Exploring the Antibacterial Activity of Pestalotiopsis spp. under Different Culture Conditions and Their Chemical Diversity Using LC–ESI–Q–TOF–MS


 ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Isolation and Identification of Fungal Isolates
2.3. Media Preparation and Cultivation of Fungal Strains
2.4. Extraction and Sample Preparation
2.5. Antibacterial Activity
2.6. Analysis of Organic Extracts by LC–MS
3. Results
3.1. Culture Conditions
3.2. Antibacterial Activity
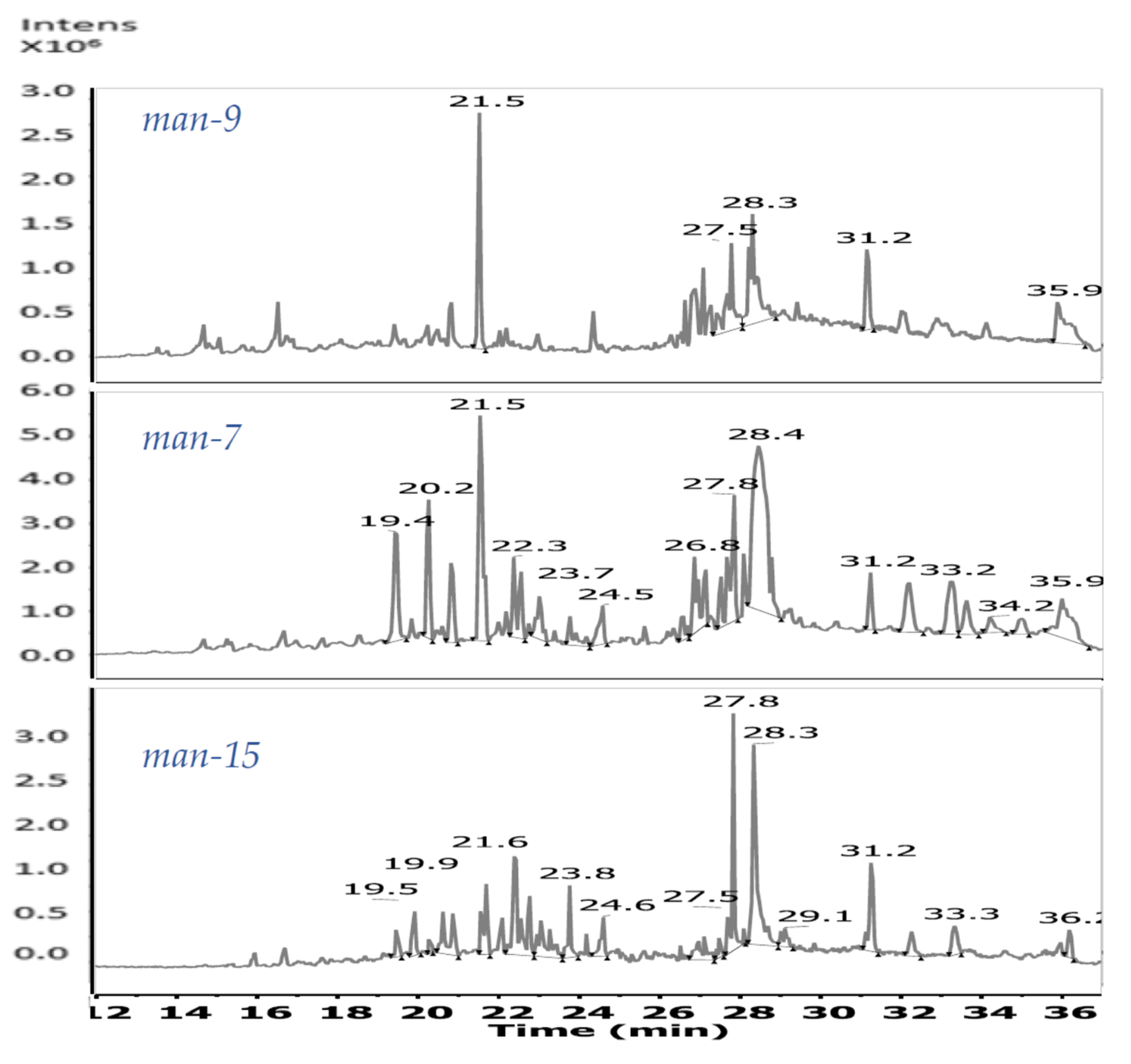
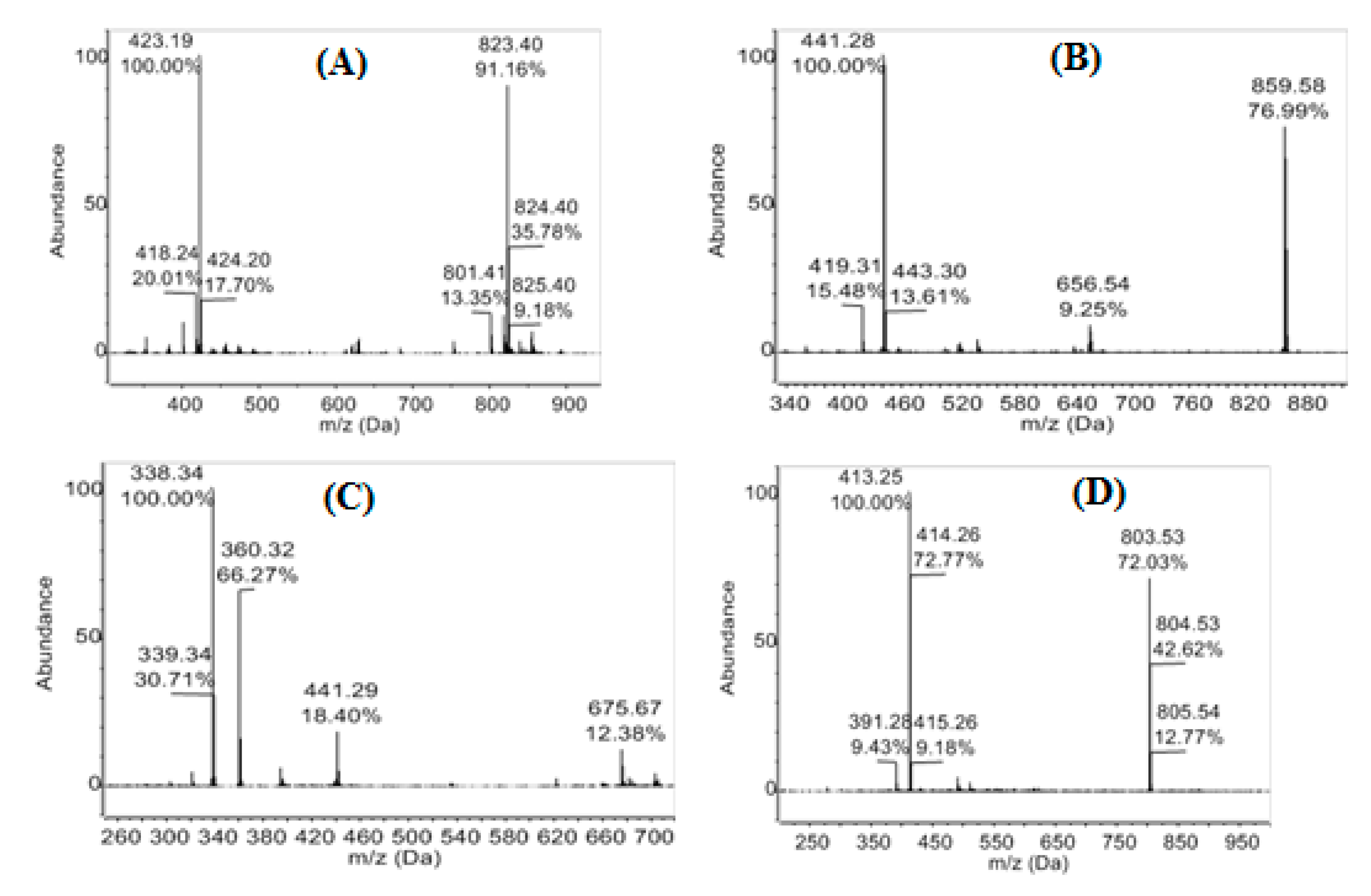
3.3. Evaluation of the Chemical Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Escolà-Vergé, L.; Los-Arcos, I.; Almirante, B. New antibiotics for the treatment of infections by multidrug-resistant microorganisms. Med. Clín. (Engl. Ed.) 2020, 154, 351–357. [Google Scholar] [CrossRef]
- Tacconelli, E.; Magrini, N. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Mani Chandrika, K.V.S.; Sharma, S. Promising antifungal agents: A minireview. Bioorg. Med. Chem. 2020, 28, 115398. [Google Scholar] [CrossRef] [PubMed]
- Outterson, K.; Rex, J.H. Evaluating for-profit public benefit corporations as an additional structure for antibiotic development and commercialization. Transl. Res. 2020, 220, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.C.; Kirchhelle, C.; Roberts, A.P. (Inter)nationalising the antibiotic research and development pipeline. Lancet Infect. Dis. 2020, 20, e54–e62. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Plant Endophytes and Epiphytes: Burgeoning Sources of Known and “Unknown” Cytotoxic and Antibiotic Agents? Planta Med. 2020. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Chaturvedi, P.; Kulkarni, M.G.; Van Staden, J. A critical review on exploiting the pharmaceutical potential of plant endophytic fungi. Biotechnol. Adv. 2020, 39, 107462. [Google Scholar] [CrossRef]
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; Brooks, S.; et al. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Fernández, R.E.; Sánchez-Ortiz, B.L.; Sandoval-Espinosa, Y.K.M.; Ulloa-Benítez, Á.; Armendáriz-Guillén, B.; García-Méndez, M.C.; Macías-Rubalcava, M.L. Hongos Endófitos: Fuente Potencial de Metabolitos Secundarios Bioactivos con Utilidad en Agricultura y Medicina. Tip Rev. Espec. Cienc. Quím. Biol. 2013, 16, 132–146. [Google Scholar] [CrossRef] [Green Version]
