galactose transitions. Microbiology 2009, 155, 1340–1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatri, I.; Akhtar, A.; Kaur, K.; Tomar, R.; Prasad, G.S. Gleaning evolutionary insights from the genome sequence of a probiotic yeast Saccharomyces boulardii. Gut Pathog. 2013, 5, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinsen, T.C.; Bergh, K.; Waldum, H.L. Gastric Juice: A Barrier Against Infectious Diseases. Basic Clin. Pharmacol. Toxicol. 2005, 96, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Holzapfel, W.H.; Haberer, P.; Snel, J.; Schillinger, U.; Huis In’T Veld, J.H.J. Overview of gut flora and probiotics. Int. J. Food Microbiol. 1998, 41, 85–101. [Google Scholar] [CrossRef]
- Du Le, H.; Trinh, K.S. Survivability of Lactobacillus acidophilus, Bacillus clausii and Saccharomyces boulardii encapsulated in alginate gel microbeads. Carpathian J. Food Sci. Technol. 2018, 10, 95–103. [Google Scholar]
- Cordonnier, C.; Thévenot, J.; Etienne-Mesmin, L.; Denis, S.; Alric, M.; Livrelli, V.; Blanquet-Diot, S. Dynamic In Vitro Models of the Human Gastrointestinal Tract as Relevant Tools to Assess the Survival of Probiotic Strains and Their Interactions with Gut Microbiota. Microorganisms 2015, 3, 725–745. [Google Scholar] [CrossRef]
- Filho-Lima, J.V.M.; Vieira, E.C.; Nicoli, J.R. Antagonistic effect of Lactobacillus acidophilus, Saccharomyces boulardii and Escherichia coli combinations against experimental infections with Shigella flexneri and Salmonella enteritidis subsp. typhimurium in gnotobiotic mice. J. Appl. Microbiol. 2000, 88, 365–370. [Google Scholar] [CrossRef] [Green Version]
- Bajaj, B.K.; Claes, I.J.J.; Lebeer, S. Functional mechanisms of probiotics. J. Microbiol. Biotechnol. Food Sci. 2015, 4, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Ceapa, C.; Wopereis, H.; Rezaïki, L.; Kleerebezem, M.; Knol, J.; Oozeer, R. Influence of fermented milk products, prebiotics and probiotics on microbiota composition and health. Best Pract. Res. Clin. Gastroenterol. 2013, 27, 139–155. [Google Scholar] [CrossRef] [PubMed]
- Dethlefsen, L.; Huse, S.; Sogin, M.L.; Relman, D.A. The Pervasive Effects of an Antibiotic on the Human Gut Microbiota, as Revealed by Deep 16S rRNA Sequencing. PLoS Biol. 2008, 6, e280. [Google Scholar] [CrossRef]
- McFarland, L.V. Use of probiotics to correct dysbiosis of normal microbiota following disease or disruptive events: A systematic review. BMJ Open 2014, 4, e005047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic Mechanisms of Action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Ducluzeau, R.; Bensaada, M. Comparative effect of a single or continuous administration of “Saccharomyces boulardii” on the establishment of various strains of “candida” in the digestive tract of gnotobiotic mice. Ann. Microbiol. (Paris) 1982, 133, 491–501. [Google Scholar] [PubMed]
- de Arauz, L.J.; Jozala, A.F.; Mazzola, P.G.; Vessoni Penna, T.C. Nisin biotechnological production and application: A review. Trends Food Sci. Technol. 2009, 20, 146–154. [Google Scholar] [CrossRef]
- Vilagravel, B.; Esteve-Garcia, E.; Brufau, J. Probiotic micro-organisms: 100 years of innovation and efficacy; Modes of action. Worlds. Poult. Sci. J. 2010, 66, 369–380. [Google Scholar]
