Alginate-Derived Elicitors Enhance β-Glucan Content and Antioxidant Activities in Culinary and Medicinal Mushroom, Sparassis latifolia



Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Marine Brown Algae
2.2. Extraction of Sodium Alginate
2.3. Preparation of Oligosaccharide Fractions
2.4. Physicochemical Analysis of Oligosaccharide Fractions
2.4.1. Molecular Weight Analysis
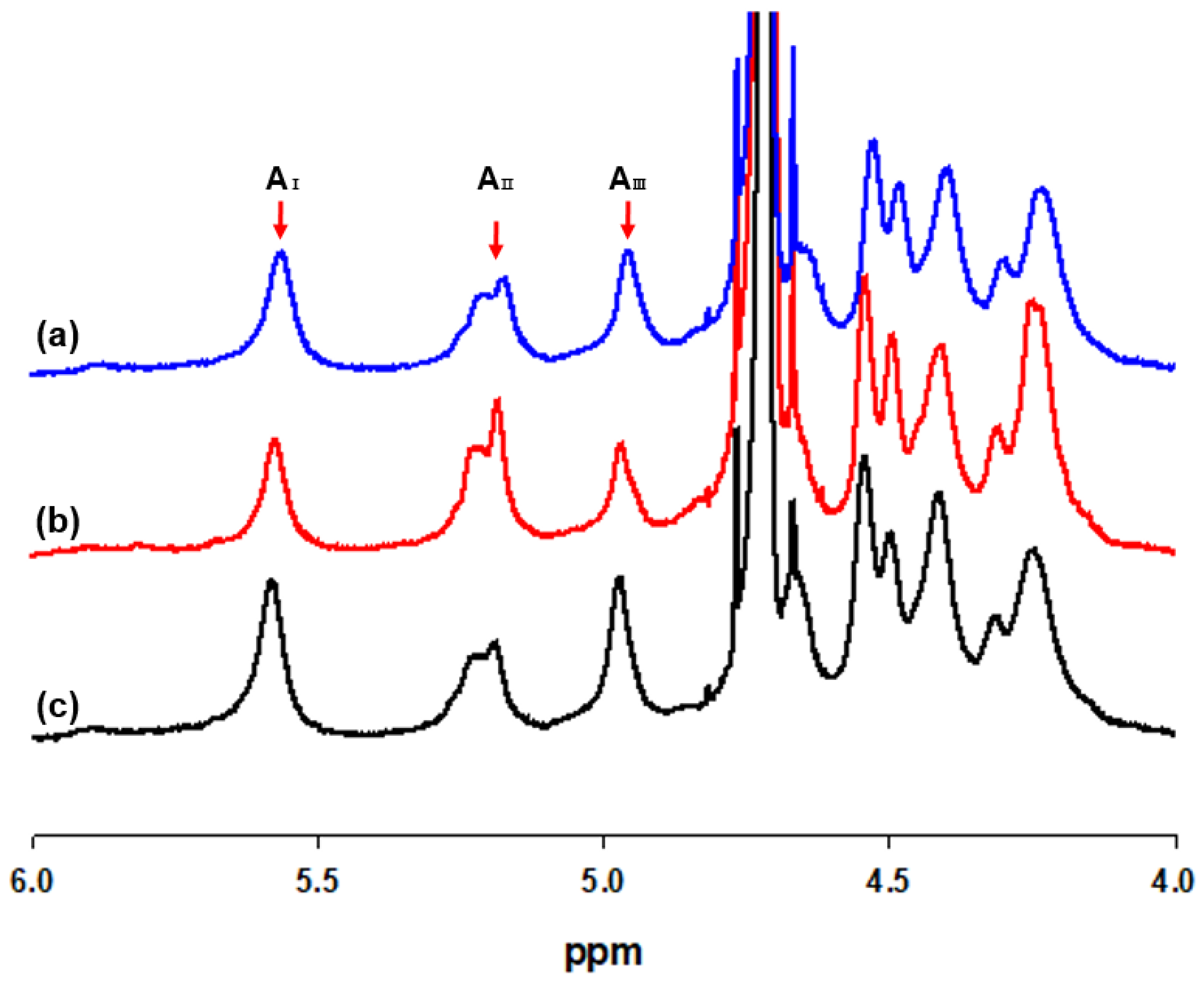
2.4.2. H NMR Spectroscopy Analysis
2.4.3. FT-IR Spectroscopy Analysis
2.4.4. Monosaccharide Analysis
2.5. Cultivation of Sparassis Latifolia
2.6. Elicitation by Alginate Oligosaccharide Fractions
2.7. Assay of Glucan Content
2.8. Determination of Total Polyphenol Content
2.9. Measurement of Antioxidant Activities
2.9.1. Assay of ABTS Radical Scavenging Activity
2.9.2. Assay of SOD-Like Activity
2.10. Statistical Analysis
3. Results
3.1. Physicochemical Properties of Alginate
3.2. Oligosaccharide Analysis
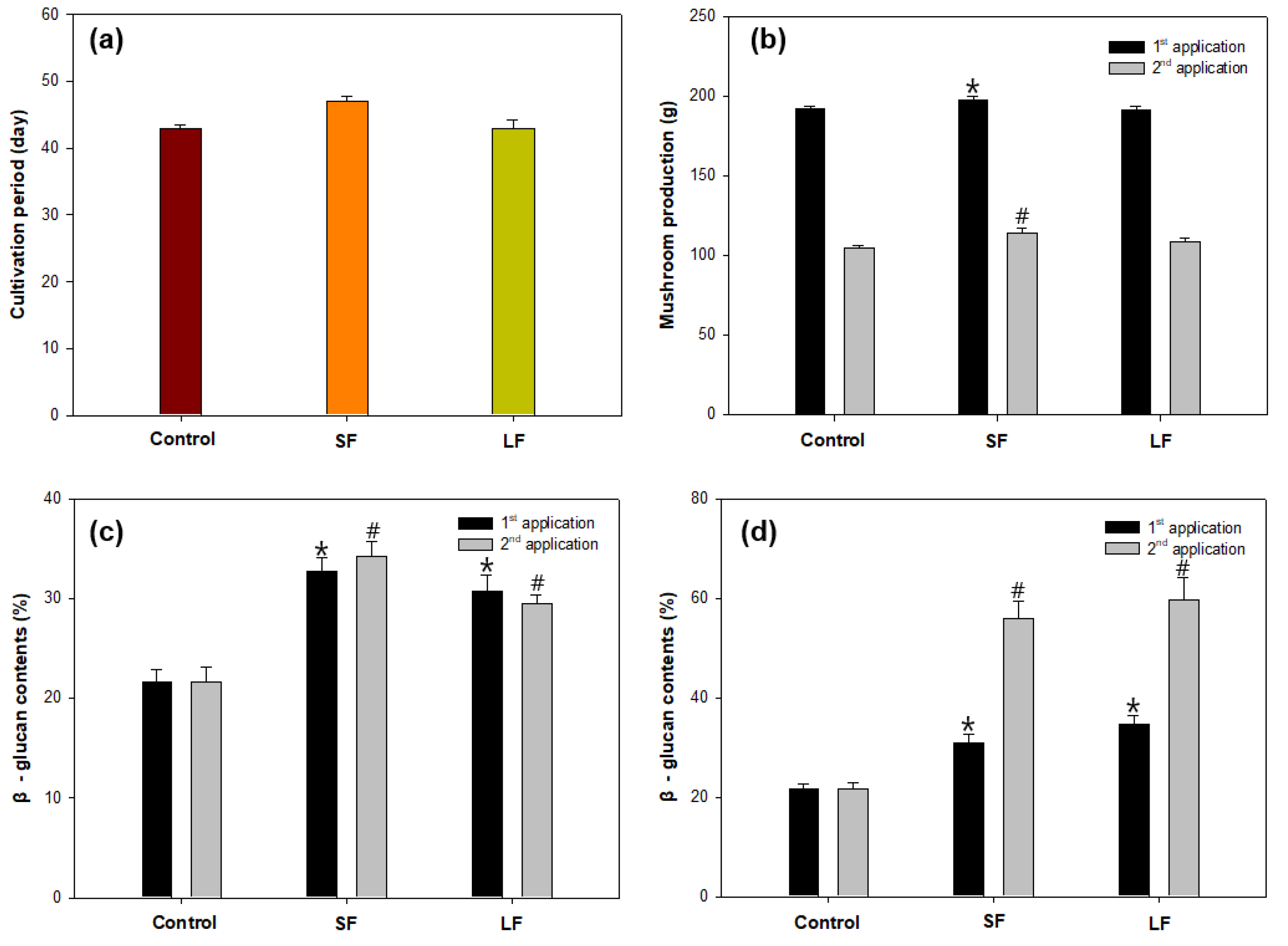
3.3. Elicitation by Alginate on β-Glucan Contents
3.4. Alginate Effects on Total Polyphenol Content
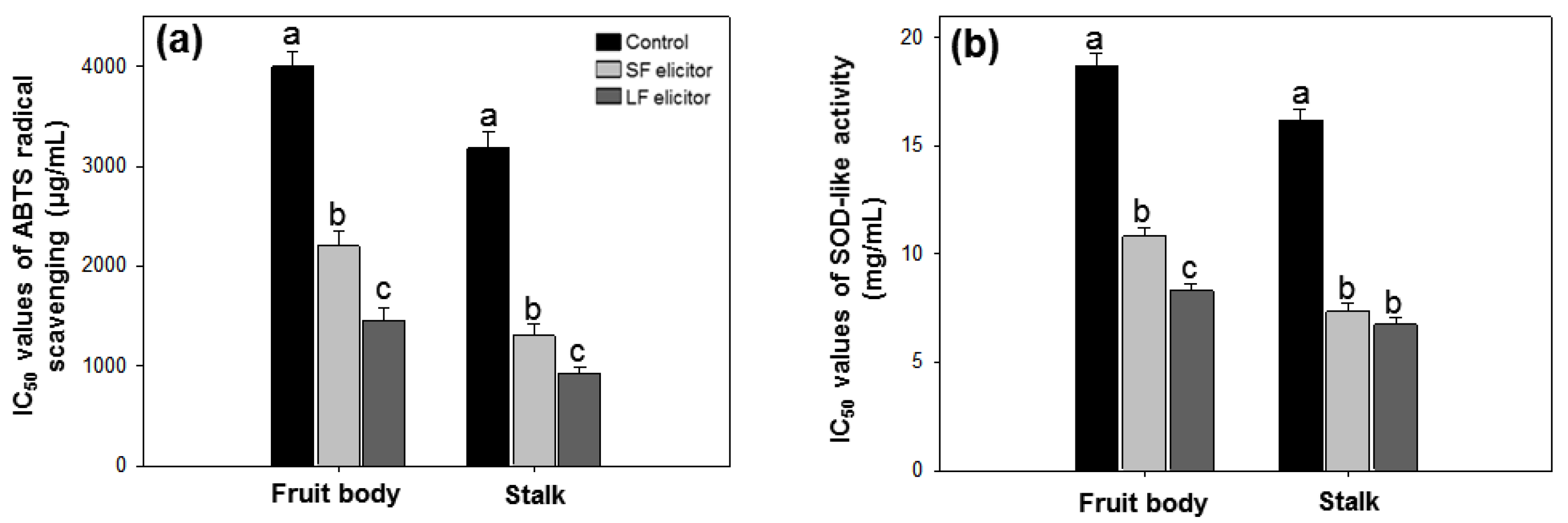
3.5. Alginate Effects on Antioxidant Activities
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Roncero-Ramos, I.; Delgado-Andrade, C. The beneficial role of edible mushrooms in human health. Curr. Opin. Food Sci. 2017, 14, 122–128. [Google Scholar] [CrossRef]
- Kimura, T. Natural products and biological activity of the pharmacologically active cauliflower mushroom Sparassis crispa. BioMed Res. Int. 2013, 2013, 982317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryoo, R.; Sou, H.D.; Ka, K.H.; Park, H. Phylogenetic relationships of Korean Sparassis latifolia based on morphological and its rDNA characteristics. J. Microbiol. 2013, 51, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Feng, B.; Yang, Z.L.; Dai, Y.C.; Wang, Z.; Tolgor, B. New species and distinctive geographical divergences of the genus Sparassis (Basidiomycota): Evidence from morphological and molecular data. Mycol. Prog. 2013, 12, 445–454. [Google Scholar] [CrossRef]
- Ryu, S.R.; Ka, K.H.; Park, H.; Bak, W.C.; Lee, B.H. Cultivation characteristics of Sparassis crispa strains using sawdust medium of Larix kaempferi. Korean J. Mycol. 2009, 37, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekaran, G.; Oh, D.S.; Shin, H.J. Properties and potential applications of the culinary-medicinal cauliflower mushroom, Sparassis crispa Wulf.Fr. (Aphyllophoromycetideae): A review. Int. J. Med. Mushrooms 2011, 13, 177–183. [Google Scholar] [CrossRef]