- Pye, C.R.; Bertin, M.J.; Lokey, R.S.; Gerwick, W.H.; Linington, R.G. Retrospective analysis of natural products provides insights for future discovery trends. Proc. Natl. Acad. Sci. USA 2017, 114, 5601–5606. [Google Scholar] [CrossRef] [Green Version]
- Torres-Mendoza, D.; Ortega, H.E.; Cubilla-Rios, L. Patents on Endophytic Fungi Related to Secondary Metabolites and Biotransformation Applications. J. Fungi 2020, 6, 58. [Google Scholar] [CrossRef]
- Urones, J.G.; Marcos, I.S.; Diez, D.; Cubilla, L.R. Tricyclic diterpenes from Hyptys dilatata. Phytochemistry 1998, 48, 1035–1038. [Google Scholar] [CrossRef]
- Yang, X.L.; Zhang, J.Z.; Luo, D.Q. The taxonomy, biology and chemistry of the fungal Pestalotiopsis genus. Nat. Prod. Rep. 2012, 29, 622–641. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Lei, J.; Wei, J.; Yao, N. Bioactive Natural Compounds from the Plant Endophytic Fungi Pestalotiopsis spp. Mini Rev. Med. Chem. 2012, 12, 1382–1393. [Google Scholar] [PubMed]
- Kumar Deshmukh, S.; Prakash, V.; Ranjan, H. Recent advances in the discovery of bioactive metabolites from Pestalotiopsis. Phytochem. Rev. 2017, 16, 883–920. [Google Scholar] [CrossRef]
- Xu, J.; Yang, X.; Lin, Q. Chemistry and biology of Pestalotiopsis-derived natural products. Fungal Divers. 2014, 66, 37–68. [Google Scholar] [CrossRef]
- Xu, J.; Ebada, S.S.; Proksch, P. Pestalotiopsis a highly creative genus: Chemistry and bioactivity of secondary metabolites. Fungal Divers. 2010, 44, 15–31. [Google Scholar] [CrossRef]
- Helaly, S.E.; Thongbai, B.; Stadler, M. Diversity of biologically active secondary metabolites from endophytic and saprotrophic fungi of the ascomycete order Xylariales. Nat. Prod. Rep. 2018, 35, 992–1014. [Google Scholar] [CrossRef]
- Ortega, H.E.; Shen, Y.Y.; Tendyke, K.; Ríos, N.; Cubilla-Ríos, L. Polyhydroxylated macrolide isolated from the endophytic fungus Pestalotiopsis mangiferae. Tetrahedron Lett. 2014, 55, 2642–2645. [Google Scholar] [CrossRef]
- Maram, L.; Das, B. The First Stereoselective Total Synthesis of Mangiferaelactone: An Antibacterial Fungal Nonanolide. Synlett 2014, 25, 2327–2330. [Google Scholar]
- Kumar, R.; Rej, R.K.; Nanda, S. Asymmetric total synthesis of (-)-mangiferaelactone by using an appropriately substituted thiophene as a masked synthon for C-alkyl glycoside. Tetrahedron Asymmetry 2015, 26, 751–759. [Google Scholar] [CrossRef]