- Corr, S.C.; Li, Y.; Riedel, C.U.; O’Toole, P.W.; Hill, C.; Gahan, C.G.M. Bacteriocin production as a mechanism for the antiinfective activity of Lactobacillus salivarius UCC118. Proc. Natl. Acad. Sci. USA. 2007, 104, 7617–7621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, D.S.; Cho, G.S.; Hanak, A.; Huch, M.; Franz, C.M.A.P.; Arneborg, N. The effect of bacteriocin-producing Lactobacillus plantarum strains on the intracellular pH of sessile and planktonic Listeria monocytogenes single cells. Int. J. Food Microbiol. 2010, 141, S53–S59. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Vallance, B.A.; Boyer, L.; Bergstrom, K.S.B.; Walker, J.; Madsen, K.; O’Kusky, J.R.; Buchan, A.M.; Jacobson, K. Saccharomyces boulardii ameliorates Citrobacter rodentium-induced colitis through actions on bacterial virulence factors. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 294, G295–G306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castagliuolo, I.; Riegler, M.F.; Valenick, L.; LaMont, J.T.; Pothoulakis, C. Saccharomyces boulardii protease inhibits the effects of Clostridium difficile toxins A and B in human colonic mucosa. Infect. Immun. 1999, 67, 302–307. [Google Scholar] [CrossRef] [Green Version]
- Castagliuolo, I.; Thomas Lamont, J.; Nikulasson, S.T.; Pothoulakis, C. Saccharomyces boulardii protease inhibits Clostridium difficile toxin A effects in the rat ileum. Infect. Immun. 1996, 64, 5225–5232. [Google Scholar] [CrossRef] [Green Version]
- Buts, J.P.; Bernasconi, P.; Van Craynest, M.P.; Maldague, P.; Meyer, R. Response of human and rat small intestinal mucosa to oral administration of saccharomyces boulardii. Pediatr. Res. 1986, 20, 192–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buts, J.P.; Bernasconi, P.; Vaerman, J.P.; Dive, C. Stimulation of secretory IgA and secretory component of immunoglobulins in small intestine of rats treated with Saccharomyces boulardii. Dig. Dis. Sci. 1990, 35, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Buts, J.P.; Dekeyser, N.; Stilmant, C.; Delem, E.; Smets, F.; Sokal, E. Saccharomyces boulardii produces in rat small intestine a novel protein phosphatase that inhibits Escherichia coli endotoxin by dephosphorylation. Pediatr. Res. 2006, 60, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Czerucka, D.; Roux, I.; Rampal, P. Saccharomyces boulardii inhibits secretagogue-mediated adenosine 3′,5′-cyclic monophosphate induction in intestinal cells. Gastroenterology 1994, 106, 65–72. [Google Scholar] [CrossRef]
- Brandão, R.L.; Castro, I.M.; Bambirra, E.A.; Amaral, S.C.; Fietto, L.G.; Tropia, M.J.M.; Neves, M.J.; Dos Santos, R.G.; Gomes, N.C.M.; Nicoli, J.R. Intracellular signal triggered by cholera toxin in Saccharomyces boulardii and Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1998, 64, 564–568. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, D.A.; Hicks, C.W.; Cui, X.; Li, Y.; Eichacker, P.Q. Anthrax infection. Am. J. Respir. Crit. Care Med. 2011, 184, 1333–1341. [Google Scholar] [CrossRef]
- Pontier-bres, R.; Rampal, P.; Peyron, J.; Munro, P.; Lemichez, E.; Czerucka, D. The Saccharomyces boulardii CNCM I-745 Strain Shows Protective Effects against the B. anthracis LT Toxin. Toxin 2015, 7, 4455–4467. [Google Scholar] [CrossRef] [Green Version]
- Albenberg, L.; Esipova, T.V.; Judge, C.P.; Bittinger, K.; Chen, J.; Laughlin, A.; Grunberg, S.; Baldassano, R.N.; Lewis, J.D.; Li, H.; et al. Correlation between intraluminal oxygen gradient and radial partitioning of intestinal microbiota. Gastroenterology 2014, 147, 1055–1063. [Google Scholar] [CrossRef] [Green Version]