- Yoshikawa, K.; Kokudo, N.; Hashimoto, T.; Yamamoto, K.; Inose, T.; Kimura, T. Novel phthalide compounds from Sparassis crispa (Hanabiratake), Hanabiratakelide A-C, exhibiting anti-cancer related activity. Biol. Pharm. Bull. 2010, 33, 1355–1359. [Google Scholar] [CrossRef] [Green Version]
- Bang, S.; Chae, H.S.; Lee, C.; Choi, H.G.; Ryu, J.; Li, W.; Lee, H.; Jeong, G.S.; Chin, Y.W.; Shim, S.H. New aromatic compounds from the fruiting body of Sparassis crispa (Wulf.) and their inhibitory activities on proprotein convertase subtilisin/kexin type 9 mRNA expression. J. Agric. Food Chem. 2017, 65, 6152–6157. [Google Scholar] [CrossRef]
- Ruthes, A.C.; Smiderle, F.R.; Iacomini, M. D-Glucans from edible mushrooms: A review on the extraction, purification and chemical characterization approaches. Carbohyd. Polym. 2015, 117, 753–761. [Google Scholar] [CrossRef]
- Synytsya, A.; Novák, M. Structural diversity of fungal glucans. Carbohyd. Polym. 2013, 92, 792–809. [Google Scholar] [CrossRef] [PubMed]
- Smiderle, F.R.; Alquini, G.; Tadra-Sfeir, M.Z.; Iacomini, M.; Wichers, H.J.; van Griensven, L.J. Agaricus bisporus and Agaricus brasiliensis (1→6)-β-d-glucans show immunostimulatory activity on human THP-1 derived macrophages. Carbohyd. Polym. 2013, 94, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Morales, D.; Smiderle, F.R.; Villalva, M.; Abreu, H.; Rico, C.; Santoyo, S.; Iacomini, M.; Soler-Rivas, C. Testing the effect of combining innovative extraction technologies on the biological activities of obtained β-glucan-enriched fractions from Lentinula edodes. J. Funct. Foods 2019, 60, 103446. [Google Scholar] [CrossRef]
- Chan, G.C.; Chan, W.K.; Sze, D.M. The effects of β-glucan on human immune and cancer cells. J. Hematol. Oncol. 2009, 2, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohno, N.; Miura, N.N.; Nakajima, M.; Yadomae, T. Antitumor 1,3-β-glucan from cultured fruit body of Sparassis crispa. Biol. Pharm. Bull. 2000, 23, 866–872. [Google Scholar] [CrossRef] [Green Version]