- Reddy, B.V.S.; Reddy, P.S.; Babu, K.V.; Reddy, B.P.; Yadav, J.S. Stereoselective Total Synthesis of Mangiferaelactone using D-Mannose as a Chiral Pool. Helv. Chim. Acta 2015, 98, 1395–1402. [Google Scholar] [CrossRef]
- Strobel, G.; Yang, X.; Sears, J.; Kramer, R.; Sidhu, R.S.; Hess, W.M. Taxol from Pestalotiopsis microspora, an endophytic fungus of Taxus wallachiana. Microbiology 1996, 142, 3–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strobel, G.; Ford, E.; Worapong, J.; Harper, J.K.; Arif, A.M.; Grant, D.M.; Fung, P.C.W.; Ming Wah Chau, R. Isopestacin, an isobenzofuranone from Pestalotiopsis microspora, possessing antifungal and antioxidant activities. Phytochemistry 2002, 60, 179–183. [Google Scholar] [CrossRef]
- Liu, S.; Dai, H.; Makhloufi, G.; Heering, C.; Janiak, C.; Hartmann, R.; Mándi, A.; Kurtán, T.; Müller, W.E.G.; Kassack, M.U.; et al. Cytotoxic 14-Membered Macrolides from a Mangrove-Derived Endophytic Fungus, Pestalotiopsis microspora. J. Nat. Prod. 2016, 79, 2332–2340. [Google Scholar] [CrossRef]
- Nalin Rathnayake, G.R.; Savitri Kumar, N.; Jayasinghe, L.; Araya, H.; Fujimoto, Y. Secondary Metabolites Produced by an Endophytic Fungus Pestalotiopsis microspora. Nat. Prod. Bioprospect. 2019, 9, 411–417. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, S.; Bohni, N.; Schnee, S.; Schumpp, O.; Gindro, K.; Wolfender, J.-L. Metabolite induction via microorganism co-culture: A potential way to enhance chemical diversity for drug discovery. Biotechnol. Adv. 2014, 32, 1180–1204. [Google Scholar] [CrossRef]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big Effects from Small Changes: Possible Ways to Explore Nature’s Chemical Diversity. Chembiochem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Takahashi, J.A.; Campos Teles, A.P.; De Almeida Pinto Bracarense, A.; Corrêia Gomes, D. Classical and epigenetic approaches to metabolite diversification in filamentous fungi. Phytochem. Rev. 2013, 12, 773–789. [Google Scholar] [CrossRef]
- Frey-Klett, P.; Burlinson, P.; Deveau, A.; Barret, M.; Tarkka, M.; Sarniguet, A. Bacterial-Fungal Interactions: Hyphens between Agricultural, Clinical, Environmental, and Food Microbiologists. Microbiol. Mol. Biol. Rev. 2011, 75, 583–609. [Google Scholar] [CrossRef] [Green Version]