- Vacca, I. Microbiome: The microbiota maintains oxygen balance in the gut. Nat. Rev. Microbiol. 2017, 15, 574. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Chávez, F.; Lopez, C.A.; Bäumler, A.J. Oxygen as a driver of gut dysbiosis. Free Radic. Biol. Med. 2017, 105, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Gogineni, V.K.; Morrow, L.E. Probiotics: Mechanisms of Action and Clinical Applications. J. Probiotics Heal. 2013, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Tiago, F.C.P.; Martins, F.S.; Souza, E.L.S.; Pimenta, P.F.P.; Araujo, H.R.C.; Castro, I.M.; Branda, R.L.; Nicoli, J.R.; Nicoli, J.R. Adhesion to the yeast cell surface as a mechanism for trapping pathogenic bacteria by Saccharomyces probiotics. J. Med Microbiol. 2012, 61, 1194–1207. [Google Scholar] [CrossRef] [PubMed]
- Gedek, B.R. Adherence of Escherichia coli serogroup 0 157 and the Salmonella Typhimurium mutant DT 104 to the surface of Saccharomyces boulardii. Mycoses 1999, 42, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Pontier-bres, R.; Munro, P.; Boyer, L.; Anty, R.; Rampal, P.; Lemichez, E. Saccharomyces boulardii Modifies Salmonella Typhimurium Traffic and Host Immune Responses along the Intestinal Tract. PLoS ONE 2014, 9, e103069. [Google Scholar] [CrossRef] [PubMed]
- Martins, F.S.; Dalmasso, G.; Arantes, R.M.E.; Doye, A.; Lemichez, E.; Lagadec, P.; Imbert, V.; Peyron, J.F.; Rampal, P.; Nicoli, J.R.; et al. Interaction of Saccharomyces boulardii with Salmonella enterica serovar typhimurium protects mice and modifies T84 cell response to the infection. PLoS ONE 2010, 5, e8925. [Google Scholar] [CrossRef] [Green Version]
- Tasteyre, A.; Barc, M.C.; Karjalainen, T.; Bourlioux, P.; Collignon, A. Inhibition of in vitro cell adherence of Clostridium difficile by Saccharomyces boulardii. Microb. Pathog. 2002, 32, 219–225. [Google Scholar] [CrossRef]
- Rigothier, M.C.; Maccario, J.; Gayral, P. Inhibitory activity of saccharomyces yeasts on the adhesion of Entamoeba histolytica trophozoites to human erythrocytes in vitro. Parasitol. Res. 1994, 80, 10–15. [Google Scholar] [CrossRef]
- Pontier-Bres, R.; Prodon, F.; Munro, P.; Rampal, P.; Lemichez, E.; Peyron, J.F.; Czerucka, D. Modification of salmonella typhimurium motility by the probiotic yeast strain saccharomyces boulardii. PLoS ONE 2012, 7, e33796. [Google Scholar] [CrossRef]
- Kline, K.A.; Fälker, S.; Dahlberg, S.; Normark, S.; Henriques-Normark, B. Bacterial Adhesins in Host-Microbe Interactions. Cell Host Microbe 2009, 5, 580–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cusumano, C.K.; Hultgren, S.J. Bacterial adhesion—A source of alternate antibiotic targets. IDrugs 2009, 12, 699–705. [Google Scholar] [PubMed]
- Gómez Zavaglia, A.; Kociubinski, G.; Pérez, P.; Disalvo, E.; De Antoni, G. Effect of bile on the lipid composition and surface properties of bifidobacteria. J. Appl. Microbiol. 2002, 93, 794–799. [Google Scholar] [CrossRef] [Green Version]
- Kaur, I.P.; Kuhad, A.; Garg, A.; Chopra, K. Probiotics: Delineation of Prophylactic and Therapeutic Benefits. J. Med. Food 2009, 12, 219–235. [Google Scholar] [CrossRef]
- Chen, K.-H.; Miyazaki, T.; Tsai, H.-F.; Bennett, J.E. The bZip transcription factor Cgap1p is involved in multidrug resistance and required for activation of multidrug transporter gene CgFLR1 in Candida glabrata. Gene 2007, 386, 63–72. [Google Scholar] [CrossRef]
- Fidan, I.; Kalkanci, A.; Yesilyurt, E.; Yalcin, B.; Erdal, B.; Kustimur, S.; Imir, T. Effects of Saccharomyces boulardii on cytokine secretion from intraepithelial lymphocytes infected by Escherichia coli and Candida albicans. Mycoses 2009, 52, 29–34. [Google Scholar] [CrossRef]