- Harada, T.; Miura, N.N.; Adachi, Y.; Nakajima, M.; Yadomae, T.; Ohno, N. IFN-γ induction by SCG, 1, 3-β-D-glucan from Sparassis crispa, in DBA/2 mice in vitro. J. Interf. Cytok. Res. 2002, 22, 1227–1239. [Google Scholar] [CrossRef]
- Harada, T.; Kawaminami, H.; Miura, N.N.; Adachi, Y.; Nakajima, M.; Yadomae, T.; Ohno, N. Comparison of the immunomodulating activities of 1,3-β-glucan fractions from the culinary-medicinal mushroom Sparassis crispa Wulf.: Fr. (Aphyllophoromycetideae). Int. J. Med. Mushrooms 2006, 8, 231–244. [Google Scholar] [CrossRef]
- Tada, R.; Harada, T.; Nagi-Miura, N.; Adachi, Y.; Nakajima, M.; Yadomae, T.; Ohno, N. NMR characterization of the structure of a β-(1→3)-D-glucan isolate from cultured fruit bodies of Sparassis crispa. Carbohyd. Res. 2007, 342, 2611–2618. [Google Scholar] [CrossRef]
- Nameda, S.; Harada, T.; Miura, N.N.; Adachi, Y.; Yadomae, T.; Nakajima, M.; Ohno, N. Enhanced cytokine synthesis of leukocytes by a β-glucan preparation, SCG, extracted from a medicinal mushroom, Sparassis crispa. Immunopharm. Immunot. 2003, 25, 321–335. [Google Scholar] [CrossRef]
- Park, H.G.; Shim, Y.Y.; Choi, S.O.; Park, W.M. New method development for nanoparticle extraction of water-soluble β-(1→3)-D-glucan from edible mushrooms, Sparassis crispa and Phellinus linteus. J. Agric. Food Chem. 2009, 57, 2147–2154. [Google Scholar] [CrossRef]
- Narayani, M.; Srivastava, S. Elicitation: A stimulation of stress in in vitro plant cell/tissue cultures for enhancement of secondary metabolite production. Phytochem. Rev. 2017, 16, 1227–1252. [Google Scholar] [CrossRef]
- Pettit, R.K. Small-molecule elicitation of microbial secondary metabolites. Microb. Biotechnol. 2011, 4, 471–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryoo, R.; Sou, H.D.; Ka, K.H.; Park, H. Elicitor-induced β-glucan contents in fruit body of cauliflower mushroom (Sparassis latifolia). For. Sci. Technol. 2018, 14, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Ka, K.H.; Ryu, S.R. Enhancement of β-glucan content in the cultivation of cauliflower mushroom (Sparassis latifolia) by elicitation. Mycobiology 2014, 42, 41–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salachna, P.; Grzeszczuk, M.; Meller, E.; Soból, M. Oligo-alginate with low molecular mass improves growth and physiological activity of Eucomis autumnalis under salinity stress. Molecules 2018, 23, 812. [Google Scholar] [CrossRef] [Green Version]
