Intranasal Vaccine Using P10 Peptide Complexed within Chitosan Polymeric Nanoparticles as Experimental Therapy for Paracoccidioidomycosis in Murine Model

 , , ,
, , ,  , ,
, , 
Abstract
:1. Introduction
2. Methods
2.1. P10-Chitosan Nanoparticles Preparation and Characterization
2.2. Physical-Chemical Characterization of Nanoparticles
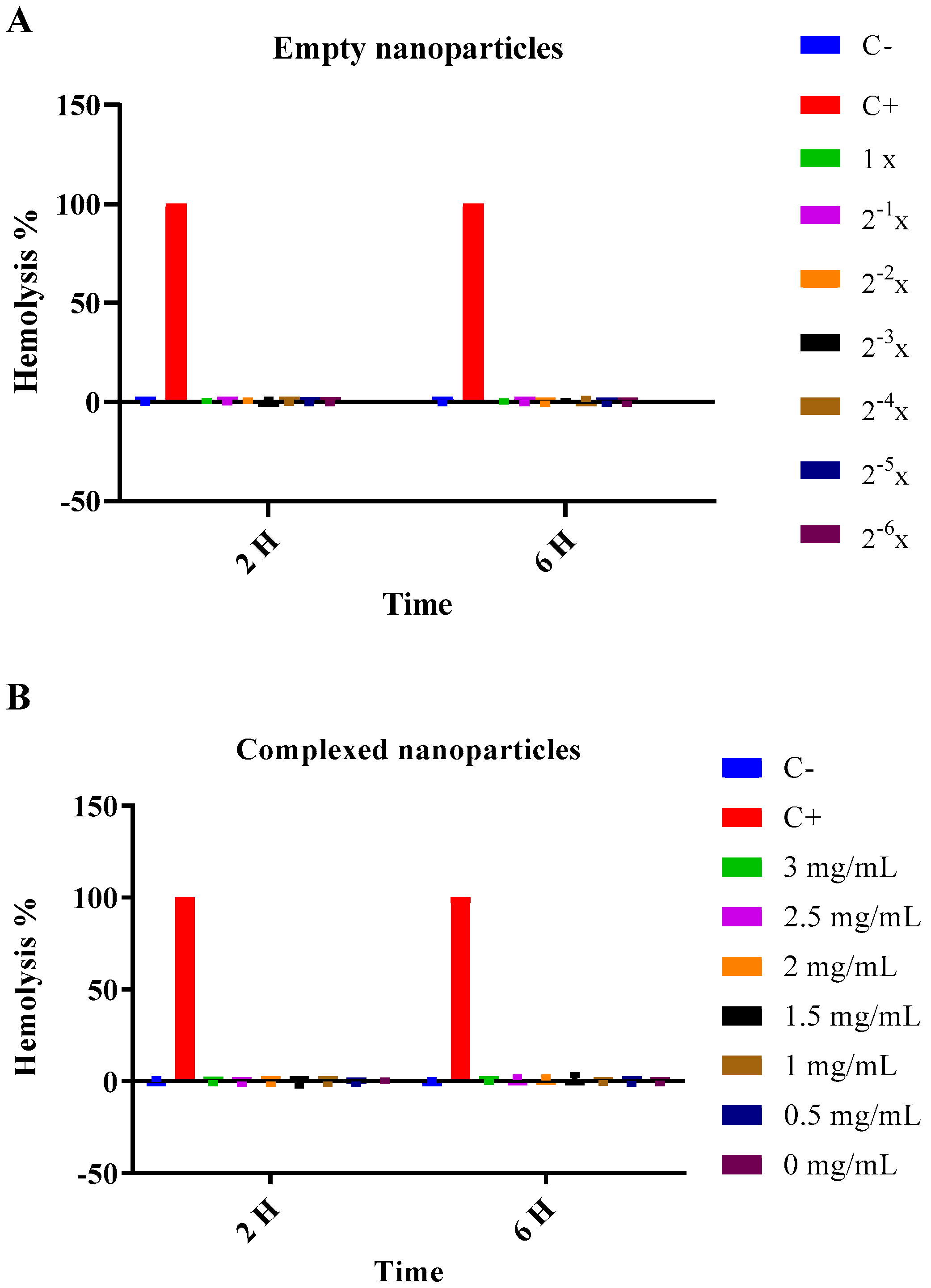
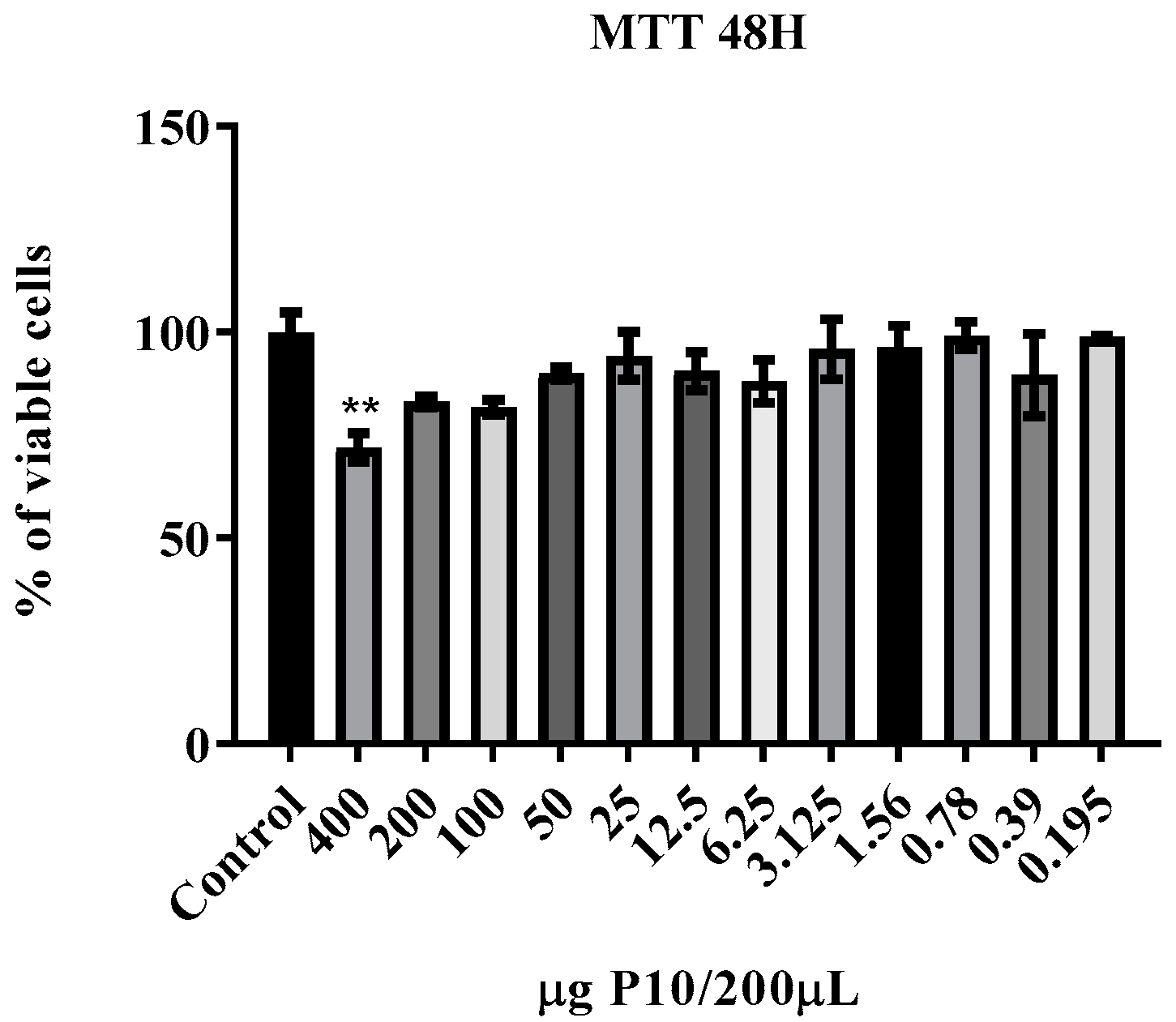
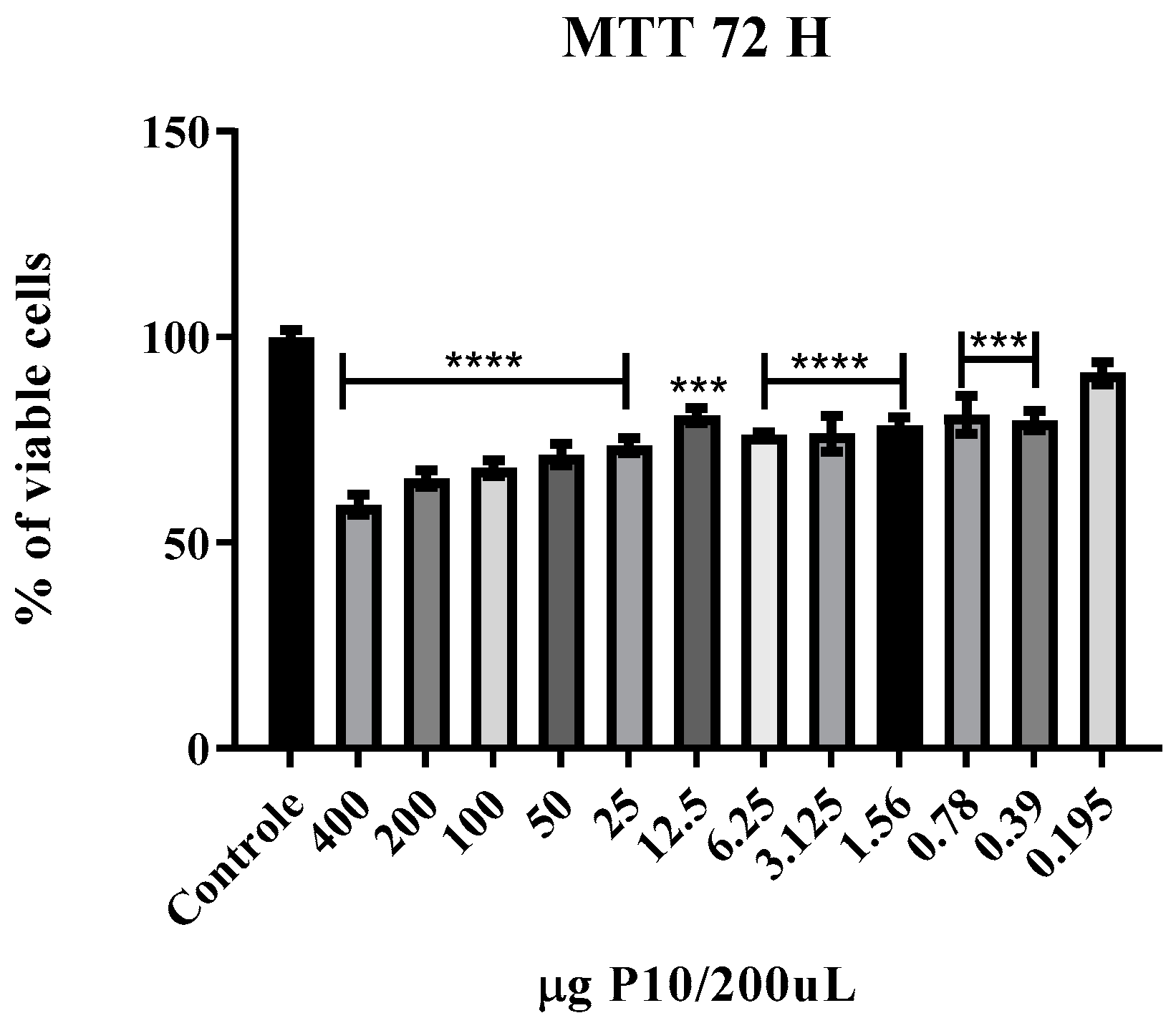
2.3. Nanoparticles Safety Assays
2.3.1. Hemolysis
2.3.2. Cell Viability
2.4. In Vivo P10-Nanoparticles Vaccine Evaluation PBS
2.4.1. Yeast
2.4.2. Animals
2.4.3. Intratracheal Infection
2.4.4. Immunization and Treatment
2.4.5. Evaluation of Treatment
2.4.6. Cytokine Production Evaluation
2.4.7. Lung Histology
2.5. Statistical Analysis
3. Results
3.1. P10-Based Chitosan Nanoparticles Vaccine
3.2. Cytotoxicity
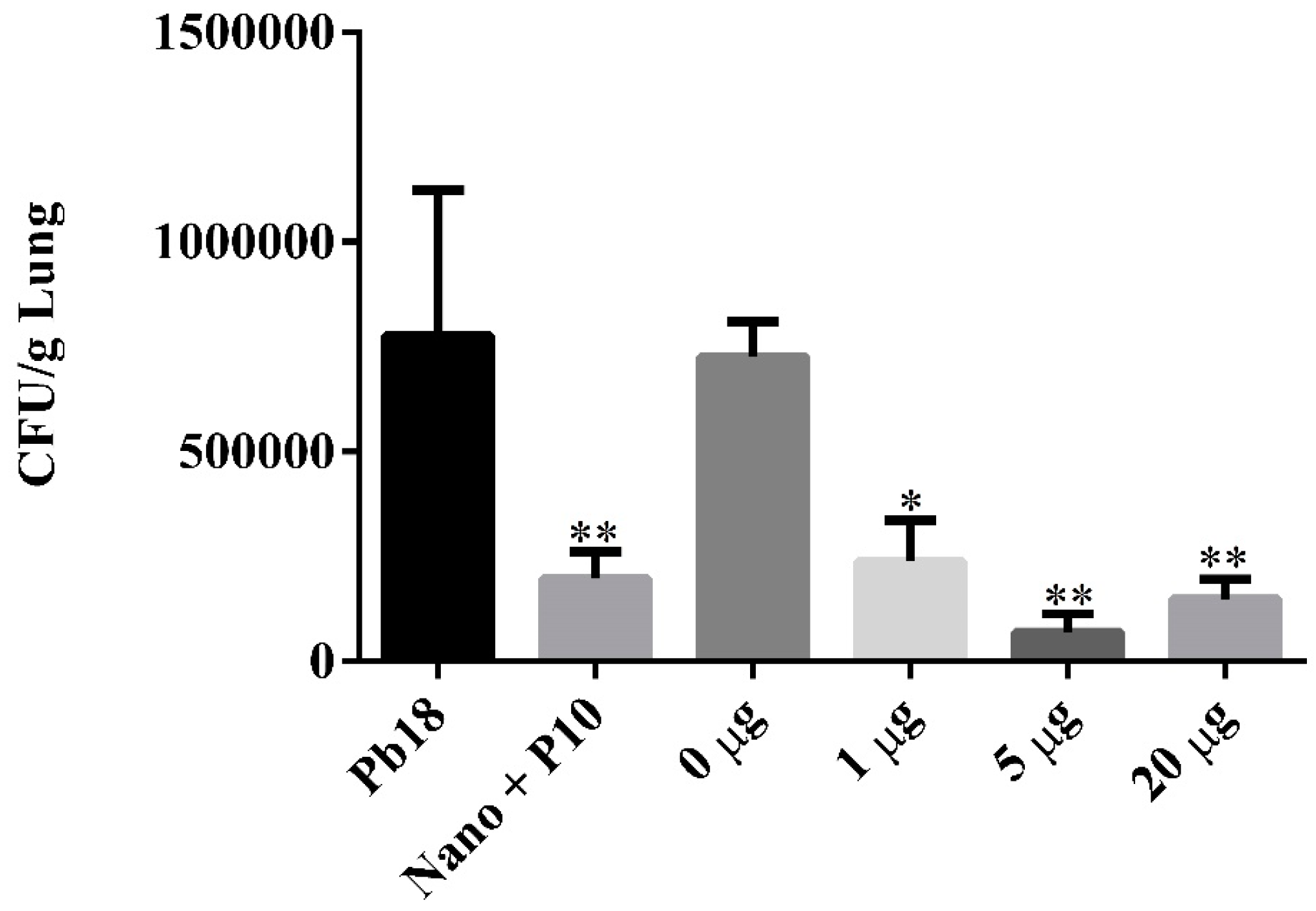
3.3. In Vivo Antifungal Efficacy of P10-Complexed Chitosan Nanoparticles
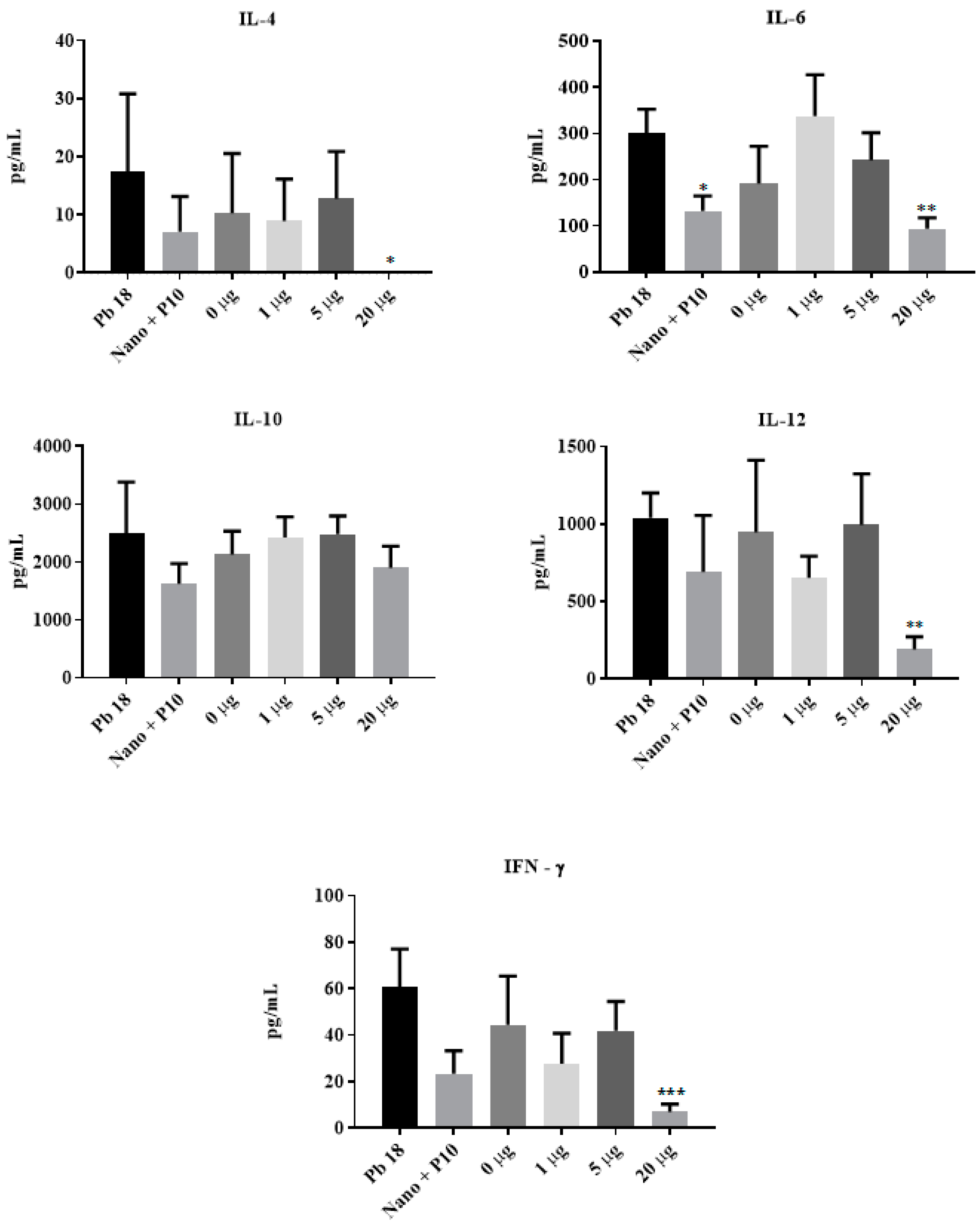
3.4. Cytokine Production Induced by P10-Complexed Chitosan Nanoparticles
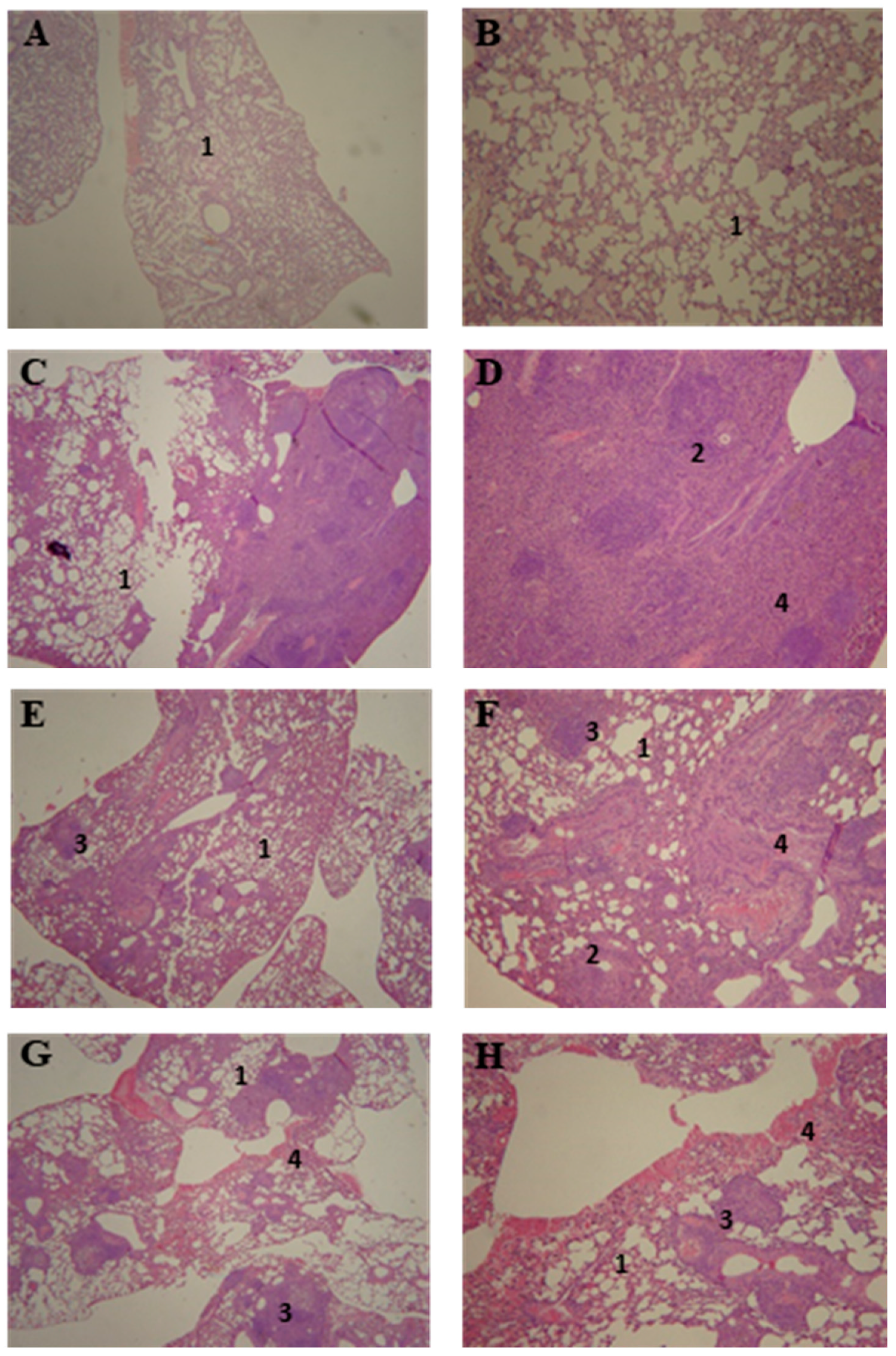
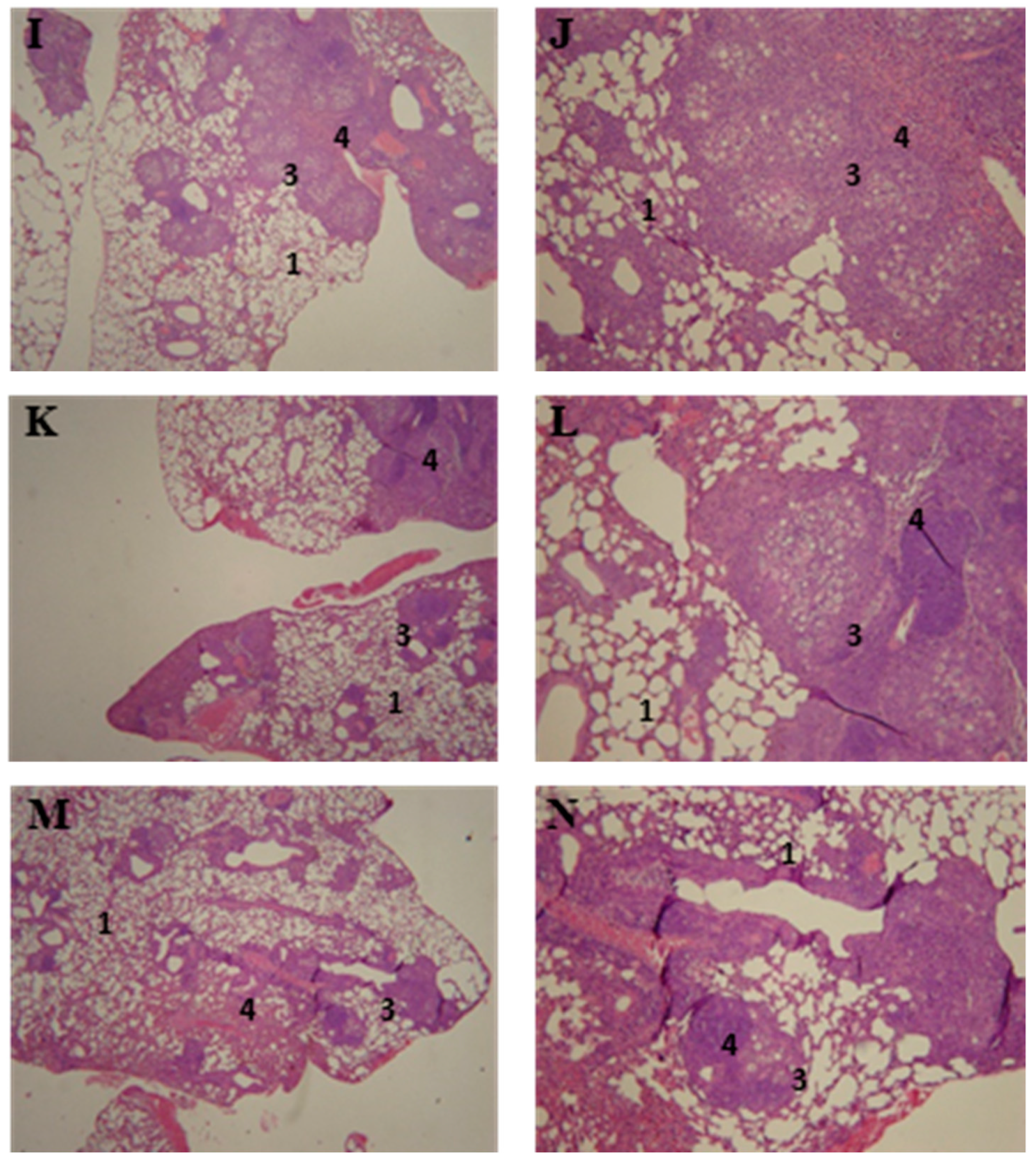
3.5. Lung Histopathology of P10-Complexed Nanoparticles Treated Animals
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Martinez, R. Epidemiology of Paracoccidioidomycosis. Rev. Inst. Med. Trop. São Paulo 2015, 57 (Suppl. 1), 11–20. [Google Scholar] [CrossRef] [Green Version]
- Martinez, R. New Trends in Paracoccidioidomycosis Epidemiology. J. Fungi 2017, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Shikanai-Yasuda, M.A.; Mendes, R.P.; Colombo, A.L.; de Queiroz-Telles, F.; Kono, A.S.G.; Paniago, A.M.M.; Nathan, A.; do Valle, A.C.F.; Bagagli, E.; Benard, G.; et al. Brazilian guidelines for the clinical management of paracoccidioidomycosis. Rev. Soc. Bras. Med. Trop. 2017, 50, 715–740. [Google Scholar] [CrossRef]