- Qamar, A.; Aboudola, S.; Warny, M.; Michetti, P.; Kelly, N.P.; Division, G.; Israel, B.; Medical, D. Saccharomyces boulardii Stimulates Intestinal Immunoglobulin A Immune Response to Clostridium difficile Toxin A in Mice. Infect. Immun. 2001, 69, 2762–2765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyne, L.; Warny, M.; Qamar, A.; Kelly, C.P. Association between antibody response to toxin A and protection against recurrent Clostridium difficile diarrhoea. Lancet 2001, 357, 189–193. [Google Scholar] [CrossRef]
- Rajput, I.R.; Hussain, A.; Li, Y.L.; Zhang, X.; Xu, X.; Long, M.Y.; You, D.Y.; Li, W.F. Saccharomyces boulardii and Bacillus subtilis B10 Modulate TLRs Mediated Signaling to Induce Immunity by Chicken BMDCs. J. Cell. Biochem. 2014, 115, 189–198. [Google Scholar] [CrossRef]
- Badia, R.; Zanello, G.; Chevaleyre, C.; Lizardo, R.; Meurens, F.; Martínez, P.; Brufau, J.; Salmon, H. Effect of Saccharomyces cerevisiae var. Boulardii and β-galactomannan oligosaccharide on porcine intestinal epithelial and dendritic cells challenged in vitro with Escherichia coli F4 (K88). Vet. Res. 2012, 43, 4. [Google Scholar] [CrossRef] [Green Version]
- Badia, R.; Brufau, M.T.; Guerrero-Zamora, A.M.; Lizardo, R.; Dobrescu, I.; Martin-Venegas, R.; Ferrer, R.; Salmon, H.; Martínez, P.; Brufau, J. β-galactomannan and Saccharomyces cerevisiae var. boulardii modulate the immune response against Salmonella enterica serovar typhimurium in porcine intestinal epithelial and dendritic cells. Clin. Vaccine Immunol. 2012, 19, 368–376. [Google Scholar] [CrossRef]
- Dalmasso, G.; Cottrez, F.; Imbert, V.; Lagadec, P.; Peyron, J.F.; Rampal, P.; Czerucka, D.; Groux, H. Saccharomyces boulardii Inhibits Inflammatory Bowel Disease by Trapping T Cells in Mesenteric Lymph Nodes. Gastroenterology 2006, 131, 1812–1825. [Google Scholar] [CrossRef] [PubMed]
- Moré, M.I.; Vandenplas, Y. Saccharomyces boulardii CNCM I-745 Improves Intestinal Enzyme Function: A Trophic Effects Review. Clin. Med. Insights Gastroenterol. 2018, 11, 1179552217752679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buts, J.P.; De Keyser, N.; Raedemaeker, L. De Saccharomyces boulardii enhances rat intestinal enzyme expression by endoluminal release of polyamines. Pediatr. Res. 1994, 36, 522–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaouche, A.; Loukil, C.; De Lagausie, P.; Peuchmaur, M.; Macry, J.; Fitoussi, F.; Bernasconi, P.; Bingen, E.; Cezard, J.P. Effects of oral Saccharomyces boulardii on bacterial overgrowth, translocation, and intestinal adaptation after small-bowel resection in rats. Scand. J. Gastroenterol. 2000, 35, 160–165. [Google Scholar] [CrossRef]
- Buts, J.P.; De Keyser, N.; Marandi, S.; Hermans, D.; Sokal, E.M.; Chae, Y.H.E.; Lambotte, L.; Chanteux, H.; Tulkens, P.M. Saccharomyces boulardii upgrades cellular adaptation after proximal enterectomy in rats. Gut 1999, 45, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Harms, H.-K.; Bertele-Harms, R.-M.; Bruer-Kleis, D. Enzyme-Substitution Therapy with the Yeast Saccharomyces cerevisiae in Congenital Sucrase-Isomaltase Deficiency. N. Engl. J. Med. 1987, 316, 1306–1309. [Google Scholar] [CrossRef]
- Buts, J.-P.; De Keyser, N.; Stilmant, C.; Sokal, E.; Marandi, S. Saccharomyces boulardii Enhances N-Terminal Peptide Hydrolysis in Suckling Rat Small Intestine by Endoluminal Release of a Zinc-Binding Metalloprotease. Pediatr. Res. 2002, 51, 528–534. [Google Scholar] [CrossRef] [Green Version]