- Cai, Z.; Kastell, A.; Mewis, I.; Knorr, D.; Smetanska, I. Polysaccharide elicitors enhance anthocyanin and phenolic acid accumulation in cell suspension cultures of Vitis vinifera. Plant Cell Tissue Organ Cult. 2012, 108, 401–409. [Google Scholar] [CrossRef]
- Bouissil, S.; Alaoui-Talibi, E.; Pierre, G.; Michaud, P.; El Modafar, C.; Delattre, C. Use of alginate extracted from Moroccan brown algae to stimulate natural defense in date palm roots. Molecules 2020, 25, 720. [Google Scholar] [CrossRef] [Green Version]
- Ben Salah, I.; Aghrouss, S.; Douira, A.; Aissam, S.; El Alaoui-Talibi, Z.; Filali-Maltouf, A.; El Modafar, C. Seaweed polysaccharides as bio-elicitors of natural defenses in olive trees against verticillium wilt of olive. J. Plant Interact. 2018, 13, 248–255. [Google Scholar] [CrossRef]
- Pawar, S.N.; Edgar, K.J. Alginate derivatization: A review of chemistry, properties and applications. Biomaterials 2012, 33, 3279–3305. [Google Scholar] [CrossRef]
- Silva, T.H.; Alves, A.; Ferreira, B.M.; Oliveira, J.M.; Reys, L.L.; Ferreira, R.J.; Sousa, R.A.; Silva, S.S.; Mano, J.F.; Reis, R.L. Materials of marine origin: A review on polymers and ceramics of biomedical interest. Int. Mater. Rev. 2012, 57, 276–306. [Google Scholar] [CrossRef] [Green Version]
- Davis, T.A.; Ramirez, M.; Mucci, A.; Larsen, B. Extraction, isolation and cadmium binding of alginate from Sargassum spp. J. Appl. Phycol. 2004, 16, 275–284. [Google Scholar] [CrossRef]
- Asilonu, E.; Bucke, C.; Keshavarz, T. Enhancement of chrysogenin production in cultures of Penicillium chrysogenum by uronic acid oligosaccharides. Biotechnol. Lett. 2000, 22, 931–936. [Google Scholar] [CrossRef]
- Alban, S.; Schauerte, A.; Franz, G. Anticoagulant sulfated polysaccharides. I. Synthesis and structure-activity relationships of new pullulan sulfates. Carbohyd. Polym. 2001, 47, 267–276. [Google Scholar] [CrossRef]
- Grasdalen, H. High-field, 1H-n.m.r. spectroscopy of alginate: Sequential structure and linkage conformations. Carbohyd. Res. 1983, 118, 255–260. [Google Scholar] [CrossRef]
- Manns, D.; Deutschle, A.L.; Saake, B.; Meyer, A.S. Methodology for quantitative determination of the carbohydrate composition of brown seaweeds (Laminariaceae). RSC Adv. 2014, 4, 25736–25746. [Google Scholar] [CrossRef] [Green Version]
