Self-Inhibitory Activity of Trichoderma Soluble Metabolites and Their Antifungal Effects on Fusarium oxysporum





Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Strains and Culture Conditions
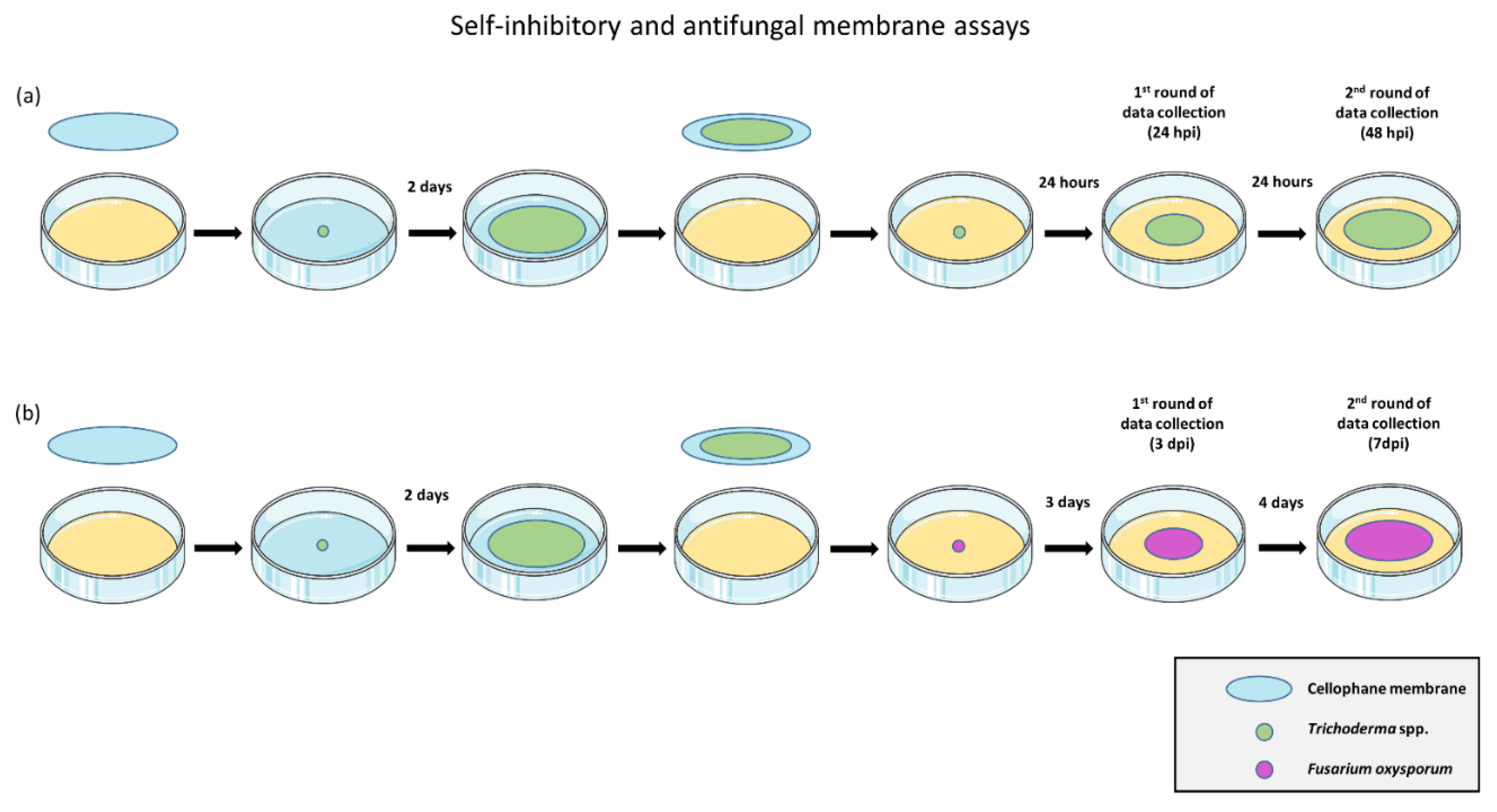
2.2. In Vitro Evaluation of Antifungal and Self-Inhibitory Activity of Soluble Metabolites Produced by Trichoderma spp.
2.3. Data Treatment and Statistical Analysis
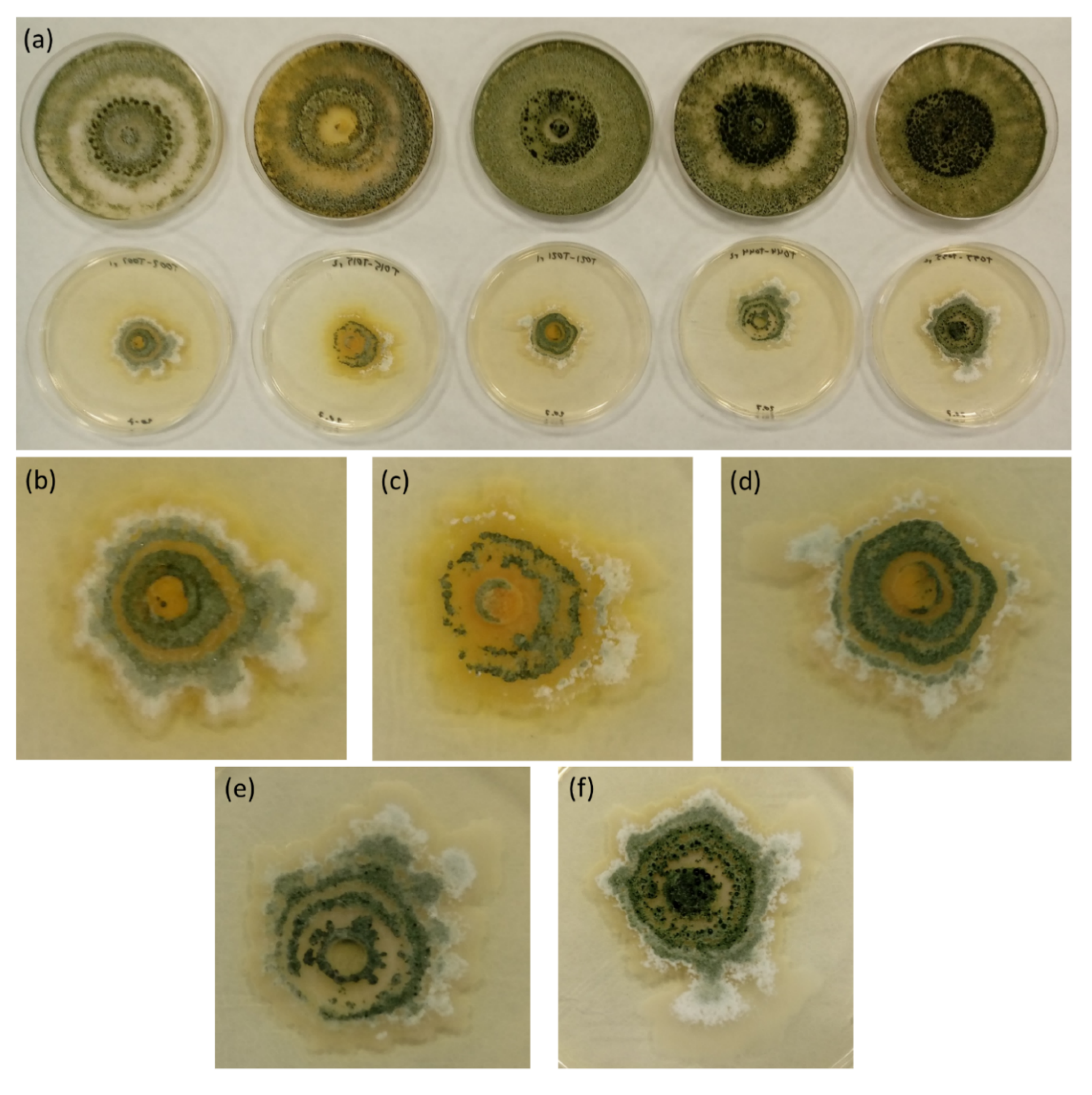
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mazzoleni, S.; Cartenì, F.; Bonanomi, G.; Senatore, M.; Termolino, P.; Giannino, F.; Incerti, G.; Rietkerk, M.; Lanzotti, V.; Chiusano, M.L. Inhibitory effects of extracellular self-DNA: A general biological process? New Phytol. 2015, 206, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Hogan, D.A. Talking to themselves: Autoregulation and quorum sensing in fungi. Eukaryot. Cell 2006, 5, 613–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillot, G.; Decourcelle, N.; Dauer, G.; Barbier, G.; Coton, E.; Delmail, D.; Mounier, J. 1-Octanol, a self-inhibitor of spore germination in Penicillium camemberti. Food Microbiol. 2016, 57, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Polyanskaya, L.M.; Tolstikhina, T.E.; Kochkina, G.A.; Ivanushkina, N.E.; Zvyagintsev, D.G. Regularities in the germination of conidia of phytopathogenic fungi. Microbiology 2004, 73, 383–388. [Google Scholar] [CrossRef]
- Chitarra, G.S.; Abee, T.; Rombouts, F.M.; Posthumus, M.A.; Dijksterhuis, J. Germination of Penicillium paneum conidia is regulated by 1-octen-3-ol, a volatile self-inhibitor. Appl. Environ. Microbiol. 2004, 70, 2823–2829. [Google Scholar] [CrossRef] [Green Version]