- Pettit, R.K. Mixed fermentation for natural product drug discovery. Appl. Microbiol. Biotechnol. 2009, 83, 19–25. [Google Scholar] [CrossRef]
- Shank, E.A.; Kolter, R. New developments in microbial interspecies signaling. Curr. Opin. Microbiol. 2009, 12, 205–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adelin, E.; Slimani, N.; Cortial, S.; Schmitz-Alfonso, I.; Ouazzani, J. Platotex: An innovative and fully automated device for cell growth scale-up of agar-supported solid-state fermentation. J. Ind. Microbiol. Biotechnol. 2011, 38, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Kjærbølling, I.; Mortensen, U.H.; Vesth, T.; Andersen, M.R. Strategies to establish the link between biosynthetic gene clusters and secondary metabolites. Fungal Genet. Biol. 2019, 130, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Ramesha, K.P.; Mohana, N.C.; Nuthan, B.R.; Rakshith, D.; Satish, S. Epigenetic modulations of mycoendophytes for novel bioactive molecules. Biocatal. Agric. Biotechnol. 2018, 16, 663–668. [Google Scholar] [CrossRef]
- Chaichanan, J.; Wiyakrutta, S.; Pongtharangkul, T.; Isarangkul, D.; Meevootisom, V. Optimization of zofimarin production by an endophytic fungus, Xylaria sp. Acra L38. Braz. J. Microbiol. 2014, 45, 287–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacheco Fill, T.; Pallini, H.F.; da Silva Amaral, L.; da Silva, J.V.; Lazarin Bidóia, D.; Peron, F.; Pelegrin Garcia, F.; Vataru Nakamura, C.; Rodrigues-Filho, E. Copper and Manganese Cations Alter Secondary Metabolism in the Fungus Penicillium brasilianum. J. Braz. Chem. Soc. 2016, 27, 1444–1451. [Google Scholar]
- Fillat, Ú.; Martín-Sampedro, R.; Macaya-Sanz, D.; Martín, J.A.; Ibarra, D.; Martínez, M.J.; Eugenio, M.E. Screening of eucalyptus wood endophytes for laccase activity. Process Biochem. 2016, 51, 589–598. [Google Scholar] [CrossRef]
- Goutam, J.; Sharma, V.K.; Verma, S.K.; Singh, D.K.; Kumar, J.; Mishra, A.; Kumar, A.; Kharwar, R.N. Optimization of Culture Conditions for Enhanced Production of Bioactive Metabolites Rich in Antimicrobial and Antioxidant Activities Isolated from Emericella quadrilineata an Endophyte of Pteris pellucida. J. Pure Appl. Microbiol. 2014, 8, 2059–2073. [Google Scholar]
- Li, P.; Xu, L.; Mou, Y.; Shan, T.; Mao, Z.; Lu, S.; Peng, Y.; Zhou, L. Medium optimization for exopolysaccharide production in liquid culture of endophytic fungus Berkleasmium sp. Dzf12. Int. J. Mol. Sci. 2012, 13, 11411–11426. [Google Scholar] [CrossRef]