- McCleary, B.V.; Draga, A. Measurement of β-glucan in mushrooms and mycelial products. J. AOAC Int. 2016, 99, 364–373. [Google Scholar] [CrossRef] [Green Version]
- Wootton-Beard, P.C.; Moran, A.; Ryan, L. Stability of the total antioxidant capacity and total polyphenol content of 23 commercially available vegetable juices before and after in vitro digestion measured by FRAP, DPPH, ABTS and Folin–Ciocalteu methods. Food Res. Int. 2011, 44, 217–224. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C.A. Antioxidant activity applying an improved ABTS radical cation decolourising assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Odeyemi, S.; Dewar, J. Repression of acetaminophen-induced hepatotoxicity in HepG2 cells by polyphenolic compounds from Lauridia tetragona (L.f.) R.H. Archer. Molecules 2019, 24, 2118. [Google Scholar] [CrossRef] [Green Version]
- Haug, A.; Larsen, B.; Smidsrød, O. Uronic acid sequence in alginate from different sources. Carbohyd. Res. 1974, 32, 217–225. [Google Scholar] [CrossRef]
- Leal, D.; Matsuhiro, B.; Rossi, M.; Caruso, F. FT-IR spectra of alginic acid block fractions in three species of brown seaweeds. Carbohyd. Res. 2008, 343, 308–316. [Google Scholar] [CrossRef]
- Fenoradosoa, T.A.; Ali, G.; Delattre, C.; Laroche, C.; Petit, E.; Wadouachi, A.; Michaud, P. Extraction and characterization of an alginate from the brown seaweed Sargassum turbinarioides Grunow. J. Appl. Phycol. 2010, 22, 131–137. [Google Scholar] [CrossRef]
- Kim, S.R.; Kang, H.W.; Ro, H.S. Generation and evaluation of high β-glucan producing mutant strains of Sparassis crispa. Mycobiology 2013, 41, 159–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, C.S.; Ganesan, P.; Suresh, P.V.; Bhaskar, N. Seaweeds as a source of nutritionally beneficial compounds-a review. J. Food Sci. Technol. 2008, 45, 1. [Google Scholar]
- Sinha, S.; Astani, A.; Ghosh, T.; Schnitzler, P.; Ray, B. Polysaccharides from Sargassum tenerrimum: Structural features, chemical modification and anti-viral activity. Phytochemistry 2010, 71, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Yokoi, K.; Konomi, A. Toxicity of so-called edible hijiki seaweed (Sargassum fusiforme) containing inorganic arsenic. Regul. Toxicol. Pharmacol. 