- Bottone, E.J.; Nagarsheth, N.; Chiu, K. Evidence of self-inhibition by filamentous fungi accounts for unidirectional hyphal growth in colonies. Can. J. Microbiol. 1998, 44, 390–393. [Google Scholar] [CrossRef]
- Mayo, S.; Gutiérrez, S.; Malmierca, M.G.; Lorenzana, A.; Campelo, M.P.; Hermosa, R.; Casquero, P.A. Influence of Rhizoctonia solani and Trichoderma spp. in growth of bean (Phaseolus vulgaris L.) and in the induction of plant defense-related genes. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Zachow, C.; Berg, C.; Müller, H.; Monk, J.; Berg, G. Endemic plants harbour specific Trichoderma communities with an exceptional potential for biocontrol of phytopathogens. J. Biotechnol. 2016, 235, 162–170. [Google Scholar] [CrossRef]
- Szczałba, M.; Kopta, T.; Gąstoł, M.; Sękara, A. Comprehensive insight into arbuscular mycorrhizal fungi, Trichoderma spp. and plant multilevel interactions with emphasis on biostimulation of horticultural crops. J. Appl. Microbiol. 2019, 127, 630–647. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Del-Val, E.; Larsen, J. Ecological functions of Trichoderma spp. and their secondary metabolites in the rhizosphere: Interactions with plants. FEMS Microbiol. Ecol. 2016, 92, fiw036. [Google Scholar] [CrossRef] [Green Version]
- Kashyap, P.L.; Rai, P.; Srivastava, A.K.; Kumar, S. Trichoderma for climate resilient agriculture. World J. Microbiol. Biotechnol. 2017, 33, 155. [Google Scholar] [CrossRef]
- de Lamo, F.J.; Takken, F.L.W. Biocontrol by Fusarium oxysporum Using Endophyte-Mediated Resistance. Front. Plant Sci. 2020, 11, 37. [Google Scholar] [CrossRef] [Green Version]