- Mou, Y.; Luo, H.; Mao, Z.; Shan, T.; Sun, W.; Zhou, K.; Zhou, L. Enhancement of palmarumycins C12 and C13 production in liquid culture of endophytic fungus Berkleasmium sp. Dzf12 after treatments with metal ions. Int. J. Mol. Sci. 2013, 14, 979–998. [Google Scholar] [CrossRef] [PubMed]
- Nomila Merlin, J.; Nimal Christhudas, I.V.S.; Praveen Kumar, P.; Agastian, P. Optimization of growth and bioactive metabolite production: Fusarium solani. Asian J. Pharm. Clin. Res. 2013, 6, 98–103. [Google Scholar]
- Panuthai, T.; Sihanonth, P.; Piapukiew, J.; Sooksai, S.; Sangvanich, P.; Karnchanatat, A. An extracellular lipase from the endophytic fungi Fusarium oxysporum isolated from the Thai medicinal plant, Croton oblongifolius Roxb. Afr. J. Microbiol. Res. 2012, 6, 2622–2638. [Google Scholar]
- Sorgatto, M.; Guimaraes, N.C.A.; Zanoelo, F.F.; Marques, M.R.; Peixoto-Nogueira, S.C.; Giannesi, G.G. Purification and characterization of an extracellular xylanase produced by the endophytic fungus, Aspergillus terreus, grown in submerged fermentation. Afr. J. Biotechnol. 2012, 11, 8076–8084. [Google Scholar] [CrossRef]
- Zhao, X.-M.; Wang, Z.-Q.; Shu, S.-H.; Wang, W.-J.; Xu, H.-J.; Ahn, Y.-J.; Wang, M.; Hu, X. Ethanol and Methanol Can Improve Huperzine a Production from Endophytic Colletotrichum gloeosporioides ES026. PLoS ONE 2013, 8, e61777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Wang, X.; Sun, W.; Mou, Y.; Peng, Y.; Zhou, L. Medium optimization for palmarumycin C13 production in liquid culture of endophytic fungus Berkleasmium sp. Dzf12 using response surface methodology. Electron. J. Biotechnol. 2013, 16, 16. [Google Scholar] [CrossRef]
- Kaeberlein, T.; Lewis, K.; Epstein, S.S. Isolating “Uncultivable” Microorganisms in Pure Culture in a Simulated Natural Environment. Science 2002, 296, 1127–1129. [Google Scholar] [CrossRef] [Green Version]
- Wohlleben, W.; Mast, Y.; Stegmann, E.; Ziemert, N. Antibiotic drug discovery. Microb. Biotechnol. 2016, 9, 541–548. [Google Scholar] [CrossRef] [Green Version]
- Netzker, T.; Fischer, J.; Weber, J.; Mattern, D.J.; König, C.C.; Valiante, V.; Schroeckh, V.; Brakhage, A.A. Microbial communication leading to the activation of silent fungal secondary metabolite gene clusters. Front. Microbiol. 2015, 6, 1–13. [Google Scholar] [CrossRef]