- Jahn, H.U.; Ullrich, R.; Schneider, T.; Liehr, R.M.; Schieferdecker, H.L.; Holst, H.; Zeitz, M. Immunological and trophical effects of saccharomyces boulardii on the small intestine in healthy human volunteers. Digestion 1996, 57, 95–104. [Google Scholar] [CrossRef]
- Buts, J.P.; Keyser, N. De Transduction pathways regulating the trophic effects of Saccharomyces boulardii in rat intestinal mucosa. Scand. J. Gastroenterol. 2010, 45, 175–185. [Google Scholar] [CrossRef]
- Buts, J.P.; Dekeyser, N. Raf: A key regulatory kinase for transduction of mitogenic and metabolic signals of the probiotic Saccharomyces boulardii. Clin. Res. Hepatol. Gastroenterol. 2011, 35, 596–597. [Google Scholar] [CrossRef]
- Chang, C.; Wang, K.; Zhou, S.N.; Wang, X.D.; Wu, J.E. Protective Effect of Saccharomyces boulardii on Deoxynivalenol-Induced Injury of Porcine Macrophage via Attenuating p38 MAPK Signal Pathway. Appl. Biochem. Biotechnol. 2017, 182, 411–427. [Google Scholar] [CrossRef]
- Schneider, S.M.; Girard-Pipau, F.; Filippi, J.; Hébuterne, X.; Moyse, D.; Hinojosa, G.C.; Pompei, A.; Rampal, P. Effects of Saccharomyces boulardii on fecal short-chain fatty acids and microflora in patients on long-term total enteral nutrition. World J. Gastroenterol. 2005, 11, 6165–6169. [Google Scholar] [CrossRef]
- Girard-Pipau, F.; Pompei, A.; Schneider, S.; Nano, J.L.; Hebuterne, X.; Boquet, P.; Rampal, P. Intestinal microflora, short chain and cellular fatty acids, influence of a probiotic Saccharomyces boulardii. Microb. Ecol. Health Dis. 2002, 14, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Macfarlane, G.T.; Macfarlane, S. Bacteria, colonic fermentation, and gastrointestinal health. J. AOAC Int. 2012, 95, 50–60. [Google Scholar] [CrossRef]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T.; et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2011, 469, 543–549. [Google Scholar] [CrossRef]
- Wannaprasat, W.; Koowatananukul, C.; Ekkapobyotin, C.; Chuanchuen, R. Quality analysis of commercial probiotic products for food animals. Southeast Asian J. Trop. Med. Public Health 2009, 40, 1103–1112. [Google Scholar]
- Salminen, M.K.; Rautelin, H.; Tynkkynen, S.; Poussa, T.; Saxelin, M.; Valtonen, V.; Jarvinen, A. Lactobacillus Bacteremia, Species Identification, and Antimicrobial Susceptibility of 85 Blood Isolates. Clin. Infect. Dis. 2006, 42, e35–e44. [Google Scholar] [CrossRef] [PubMed]
- Karen, M.; Yuksel, O.; Akyürek, N.; Ofluoǧlu, E.; Çaǧlar, K.; Şahin, T.T.; Paşaoǧlu, H.; Memiş, L.; Akyürek, N.; Bostanci, H. Probiotic Agent Saccharomyces boulardii Reduces the Incidenceof Lung Injury in Acute Necrotizing Pancreatitis Induced Rats. J. Surg. Res. 2010, 160, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Lessard, M.; Dupuis, M.; Gagnon, N.; Nadeau, É.; Matte, J.J.; Goulet, J.; Fairbrother, J.M. Administration of Pediococcus acidilactici or Saccharomyces cerevisiae boulardii modulates development of porcine mucosal immunity and reduces intestinal bacterial translocation after Escherichia coli challenge. J. Anim. Sci. 2009, 87, 922–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byron, J.K.; Clemons, K.V.; McCusker, J.H.; Davis, R.W.; Stevens, D.A. Pathogenicity of Saccharomyces cerevisiae in complement factor five-deficient mice. Infect. Immun. 1995, 63, 478–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mcfarland, L.V.; Surawicz, C.M.; Elmer, G.W.; Moyer, K.A.; Melcher, S.A.; Fekety, R.; Bowen, K.E.; Cox, J.L.; Noorani, Z.; Harrington, G.; et al. A Randomized Placebo-Controlled Trial of Saccharomyces boulardii in Combination With Standard Antibiotics for Clostridium difficile Disease. JAMA J. Am. Med. Assoc. 1994, 271, 1913–1918. [Google Scholar] [CrossRef]