- Shikanai-yasuda, M.A.; Queiroz, F.D.; Filho, T.; Mendes, R.P.; Colombo, A.L.; Moretti, M.L.; Consultores, D. Consenso em paracoccidioidomicose Guideliness in paracoccidioidomycosis. Rev. Soc. Bras. Med. Trop. 2006, 39, 297–310. [Google Scholar] [CrossRef] [Green Version]
- Restrepo, A.; Gómez, B.L.; Tobón, A. Paracoccidioidomycosis: Latin America’s own fungal disorder. Curr. Fungal Infect. Rep. 2012, 6, 303–311. [Google Scholar] [CrossRef]
- Bocca, A.L.; Amaral, A.C.; Teixeira, M.M.; Sato, P.K.; Sato, P.; Shikanai-Yasuda, M.A.; Soares Felipe, M.S. Paracoccidioidomycosis: Eco-epidemiology, taxonomy and clinical and therapeutic issues. Future Microbiol. 2013, 8, 1177–1191. [Google Scholar] [CrossRef] [PubMed]
- Borges, S.R.C.; da Silva, G.M.S.; da Costa Chambela, M.; de Vasconcellos De, C.R.O.; Costa, R.L.B.; Wanke, B.; Do Valle, A.C.F. Itraconazole vs. trimethoprim-sulfamethoxazole: A comparative cohort study of 200 patients with paracoccidioidomycosis. Med. Mycol. 2014, 52, 303–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaral, A.C.; Bocca, A.L.A.L.; Ribeiro, A.M.; Nunes, J.; Peixoto, D.L.G.G.; Simioni, A.R.; Primo, F.L.; Lacava, Z.G.M.M.; Bentes, R.; Titze-de-Almeida, R.; et al. Amphotericin B in poly(lactic-co-glycolic acid) (PLGA) and dimercaptosuccinic acid (DMSA) nanoparticles against paracoccidioidomycosis. J. Antimicrob. Chemother. 2009, 63, 526–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, A.C.O.; Nascimento, A.L.L.; de Vasconcelos, N.M.M.; Jerônimo, M.S.S.; Siqueira, I.M.M.; R-Santos, L.; Cintra, D.O.S.O.S.; Fuscaldi, L.L.L.; Pires Júnior, O.R.R.; Titze-de-Almeida, R.; et al. Activity and in vivo tracking of Amphotericin B loaded PLGA nanoparticles. Eur. J. Med. Chem. 2015, 95, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Taborda, C.P.; Urán, M.E.; Nosanchuk, J.D.; Travassos, L.R. Paracoccidioidomycosis: Challenges in the development of a vaccine against an endemic mycosis in the Americas. Rev. Inst. Med. Trop. Sao Paulo 2015, 57, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Travassos, L.R.; Taborda, C.P. Paracoccidioidomycosis vaccine. Hum. Vaccines Immunother. 