- de Borba, M.C.; Garcés-Fiallos, F.R.; Stadnik, M.J. Reactions of black bean seedlings and adult plants to infection by Fusarium oxysporum f. sp. phaseoli. Crop Prot. 2017, 96, 221–227. [Google Scholar] [CrossRef]
- Li, N.; Alfiky, A.; Wang, W.; Islam, M.; Nourollahi, K.; Liu, X.; Kang, S. Volatile Compound-Mediated Recognition and Inhibition Between Trichoderma Biocontrol Agents and Fusarium oxysporum. Front. Microbiol. 2018, 9, 1–16. [Google Scholar] [CrossRef]
- Mutawila, C.; Vinale, F.; Halleen, F.; Lorito, M.; Mostert, L. Isolation, production and in vitro effects of the major secondary metabolite produced by Trichoderma species used for the control of grapevine trunk diseases. Plant Pathol. 2016, 65, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Gotor-Vila, A.; Teixidó, N.; Di Francesco, A.; Usall, J.; Ugolini, L.; Torres, R.; Mari, M. Antifungal effect of volatile organic compounds produced by Bacillus amyloliquefaciens CPA-8 against fruit pathogen decays of cherry. Food Microbiol. 2017, 64, 219–225. [Google Scholar] [CrossRef]
- Mayo-Prieto, S.; Campelo, M.P.; Lorenzana, A.; Rodríguez-González, A.; Reinoso, B.; Gutiérrez, S.; Casquero, P.A. Antifungal activity and bean growth promotion of Trichoderma strains isolated from seed vs soil. Eur. J. Plant Pathol. 2020. [Google Scholar] [CrossRef]
- Carvalho, D.D.C.; Lobo Júnior, M.; Martins, I.; Inglis, P.W.; Mello, S.C.M. Biological control of Fusarium oxysporum f. sp. phaseoli by Trichoderma harzianum and its use for common bean seed treatment. Trop. Plant Pathol. 2014, 39, 384–391. [Google Scholar] [CrossRef] [Green Version]