- Molinar, E.; Rios, N.; Spadafora, C.; Elizabeth Arnold, A.; Coley, P.D.; Kursar, T.A.; Gerwick, W.H.; Cubilla-Rios, L. Coibanoles, a new class of meroterpenoids produced by Pycnoporus sanguineus. Tetrahedron Lett. 2012, 53, 919–922. [Google Scholar] [CrossRef] [Green Version]
- U’Ren, J.M.; Lutzoni, F.; Miadlikowska, J.; Laetsch, A.D.; Elizabeth Arnold, A. Host and geographic structure of endophytic and endolichenic fungi at a continental scale. Am. J. Bot. 2012, 99, 898–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, J.M. BSAC standardized disc susceptibility testing method (version 8). J. Antimicrob. Chemother. 2009, 64, 454–489. [Google Scholar] [CrossRef] [PubMed]
- Vásquez, R.; Rios, N.; Solano, G.; Cubilla-Rios, L. Lentinoids A–D, New Natural Products Isolated from Lentinus strigellus. Molecules 2018, 23, 773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorgensen, J.H.; Turnidge, J.D. Susceptibility Test Methods Dilution and Disk Diffusion Methods. In Manual of Clinical Microbiology, 11th ed.; Jorgensen, J., Pfaller, M., Carrol, K., Funke, G., Landry, M., Richter, S., Warnock, D., Eds.; ASM Press: Washington, DC, USA, 2015; pp. 1253–1273. [Google Scholar]
- Subban, K.; Subramani, R.; Johnpaul, M. A novel antibacterial and antifungal phenolic compound from the endophytic fungus Pestalotiopsis mangiferae. Nat. Prod. Res. 2013, 27, 1445–1449. [Google Scholar] [CrossRef]
- Cutler, H.G.; Himmelsbach, D.S.; Jacyno, J.M.; Cole, P.D.; Yagen, B.; Arrendale, R.F.; Cox, R.H. Koninginin B: A Biologically Active Congener of Koninginin A from Trichoderma koningii. J. Agric. Food Chem. 1991, 39, 977–980. [Google Scholar] [CrossRef]
- Ghisalberti, E.L.; Rowland, C.Y. Antifungal Metabolites from Trichoderma harzianum. J. Nat. Prod. 1993, 56, 1799–1804. [Google Scholar] [CrossRef]
- Parker, S.R.; Cutler, H.G.; Schreiner, P.R. Koninginin E: Isolation of a Biologically Active Natural Product from Trichoderma koningii. Biosci. Biotechnol. Biochem. 1995, 59, 1747–1749. [Google Scholar] [CrossRef] [Green Version]
- Luo, D.-Q.; Deng, H.-Y.; Yang, X.-L.; Shi, B.-Z.; Zhang, J.-Z. Oleanane-Type Triterpenoids from the Endophytic Fungus Pestalotiopsis clavispora Isolated from the Chinese Mangrove Plant Bruguiera sexangula. Helv. Chim. Acta 2011, 94, 1041–1047. [Google Scholar] [CrossRef]