2012, 8, 1450–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nascimento, E.M.; Sgarbi, B.; Cristina, V.; Costa, S.; Miranda, A.; Silva, A.; Andrade, R. De Immunization with radioattenuated yeast cells of Paracoccidioides brasiliensis induces a long lasting protection in BALB/c mice. Vaccine 2007, 25, 7893–7899. [Google Scholar] [CrossRef]
- Pinto, A.R.; Puccia, R.; Diniz, S.N.; Franco, M.F.; Travassos, R. DNA-based vaccination against murine paracoccidioidomycosis using the gp43 gene from Paracoccidioides brasiliensis. Vaccine 2000, 18, 3050–3058. [Google Scholar] [CrossRef]
- Ribeiro, A.M.; Bocca, A.L.; Amaral, A.C.; Faccioli, L.H.; Galetti, F.C.S.; Zárate-Bladés, C.R.; Figueiredo, F.; Silva, C.L.; Felipe, M.S.S. DNAhsp65 vaccination induces protection in mice against Paracoccidioides brasiliensis infection. Vaccine 2009, 27, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Taborda, C.P.; Juliano, M.A.; Puccia, R.; Franco, M.; Travassos, L.R. Mapping of the T-cell epitope in the major 43-kilodalton glycoprotein of Paracoccidioides brasiliensis which induces a Th-1 response protective against fungal infection in BALB/c mice. Infect. Immun. 1998, 66, 786–793. [Google Scholar] [CrossRef] [Green Version]
- Cisalpino, P.S.; Puccia, R.; Yamauchi, L.M.; Cano, M.I.N.; da Silveira, J.F.; Travassos, L.R. Cloning, characterization, and epitope expression of the major diagnostic antigen of Paracoccidioides brasiliensis. J. Biol. Chem. 1996, 271, 4553–4560. [Google Scholar] [CrossRef] [Green Version]
- Jannuzzi, G.P.; de Araújo Souza, N.; Françoso, K.S.; Pereira, R.H.; Santos, R.P.; Kaihami, G.H.; de Almeida, J.R.F.; Batista, W.L.; Amaral, A.C.; Maranhao, A.Q.; et al. Therapeutic treatment with scFv–PLGA nanoparticles decreases pulmonary fungal load in a murine model of paracoccidioidomycosis. Microbes Infect. 2018, 20, 48–56. [Google Scholar] [CrossRef]