- John, R.P.; Tyagi, R.D.; Prévost, D.; Brar, S.K.; Pouleur, S.; Surampalli, R.Y. Mycoparasitic Trichoderma viride as a biocontrol agent against Fusarium oxysporum f. sp. adzuki and Pythium arrhenomanes and as a growth promoter of soybean. Crop Prot. 2010, 29, 1452–1459. [Google Scholar] [CrossRef]
- Zotti, M.; De Filippis, F.; Cesarano, G.; Ercolini, D.; Tesei, G.; Allegrezza, M.; Giannino, F.; Mazzoleni, S.; Bonanomi, G. One ring to rule them all: An ecosystem engineer fungus fosters plant and microbial diversity in a Mediterranean grassland. New Phytol. 2020. [Google Scholar] [CrossRef]
- Mucha, J.; Zadworny, M.; Werner, A. Cytoskeleton and mitochondrial morphology of saprotrophs and the pathogen Heterobasidion annosum in the presence of Suillus bovinus metabolites. Mycol. Res. 2009, 113, 981–990. [Google Scholar] [CrossRef]
- Mucha, J. Changes in hyphal morphology and activity of phenoloxidases during interactions between selected ectomycorrhizal fungi and two species of Trichoderma. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2011, 100, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Carrero-Carrón, I.; Trapero-Casas, J.L.; Olivares-García, C.; Monte, E.; Hermosa, R.; Jiménez-Díaz, R.M. Trichoderma asperellum is effective for biocontrol of Verticillium wilt in olive caused by the defoliating pathotype of Verticillium dahliae. Crop Prot. 2016, 88. [Google Scholar] [CrossRef]
- Jensen, B.D.; Knorr, K.; Nicolaisen, M. In vitro competition between Fusarium graminearum and Epicoccum nigrum on media and wheat grains. Eur. J. Plant Pathol. 2016, 146, 657–670. [Google Scholar] [CrossRef]