- Fu, S.B.; Yang, J.S.; Cui, J.L.; Meng, Q.F.; Feng, X.; Sun, D.A. Multihydroxylation of ursolic acid by Pestalotiopsis microspora isolated from the medicinal plant Huperzia serrata. Fitoterapia 2011, 82, 1057–1061. [Google Scholar] [CrossRef]
- Evidente, A.; Amalfitano, C.; Pengue, R.; Altomare, C. High performance liquid chromatography for the analysis of fusapyrone and deoxyfusapyrone, two antifungal α-pyrones from Fusarium semitectum. Nat. Toxins 1999, 7, 133–137. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungi | Extract | E. coli | P. aeruginosa | S. tiphy | S. flexneri | P. vulgaris | S. enterica | L. pneumophIlla | E. faecalis | E. cloacae | P. multocida | K. rhizophila | L. monocytogenes | B. cereus | S. oralis |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P. mangiferae | man-1 | 8.5 | 9.5 | ||||||||||||
| man-2 | 8.5 | 8.5 | 8.0 | ||||||||||||
| man-3 | 8.5 | 7.5 | 9.0 | 9.0 | 7.5 | ||||||||||
| man-6 | 10.0 | 8.5 | 9.0 | 9.0 | 8.0 | 11.0 | 8.0 | ||||||||
| man-7 | 8.0 | 8.5 | 8.5 | 9.0 | 7.5 | ||||||||||
| man-9 | 12.5 | 8.5 | 8.5 | 8.0 | 7.5 | 7.5 | |||||||||
| man-15 | 9.5 | ||||||||||||||
| P. microspora | mic-9 | 8.0 | 8.5 | 7.5 | 8.0 | 7.5 | 8.5 | ||||||||
| mic-10 | 8.0 | 8.5 | 8.5 | ||||||||||||
| mic-11 | 9.5 | ||||||||||||||
| mic-12 | 9.5 | 7.5 | 7.5 | ||||||||||||
| Positive control | Gentamycin sulfate (10 μg/mL) | 18.5 | 17.5 | 14.5 | 13.5 | 13.0 | 13.0 | 12.0 | 10.0 | 9.5 | 9.5 | 30.0 | 18.0 | 17.0 | 9.0 |
| Retention Time 1 (tR) | m/z | [M + H]+ | [M + Na]+ | Dimers | Molecular Formula | Extracts |
|---|---|---|---|---|---|---|
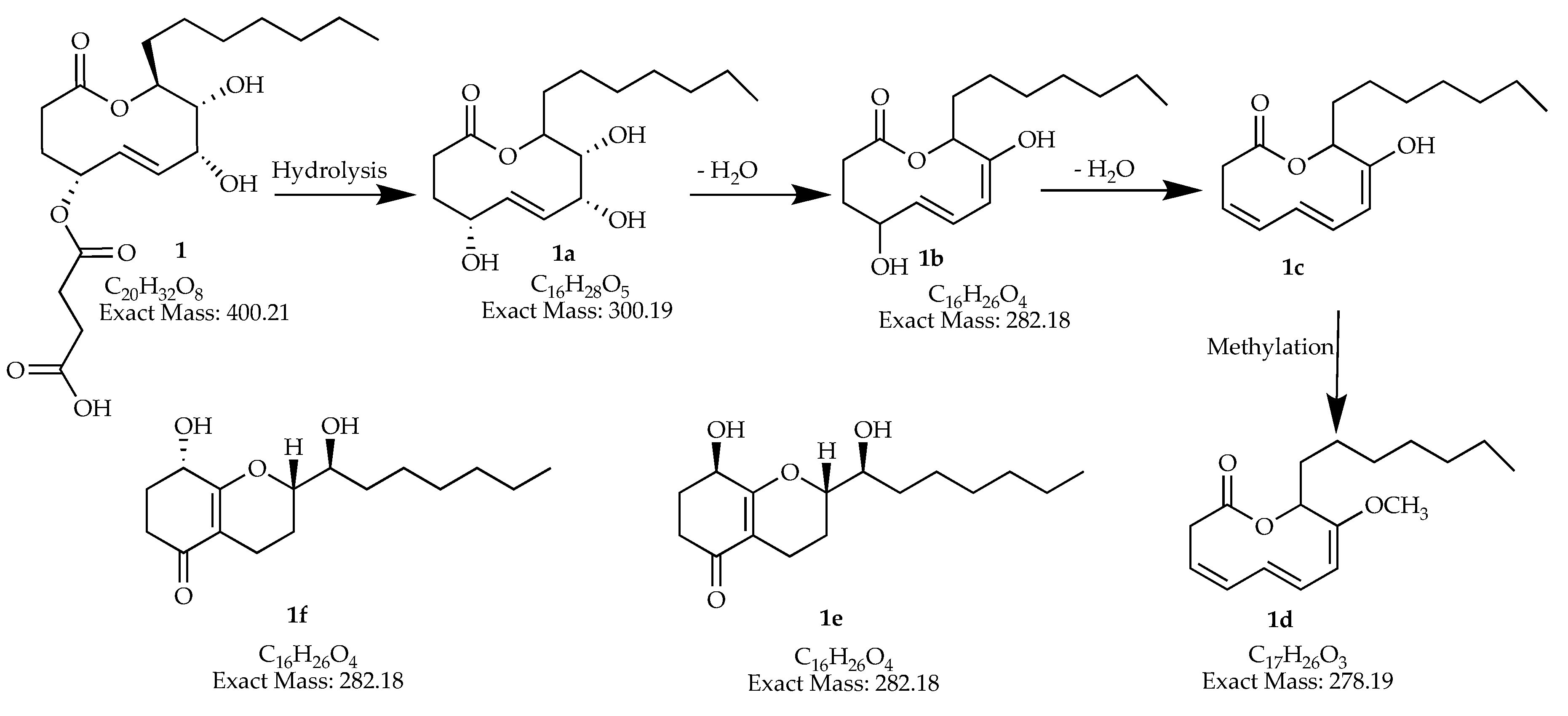
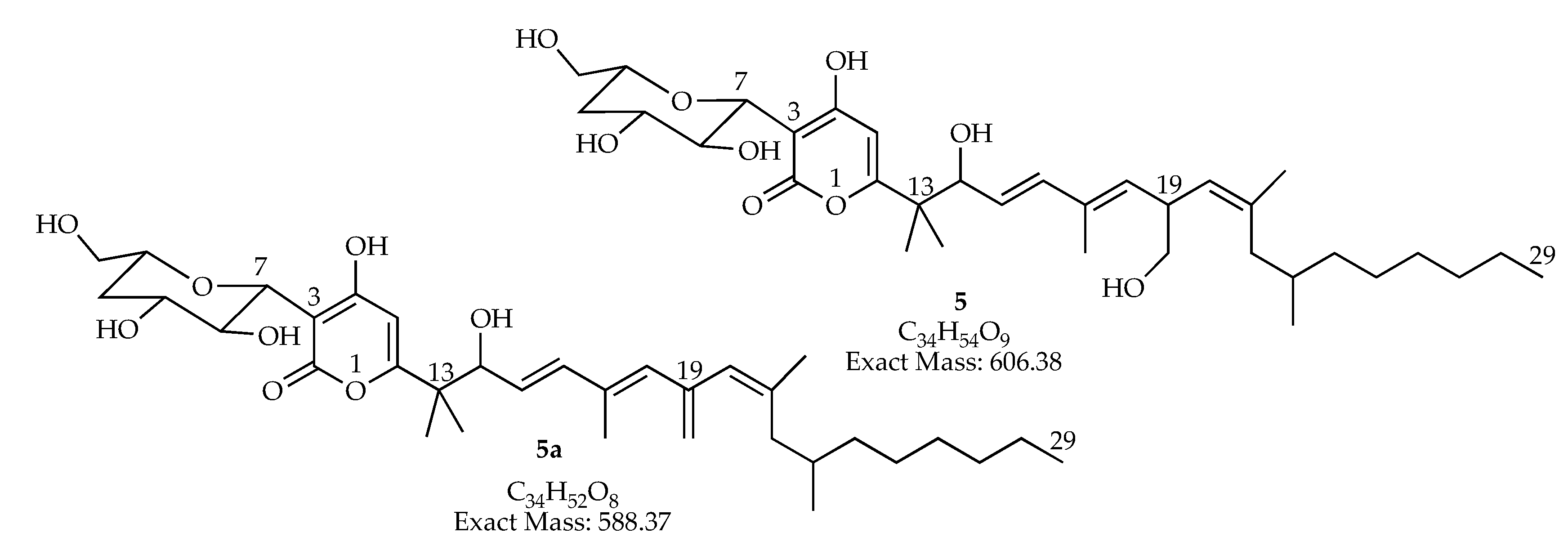
| 19.44–19.50 | 588.36567 | 589.369 | 1177.720 [2M + H]+ | C34H52O8 | man-7, man-15 | |
| 20.14–20.37 | 278.18765 | 279.194 | C17H26O3 | man-7 | ||
| 20.78–20.87 | 588.36567 | 589.369 | 1177.720 [2M + H]+ | C34H52O8 | man-7, man-15 | |
| 21.55–21.59 | 282.18524 | 283.193 | 565.375 [2M + H]+ | C16H26O4 | man-7, man-9, man-15 | |
| 400.20917 | 401.217 | 423.199 | 801.424 [2M + H]+ 823.407 [2M + Na]+ | C20H32O8 | man-7, man-9, man-15 | |
| 22.39–22.43 | 394.28663 | 395.295 | C27H38O2 | man-7, man-15 | ||
| 412.29317 | 413.303 | C22H40N2O5 | man-7, man-15 | |||
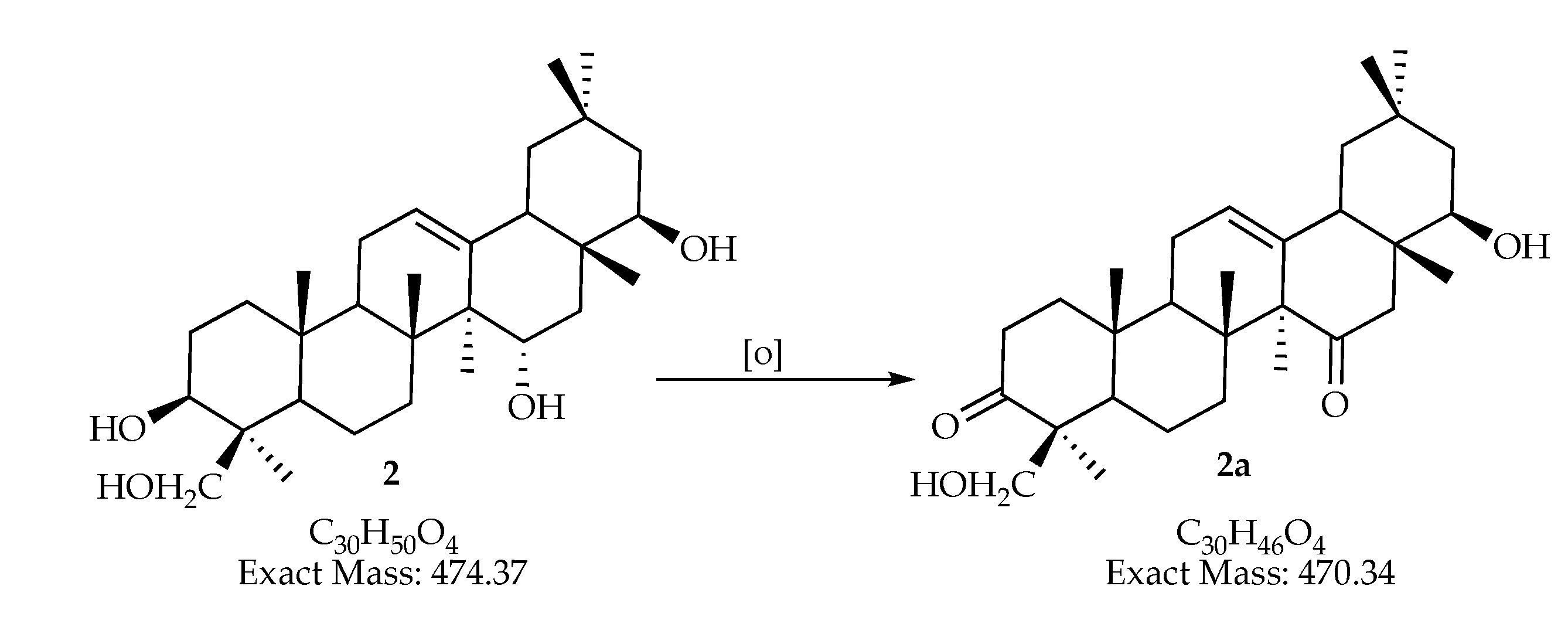
| 470.33906 | 471.347 | C30H46O4 | man-7, man-15 | |||