- Amaral, A.C.; Marques, A.F.; Muñoz, J.E.; Bocca, A.L.; Simioni, A.R.; Tedesco, A.C.; Morais, P.C.; Travassos, L.R.; Taborda, C.P.; Felipe, M.S.S. Poly(lactic acid-glycolic acid) nanoparticles markedly improve immunological protection provided by peptide P10 against murine paracoccidioidomycosis. Br. J. Pharmacol. 2010, 159, 1126–1132. [Google Scholar] [CrossRef] [Green Version]
- Casettari, L.; Illum, L. Chitosan in nasal delivery systems for therapeutic drugs. J. Control. Release 2014, 190, 189–200. [Google Scholar] [CrossRef]
- Read, R.C.; Naylor, S.C.; Potter, C.W.; Bond, J.; Jabbal-Gill, I.; Fisher, A.; Illum, L.; Jennings, R. Effective nasal influenza vaccine delivery using chitosan. Vaccine 2005, 23, 4367–4374. [Google Scholar] [CrossRef]
- Bernstein, J.M. Mucosal Immunology of the Uper Respiratory Tract. Respiration 1992, 59, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Mygind, N.; Dahl, R. Anatomy, physiology and function of the nasal cavities in health and disease. Adv. Drug Deliv. Rev. 1998, 29, 3–12. [Google Scholar] [CrossRef]
- Calvo, P.; Remuñan-López, C.; Vila-Jato, J.L.; Alonso, M.J. Chitosan and chitosan/ethylene oxide-propylene oxide block copolymer nanoparticles as novel carriers for proteins and vaccines. Pharm. Res. 1997, 14, 1431–1436. [Google Scholar] [CrossRef] [PubMed]
- Neves, A.L.D.P.; Milioli, C.C.; Müller, L.; Riella, H.G.; Kuhnen, N.C.; Stulzer, H.K. Factorial design as tool in chitosan nanoparticles development by ionic gelation technique. Colloids Surf. A Physicochem. Eng. Asp. 2014, 445, 34–39. [Google Scholar] [CrossRef]