2012, 63, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Bi, F.; Iqbal, S.; Arman, M.; Ali, A.; Hassan, M.U. Carrageenan as an elicitor of induced secondary metabolites and its effects on various growth characters of chickpea and maize plants. J. Saudi Chem. Soc. 2011, 15, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Chandía, N.P.; Matsuhiro, B. Characterization of a fucoidan from Lessonia vadosa (Phaeophyta) and its anticoagulant and elicitor properties. Int. J. Biol. Macromol. 2008, 42, 235–240. [Google Scholar] [CrossRef]
- Aziz, A.; Poinssot, B.; Daire, X.; Adrian, M.; Bézier, A.; Lambert, B.; Joubert, J.M.; Pugin, A. Laminarin elicits defense responses in grapevine and induces protection against Botrytis cinerea and Plasmopara viticola. Mol. Plant Microbe Interact. 2003, 16, 1118–1128. [Google Scholar] [CrossRef] [Green Version]
- El Modafar, C.; Elgadda, M.; El Boutachfaiti, R.; Abouraicha, E.; Zehhar, N.; Petit, E.; El Alaoui- Talibi, Z.; Courtois, B.; Courtois, J. Induction of natural defense accompanied by salicylic acid-dependant systemic acquired resistance in tomato seedlings in response to biloelicitors isolated from green algae. Sci. Hortic. 2012, 138, 55–63. [Google Scholar] [CrossRef]
- Chandía, N.P.; Matsuhiro, B.; Mejías, E.; Moenne, A. Alginic acids in Lessonia vadosa: Partial hydrolysis and elicitor properties of the polymannuronic acid fraction. J. Appl. Phycol. 2004, 16, 127–133. [Google Scholar] [CrossRef]
- Jeong, G.T.; Park, D.H.; Ryu, H.W.; Hwang, B.; Woo, J.C.; Kim, D.; Kim, S.W. Production of antioxidant compounds by culture of Panax ginseng C.A. Meyer hairy roots: I. Enhanced production of secondary metabolite in hairy root cultures by elicitation. Appl. Biochem. Biotechnol. 2005, 121, 1147–1157. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kurachi, M.; Yamaguchi, K.; Oda, T. Stimulation of multiple cytokine production in mice by alginate oligosaccharides following intraperitoneal administration. Carbohyd. Res. 2007, 342, 1133–1137. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, M.; Kurachi, M.; Nakashima, T.; Kim, D.; Yamaguchi, K.; Oda, T.; Iwamoto, Y.; Muramatsu, T. Structure–activity relationship of alginate oligosaccharides in the induction of cytokine production from RAW264. 7 cells. FEBS Lett. 2005, 579, 4423–4429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, M.H.; Ki, S.; Lee, S.E.; Lee, G.; Shin, H.J. Enhanced GABA content from sodium alginate-induced Sparassis latifolia influences dendrite development in primary cortical neurons. J. Mushroom 2019, 17, 275–283. [Google Scholar]