- Thygesen, J.B.; Glerup, H.; Tarp, B. Saccharomyces boulardii fungemia caused by treatment with a probioticum. BMJ Case Rep. 2012, 2012, bcr0620114412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appel-da-Silva, M.C.; Narvaez, G.A.; Perez, L.R.R.; Drehmer, L.; Lewgoy, J. Saccharomyces cerevisiae var. boulardii fungemia following probiotic treatment. Med. Mycol. Case Rep. 2017, 18, 15–17. [Google Scholar] [CrossRef] [PubMed]
- Cassone, M.; Serra, P.; Mondello, F.; Girolamo, A.; Scafetti, S.; Pistella, E.; Venditti, M. Outbreak of Saccharomyces cerevisiae Subtype boulardii Fungemia in Patients Neighboring Those Treated with a Probiotic Preparation of the Organism. J. Clin. Microbiol. 2003, 41, 5340–5343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enache-Angoulvant, A.; Hennequin, C. Invasive Saccharomyces Infection: A Comprehensive Review. Clin. Infect. Dis. 2005, 41, 1559–1568. [Google Scholar] [CrossRef]
- Dinleyici, E.C.; Kara, A.; Ozen, M.; Vandenplas, Y. Saccharomyces boulardii CNCM I-745 in different clinical conditions. Expert Opin. Biol. Ther. 2014, 14, 1593–1609. [Google Scholar] [CrossRef]
- Pineton de Chambrun, G.; Neut, C.; Chau, A.; Cazaubiel, M.; Pelerin, F.; Justen, P.; Desreumaux, P. A randomized clinical trial of Saccharomyces cerevisiae versus placebo in the irritable bowel syndrome. Dig. Liver Dis. 2015, 47, 119–124. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Features | S. Cerevisiae | S. Boulardii | |
|---|---|---|---|
| Optimal growth temperature [12] | 30 °C | 37 °C | |
| High temperature resistance (52 °C) [12] | 45% viability | 65% viability | |
| Acid pH resistance (pH = 2 for one hour) [12,18] | No—30% viability | Yes—75% viability | |
| Tolerance to bile acids (>0.3%(w/v)) [12] | No—Survival up to 0.15% (w/v) | No—Survival up to 0.10% (w/v) | |
| Basic pH resistance (pH = 8) [12,18] | Yes | Yes | |
| Assimilation of galactose [16,17,19] | Yes | No | |
| Ploidy [18] | Diploid or haploid | Always diploid | |
| Homo or heterothallic [11] | Homothallic | Homothallic | |
| Mating type [13] | Both | Both | |
| Sporulation [16,18] | Sporogenous | Asporogenous, but produces fertile hybrids with S. cerevisiae | |
| Pseudo-hyphal switching [18] | Normal | Increased | |
| Retrotransposon (Ty elements) [11] | Intact Ty elements | No intact Ty1, 3 or 4 elements | |
| Adhesion to epithelial cells | Normal microbiome (mice and human) [18,20] | No | No |
| Gnotobiotic mice [21] | Unknown | Yes | |
| Humans treated with ampicillin [20] | Unknown | Yes | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pais, P.; Almeida, V.; Yılmaz, M.; Teixeira, M.C. Saccharomyces boulardii: What Makes It Tick as Successful Probiotic? J. Fungi 2020, 6, 78. https://doi.org/10.3390/jof6020078
Pais P, Almeida V, Yılmaz M, Teixeira MC. Saccharomyces boulardii: What Makes It Tick as Successful Probiotic? Journal of Fungi. 2020; 6(2):78. https://doi.org/10.3390/jof6020078
Chicago/Turabian StylePais, Pedro, Vanda Almeida, Melike Yılmaz, and Miguel C. Teixeira. 2020. "Saccharomyces boulardii: What Makes It Tick as Successful Probiotic?" Journal of Fungi 6, no. 2: 78. https://doi.org/10.3390/jof6020078
APA StylePais, P., Almeida, V., Yılmaz, M., & Teixeira, M. C. (2020). Saccharomyces boulardii: What Makes It Tick as Successful Probiotic? Journal of Fungi, 6(2), 78. https://doi.org/10.3390/jof6020078