- Mayorga, O.; Munoz, J.E.; Travassos, L.R.; Carlos, P. The role of adjuvants in therapeutic protection against paracoccidioidomycosis after immunization with the P10 peptide. Front. Microbiol. 2012, 3, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Travassos, L.R.; Taborda, C.P. Linear epitopes of Paracoccidioides brasiliensis and other fungal agents of human systemic mycoses as vaccine candidates. Front. Immunol. 2017, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, A.M.; Souza, A.C.O.; Amaral, A.C.; Vasconcelos, N.M.; Jerônimo, M.S.; Carneiro, F.P.; Faccioli, L.H.; Felipe, M.S.S.; Silva, C.L.; Bocca, A.L. Nanobiotechnological Approaches to Delivery of DNA Vaccine Against Fungal Infection. J. Biomed. Nanotechnol. 2013, 9, 221–230. [Google Scholar] [CrossRef]
- Ribeiro, A.M.; Bocca, A.L.; Amaral, A.C.; Souza, A.C.C.O.; Faccioli, L.H.; Coelho-Castelo, A.A.M.; Figueiredo, F.; Silva, C.L.; Felipe, M.S.S. HSP65 DNA as therapeutic strategy to treat experimental paracoccidioidomycosis. Vaccine 2010, 28, 1528–1534. [Google Scholar] [CrossRef]
- Dananjaya, S.H.S.; Erandani, W.K.C.U.; Kim, C.-H.; Nikapitiya, C.; Lee, J.; De Zoysa, M. Comparative study on antifungal activities of chitosan nanoparticles and chitosan silver nano composites against Fusarium oxysporum species complex. Int. J. Biol. Macromol. 2017, 105, 478–488. [Google Scholar] [CrossRef]
- Xing, K.; Shen, X.; Zhu, X.; Ju, X.; Miao, X.; Tian, J.; Feng, Z.; Peng, X.; Jiang, J.; Qin, S. Synthesis and in vitro antifungal efficacy of oleoyl-chitosan nanoparticles against plant pathogenic fungi. Int. J. Biol. Macromol. 2016, 82, 830–836. [Google Scholar] [CrossRef]
- Souza, A.C.O.; Amaral, A.C. Antifungal Therapy for Systemic Mycosis and the Nanobiotechnology Era: Improving Efficacy, Biodistribution and Toxicity. Front. Microbiol. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, K.; Zhang, Y.; Zhang, X.; Shi, C.; Wang, X.; Wang, X.; Jin, Z.; Cui, S. Chitosan-coated poly(lactic-co-glycolic) acid nanoparticles as an efficient delivery system for Newcastle disease virus DNA vaccine. Int. J. Nanomed. 2014, 9, 4609–4619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Lubben, I.M.; Verhoef, J.C.; Borchard, G.; Junginger, H.E. Chitosan for mucosal vaccination. Adv. Drug Deliv. Rev. 2001, 52, 139–144. [Google Scholar] [CrossRef]