- Kiyama, R.; Furutani, Y.; Kawaguchi, K.; Nakanishi, T. Genome sequence of the cauliflower mushroom Sparassis crispa (Hanabiratake) and its association with beneficial usage. Sci. Rep. 2018, 8, 16053. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Yield (%) | Mw (kDa) | M/G | FM | FG | FMM | FMG | FGG |
|---|---|---|---|---|---|---|---|---|
| Sargassum fusiforme | 37.84 ± 0.48 | 504.65 | 1.38 | 0.58 | 0.42 | 0.55 | 0.03 | 0.39 |
| S. fulvellum | 30.88 ± 1.51 | 461.07 | 0.88 | 0.47 | 0.53 | 0.43 | 0.04 | 0.49 |
| S. horneri | 34.11 ± 1.65 | 138.10 | 0.64 | 0.39 | 0.61 | 0.30 | 0.09 | 0.52 |
| Species | M/G | Partial Hydrolysis (Yields) | Reference | ||
|---|---|---|---|---|---|
| EF (%) | SF (%) | LF (%) | |||
| Sargassum fusiforme | 1.38 | 63.8 | 20.4 | 15.8 | This study |
| S. fulvellum | 0.88 | 65.0 | 13.9 | 21.1 | This study |
| S. horneri | 0.64 | 61.1 | 16.1 | 22.8 | This study |
| Ascophyllum nodosum | 1.85 | 52 | 35 | 13 | [39] |
| Chordaria flagelliformis | 0.90 | 21 | 28 | 51 | [39] |
| Desmarestia aculeata | 0.85 | 27 | 23 | 50 | [39] |
| Dictyosiphon foeniculaceus | 0.85 | 25 | 25 | 50 | [39] |
| Fucus serratus | 1.30 | 35 | 34 | 31 | [39] |
| Laminaria digitata | 1.45 | 34 | 43 | 23 | [39] |
| L. hyperborea, fronds | 1.35 | 26 | 43 | 31 | [39] |
| L. hyperborea, stripes | 0.65 | 25 | 15 | 60 | [39] |
| Pelvetia canaliculata | 1.50 | 38 | 37 | 25 | [38] |
| Pylaiella | 0.75 | 40 | 18 | 42 | [39] |
| Scytosiphon lomentaria | 1.15 | 25 | 35 | 40 | [39] |
| Spermatochnus paradoxus | 1.30 | 35 | 32 | 33 | [39] |
| Desmarestia ligulata | 0.58 | 3.7 | 25.1 | 56.4 | [40] |
| D. ligulata | 0.77 | 3.5 | 37.0 | 47.1 | [40] |
| Lessonia flavicans | 1.03 | 8.5 | 41.3 | 22.2 | [40] |
| Species | Fraction | Fuc | Rham | Arab | Gal | Glu | Man | Xyl | Mannu | Gulur | Glucu |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Sargassum fusiforme | SA | 26.38 ± 0.88 | ND | ND | 6.21 ± 0.16 | 0.29 ± 0.02 | 7.17 ± 0.25 | 4.50 ± 0.18 | 179.64 ± 2.37 | 167.57 ± 1.88 | 10.67 ± 0.29 |
| SF | 0.32 ± 0.01 | ND | ND | 0.10 ± 0.01 | ND | ND | ND | 54.29 ± 0.14 | 13.18 ± 0.51 | ND | |
| LF | 1.84 ± 0.03 | ND | ND | 0.69 ± 0.01 | ND | 0.082 ± 0.04 | 0.35 | 8.47 ± 0.09 | 107.57 ± 1.88 | ND | |