- Taghdi, Y.; Hermosa, R.; Domínguez, S.; Rubio, M.B.; Essalmani, H.; Nicolás, C.; Monte, E. Effectiveness of composts and Trichoderma strains for control of Fusarium wilt of tomato. Phytopathol. Mediterr. 2015, 54, 232–240. [Google Scholar] [CrossRef]
- Malmierca, M.G.; Cardoza, R.E.; Alexander, N.J.; McCormick, S.P.; Hermosa, R.; Monte, E.; Gutiérrez, S. Involvement of Trichoderma trichothecenes in the biocontrol activity and induction of plant defense-related genes. Appl. Environ. Microbiol. 2012, 78, 4856–4868. [Google Scholar] [CrossRef] [Green Version]
- Kron, A.S.; Zengerer, V.; Bieri, M.; Dreyfuss, V.; Sostizzo, T.; Schmid, M.; Lutz, M.; Remus-Emsermann, M.N.P.; Pelludat, C. Pseudomonas orientalis F9 pyoverdine, safracin, and phenazine mutants remain effective antagonists against erwinia amylovora in apple flowers. Appl. Envsiron. Microbiol. 2020, 86. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Code | Trichoderma Species | Crop | Source | Municipality | Area |
|---|---|---|---|---|---|
| T002 | T. harzianum | Riñón menudo (bean) | seed | Moscas del Páramo | El Páramo |
| T007 | T. harzianum | Pinta (bean) | seed | Sueros de Cepeda | Astorga |
| T008 | T. citrinoviride | Pinta (bean) | seed | Fresno de la Vega | Esla-Campos |
| T015 | T. harzianum | Riñón menudo (bean) | seed | Veguellina de Fondo | El Páramo |
| T021 | T. harzianum | Pinta (bean) | seed | Altobar de la Encomienda | El Páramo |
| T028 | T. velutinum | Riñón (bean) | soil | Otero de Escarpizo | Astorga |
| T044 | T. harzianum | Riñón (bean) | soil | Javares de los Oteros | Esla-Campos |
| T050 | T. harzianum | Canela (bean) | soil | Bercianos del Páramo | El Páramo |