- Khalili, I.; Ghadimipour, R.; Sadigh Eteghad, S.; Fathi Najafi, M.; Ebrahimi, M.M.; Godsian, N.; Sefidi Heris, Y.; Khalili, M.T. Evaluation of Immune Response Against Inactivated Avian Influenza (H9N2) Vaccine, by Using Chitosan Nanoparticles. Jundishapur J. Microbiol. 2015, 8, 10–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, L.; Zhang, Z.; Lv, J.; Zhou, P.; Hu, W.; Fang, Y.; Chen, H.; Liu, X.; Shao, J.; Zhao, F.; et al. Induction of mucosal immune responses and protection of cattle against direct-contact challenge by intranasal delivery with foot-and-mouth disease virus antigen mediated by nanoparticles. Int. J. Nanomed. 2014, 9, 5603–5618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthold, A.; Cremer, K. Preparation and characterization of chitosan microspheres as drug carrier for prednisolone sodium phosphate as model for anti- inflammatory drugs. J. Control. Release 1996, 39, 17–25. [Google Scholar] [CrossRef]
- Mohammadpour, D.N.; Eskandari, R.; Zolfagharian, H.; Mohammad, M. Preparation and in vitro characterization of chitosan nanoparticles containing Mesobuthus eupeus scorpion venom as an antigen delivery system. J. Venom. Anim. Toxins Incl. Trop. Dis. 2012, 18, 44–52. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Hu, Y.; Yin, L.; Tang, C.; Yin, C. Effects of particle size and surface charge on cellular uptake and biodistribution of polymeric nanoparticles. Biomaterials 2010, 31, 3657–3666. [Google Scholar] [CrossRef]