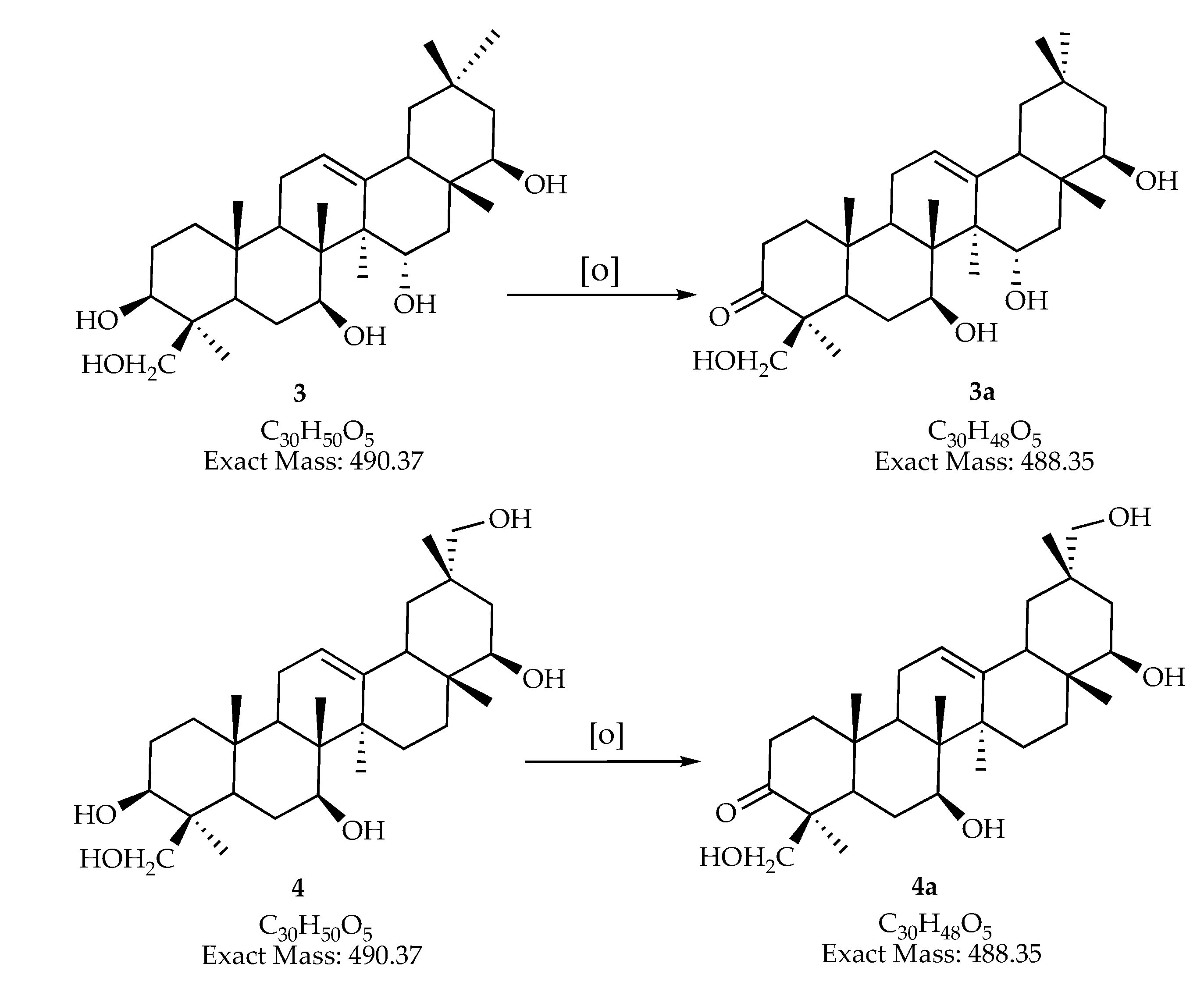
| 488.34560 | 489.354 | 995.716 [2M + H2O + H]+ | C30H48O5 | man-7, man-15 | ||
| 23.40–23.45 | 314.24516 | 315.256 | 337.236 | C38H34O4 | man-15 | |
| 23.72–23.81 | 428.30737 | 451.298 | C31H40O | man-7, man-15 | ||
| 428.31324 | 429.320 | C24H44O6 | man-15 | |||
| 24.54 | 278.22403 | 279.231 | 301.211 | 557.446 [2M + H]+ | C18H30O2 | man-7, man-15 |
| 24.60 | 452.31324 | 475.304 | 927.610 [2M + Na]+ | C26H44O6 | man-15 | |
| 26.86 | 281.27132 | 282.279 | 304.261 | C18H35NO | man-7, man-15 | |
| 27.52 | 283.28697 | 284.294 | 306.276 | C18H37NO | man-7, man-15 | |
| 27.83–27.87 | 390.27646 | 391.283 | 413.261 | 803.536 [2M + Na]+ | C24H38O4 | man-7, man-9, man-15 |
| 28–33 | 337.33392 | 338.341 | 360.320 | 675.670 [2M + H]+ | C22H43NO | man-9, man-15 |
| 28.40 | 418.28663 | 419.308 | 441.288 | 859.578 [2M + Na]+ | C29H38O2 | man-7 |
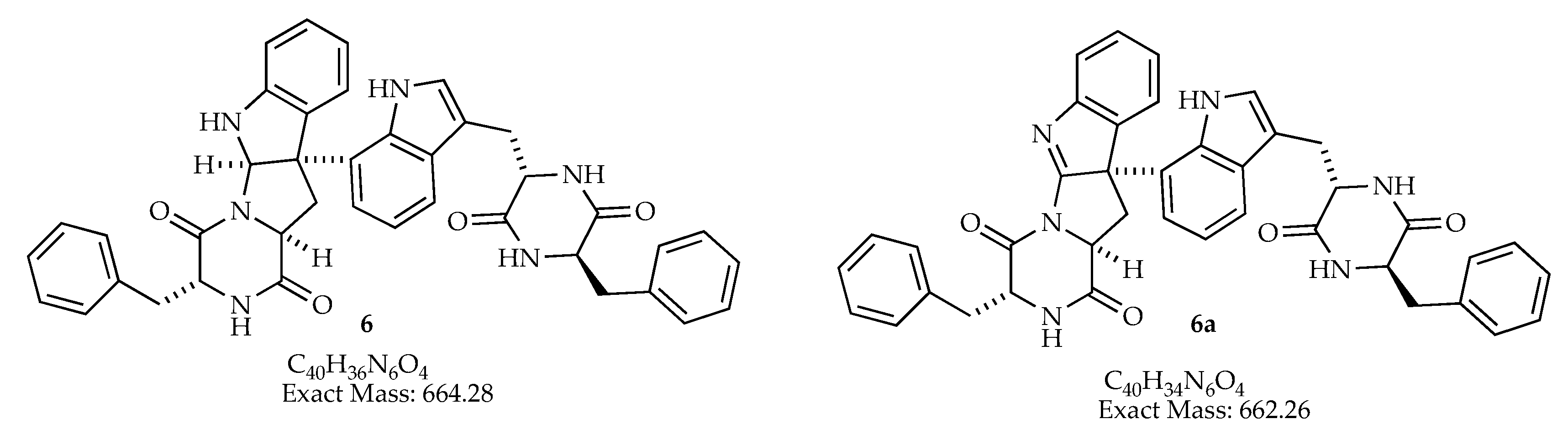
| 31.25–31.36 | 662.43884 | 663.444 | 685.40 | C40H36NO4 | man-7, man-9, man-15 | |
| 721.51233 | 722.519 | C41H71NO9 | man-15 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguilar-Pérez, M.M.; Torres-Mendoza, D.; Vásquez, R.; Rios, N.; Cubilla-Rios, L. Exploring the Antibacterial Activity of Pestalotiopsis spp. under Different Culture Conditions and Their Chemical Diversity Using LC–ESI–Q–TOF–MS. J. Fungi 2020, 6, 140. https://doi.org/10.3390/jof6030140
Aguilar-Pérez MM, Torres-Mendoza D, Vásquez R, Rios N, Cubilla-Rios L. Exploring the Antibacterial Activity of Pestalotiopsis spp. under Different Culture Conditions and Their Chemical Diversity Using LC–ESI–Q–TOF–MS. Journal of Fungi. 2020; 6(3):140. https://doi.org/10.3390/jof6030140
Chicago/Turabian StyleAguilar-Pérez, Madelaine M., Daniel Torres-Mendoza, Roger Vásquez, Nivia Rios, and Luis Cubilla-Rios. 2020. "Exploring the Antibacterial Activity of Pestalotiopsis spp. under Different Culture Conditions and Their Chemical Diversity Using LC–ESI–Q–TOF–MS" Journal of Fungi 6, no. 3: 140. https://doi.org/10.3390/jof6030140
APA StyleAguilar-Pérez, M. M., Torres-Mendoza, D., Vásquez, R., Rios, N., & Cubilla-Rios, L. (2020). Exploring the Antibacterial Activity of Pestalotiopsis spp. under Different Culture Conditions and Their Chemical Diversity Using LC–ESI–Q–TOF–MS. Journal of Fungi, 6(3), 140. https://doi.org/10.3390/jof6030140