| EF | 7.68 ± 0.10 | ND | ND | 2.8 ± 0.04 | ND | 3.59 ± 0.04 | 0.87 ± 0.02 | 5.46 ± 0.02 | 11.38 ± 0.11 | 6.39 ± 0.06 | |
| S. fulvellum | SA | 24.21 ± 0.60 | 0.12 ± 0.01 | 0.03 ± 0.00 | 15.07 ± 0.32 | 0.35 ± 0.00 | 8.59 ± 0.36 | 4.60 ± 0.05 | 136.79 ± 1.82 | 167.19 ± 1.10 | 20.27 ± 0.22 |
| SF | 0.17 ± 0.00 | ND | ND | ND | 0.11 ± 0.00 | 0.48 ± 0.00 | 0.05 ± 0.00 | 42.07 ± 2.82 | 7.53 ± 6.79 | ND | |
| LF | 3.88 ± 0.05 | 0.03 ± 0.00 | 0.05 ± 0.00 | 2.40 ± 0.03 | 1.77 ± 0.02 | 2.09 ± 0.04 | 1.08 ± 0.03 | 26.18 ± 0.77 | 100.16 ± 0.54 | 4.11 ± 0.31 | |
| EF | 8.86 ± 0.06 | 0.05 ± 0.00 | 0.01 ± 0.00 | 4.96 ± 0.04 | 0.10 ± 0.03 | 4.42 ± 0.04 | 1.03 ± 0.04 | 9.11 ± 0.18 | 3.56 ± 0.20 | 4.24 ± 0.05 | |
| S. horneri | SA | 20.14 ± 0.20 | ND | ND | 13.45 ± 0.09 | ND | 5.70 ± 0.13 | 4.40 ± 0.24 | 127.37 ± 0.39 | 189.99 ± 0.36 | 16.93 ± 0.32 |
| SF | 0.24 ± 0.00 | ND | ND | ND | 0.19 ± 0.00 | 0.14 ± 0.00 | 0.09 ± 0.00 | 43.98 ± 5.70 | 7.89 ± 1.05 | ND | |
| LF | 3.26 ± 0.02 | 0.03 ± 0.00 | 0.01 ± 0.00 | 2.51 ± 0.02 | 0.16 ± 0.02 | 2.11 ± 0.05 | 0.34 ± 0.02 | 22.21 ± 1.08 | 135.61 ± 3.18 | 3.44 ± 0.31 | |
| EF | 9.26 ± 0.12 | 0.08 ± 0.00 | ND | 6.57 ± 0.04 | 0.16 ± 0.00 | 5.52 ± 0.08 | 1.03 ± 0.02 | 11.71 ± 0.39 | 4.77 ± 0.76 | 5.65 ± 0.32 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.-W.; Wu, Y.; Choi, M.-H.; Shin, H.-J.; Li, J. Alginate-Derived Elicitors Enhance β-Glucan Content and Antioxidant Activities in Culinary and Medicinal Mushroom, Sparassis latifolia. J. Fungi 2020, 6, 92. https://doi.org/10.3390/jof6020092
Kim Y-W, Wu Y, Choi M-H, Shin H-J, Li J. Alginate-Derived Elicitors Enhance β-Glucan Content and Antioxidant Activities in Culinary and Medicinal Mushroom, Sparassis latifolia. Journal of Fungi. 2020; 6(2):92. https://doi.org/10.3390/jof6020092
Chicago/Turabian StyleKim, Yong-Woon, Yuanzheng Wu, Moon-Hee Choi, Hyun-Jae Shin, and Jishun Li. 2020. "Alginate-Derived Elicitors Enhance β-Glucan Content and Antioxidant Activities in Culinary and Medicinal Mushroom, Sparassis latifolia" Journal of Fungi 6, no. 2: 92. https://doi.org/10.3390/jof6020092
APA StyleKim, Y. -W., Wu, Y., Choi, M. -H., Shin, H. -J., & Li, J. (2020). Alginate-Derived Elicitors Enhance β-Glucan Content and Antioxidant Activities in Culinary and Medicinal Mushroom, Sparassis latifolia. Journal of Fungi, 6(2), 92. https://doi.org/10.3390/jof6020092