- Masarudin, M.J.; Cutts, S.M.; Evison, B.J.; Phillips, D.R.; Pigram, P.J. Factors determining the stability, size distribution, and cellular accumulation of small, monodisperse chitosan nanoparticles as candidate vectors for anticancer drug delivery: Application to the passive encapsulation of [14C]-doxorubicin. Nanotechnol. Sci. Appl. 2015, 8, 67. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Fan, Q.; Hao, D.; Wu, J.; Ma, G.; Su, Z. Chitosan-based mucosal adjuvants: Sunrise on the ocean. Vaccine 2015, 33, 5997–6010. [Google Scholar] [CrossRef]
- Carroll, E.C.; Jin, L.; Fitzgerald, K.A.; Bowie, A.G.; Lavelle, E.C.; Oleszycka, E.; Moran, H.B.T. The Vaccine Adjuvant Chitosan Promotes Cellular Immunity via DNA Sensor cGAS-STING-Dependent Induction of Type I Interferons Article the Vaccine Adjuvant Chitosan Promotes Cellular Immunity via DNA Sensor cGAS-STING-Dependent Induction of Type I Interfero. Immunity 2016, 44, 597–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Melo, F.M.; Braga, C.J.; Pereira, F.V.; Maricato, J.T.; Origassa, C.S.; Souza, M.F.; Melo, A.C.; Silva, P.; Tomaz, S.L.; Gimenes, K.P.; et al. Anti-metastatic immunotherapy based on mucosal administration of flagellin and immunomodulatory P10. Immunol. Cell Biol. 2015, 93, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.S.; Xu, Y.L.; Zou, X.T.; Xu, Z.R. Chitosan nanoparticles act as an adjuvant to promote both Th1 and Th2 immune responses induced by ovalbumin in mice. Mar. Drugs 2011, 9, 1038–1055. [Google Scholar] [CrossRef] [PubMed]
- Riteau, N.; Sher, A. Chitosan: An Adjuvant with an Unanticipated STING. Immunity 2016, 44, 522–524. [Google Scholar] [CrossRef] [Green Version]
- Shim, S.; Soh, S.H.; Im, Y.B.; Park, H.E.; Cho, C.S.; Kim, S.; Yoo, H.S. Elicitation of Th1/Th2 related responses in mice by chitosan nanoparticles loaded with Brucella abortus malate dehydrogenase, outer membrane proteins 10 and 19. Int. J. Med. Microbiol. 2020, 310, 151362. [Google Scholar] [CrossRef]
- Muzzarelli, R. Chitins and Chitosans as Immunoadjuvants and Non-Allergenic Drug Carriers. Mar. Drugs 2010, 8, 292–312. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Empty | Complexed |
|---|---|---|
| PDI | 0.52 ± 0.06 | 0.454 ± 0.04 |
| Size | 338 ± 8 nm | 226 ± 14 nm |
| Zeta potential | 41.14 ± 1.4 mV | 19.6 ± 3.5 mV |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues Dos Santos Junior, S.; Kelley Lopes da Silva, F.; Santos Dias, L.; Oliveira Souza, A.C.; Valdemir de Araujo, M.; Buffoni Roque da Silva, L.; Travassos, L.R.; Correa Amaral, A.; P. Taborda, C. Intranasal Vaccine Using P10 Peptide Complexed within Chitosan Polymeric Nanoparticles as Experimental Therapy for Paracoccidioidomycosis in Murine Model. J. Fungi 2020, 6, 160. https://doi.org/10.3390/jof6030160
Rodrigues Dos Santos Junior S, Kelley Lopes da Silva F, Santos Dias L, Oliveira Souza AC, Valdemir de Araujo M, Buffoni Roque da Silva L, Travassos LR, Correa Amaral A, P. Taborda C. Intranasal Vaccine Using P10 Peptide Complexed within Chitosan Polymeric Nanoparticles as Experimental Therapy for Paracoccidioidomycosis in Murine Model. Journal of Fungi. 2020; 6(3):160. https://doi.org/10.3390/jof6030160
Chicago/Turabian StyleRodrigues Dos Santos Junior, Samuel, Francenya Kelley Lopes da Silva, Lucas Santos Dias, Ana Camila Oliveira Souza, Marcelo Valdemir de Araujo, Leandro Buffoni Roque da Silva, Luiz R. Travassos, Andre Correa Amaral, and Carlos P. Taborda. 2020. "Intranasal Vaccine Using P10 Peptide Complexed within Chitosan Polymeric Nanoparticles as Experimental Therapy for Paracoccidioidomycosis in Murine Model" Journal of Fungi 6, no. 3: 160. https://doi.org/10.3390/jof6030160
APA StyleRodrigues Dos Santos Junior, S., Kelley Lopes da Silva, F., Santos Dias, L., Oliveira Souza, A. C., Valdemir de Araujo, M., Buffoni Roque da Silva, L., Travassos, L. R., Correa Amaral, A., & P. Taborda, C. (2020). Intranasal Vaccine Using P10 Peptide Complexed within Chitosan Polymeric Nanoparticles as Experimental Therapy for Paracoccidioidomycosis in Murine Model. Journal of Fungi, 6(3), 160. https://doi.org/10.3390/jof6030160