| T055 | T. harzianum | Sugarbeet | soil | La Milla del Páramo | El Páramo |
| T057 | T. gamsii | Sugarbeet | soil | La Milla del Páramo | El Páramo |
| First Round of Data Collection | Second Round of Data Collection | ||||||||
|---|---|---|---|---|---|---|---|---|---|
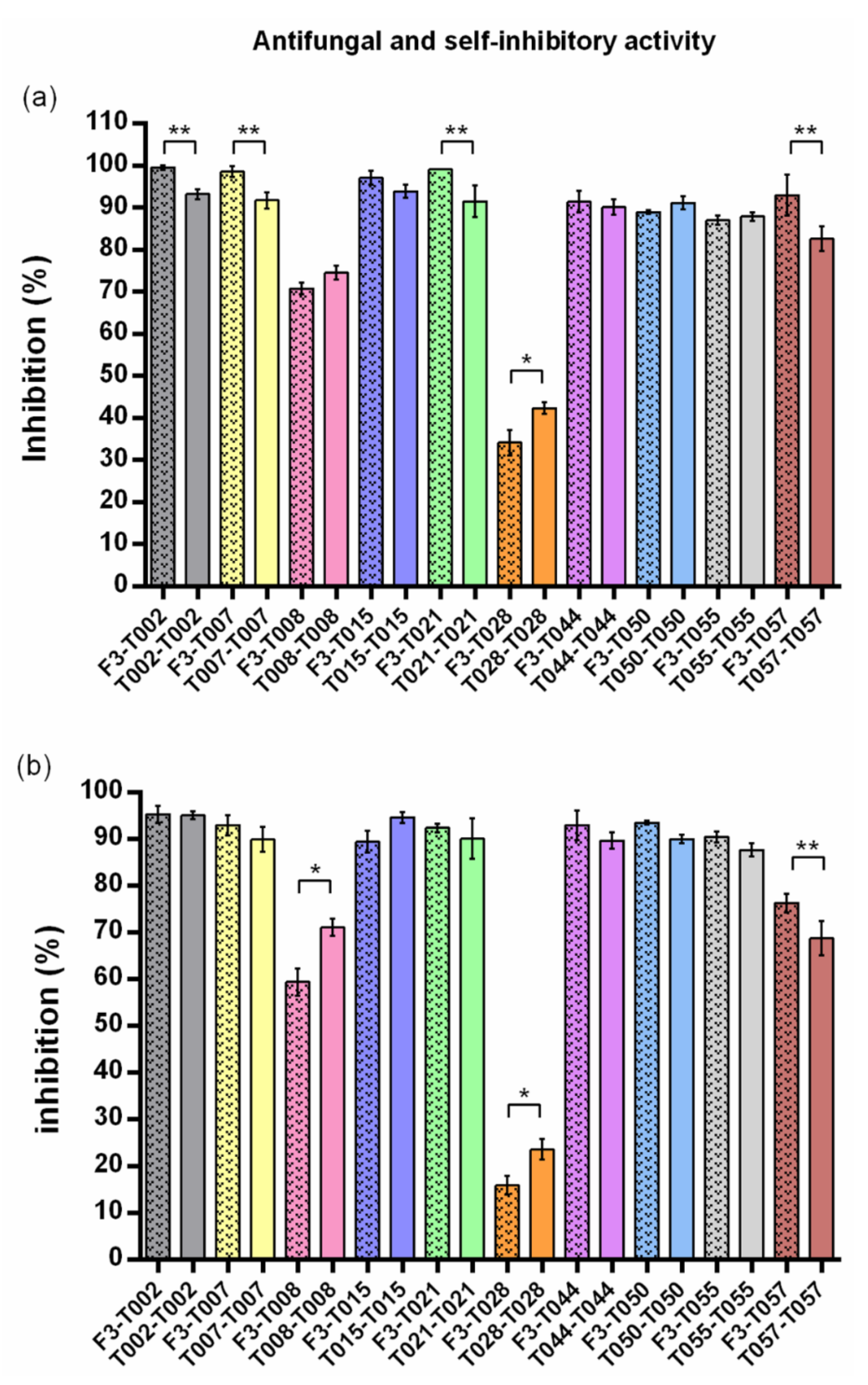
| Code | Trichoderma | Self-Inhibition (% ± SD) | Statistics 1 | F3 Inhibition (% ± SD) | Statistics 2 | Self-Inhibition (% ± SD) | Statistics 1 | F3 Inhibition (% ± SD) | Statistics 2 |
| T002 | T. harzianum | 93.18 ± 1.21 | A | 99.51 ± 0.56 | a | 95.04 ± 0.87 | A | 95.25 ± 1.89 | a |
| T007 | T. harzianum | 91.68 ± 1.89 | A,B | 98.54 ± 1.26 | a | 89.87 ± 2.67 | A,B | 92.93 ± 2.16 | a,b |
| T008 | T. citrinoviride | 74.51 ± 1.66 | D | 70.73 ± 1.38 | e | 71.02 ± 1.85 | C | 59.38 ± 2.93 | d |
| T015 | T. harzianum | 93.86 ± 1.60 | A | 97.07 ± 1.59 | a,b | 94.62 ± 1.21 | A | 89.40 ± 2.28 | b |
| T021 | T. harzianum | 91.43 ± 3.77 | A,B | 99.02 ± 0.00 | a | 90.08 ± 4.25 | A,B | 92.38 ± 0.91 | a,b |
| T028 | T. velutinum | 42.29 ± 1.40 | E | 34.15 ± 3.03 | f | 23.57 ± 2.12 | D | 15.89 ± 1.96 | e |
| T044 | T. harzianum | 90.09 ± 1.79 | A,B | 91.35 ± 2.55 | c,d | 89.66 ± 1.71 | A,B | 92.93 ± 3.22 | a,b |
| T050 | T. harzianum | 91.11 ± 1.48 | A,B | 88.91 ± 0.51 | c,d | 89.98 ± 0.94 | A,B | 93.46 ± 0.42 | a,b |
| T055 | T. harzianum | 87.86 ± 0.97 | B | 86.92 ± 1.12 | d | 87.66 ± 1.35 | B | 90.40 ± 1.11 | a,b |
| T057 | T. gamsii | 82.53 ± 2.91 | C | 92.90 ± 4.86 | b,c | 68.74 ± 3.65 | C | 76.27 ± 2.02 | c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Álvarez-García, S.; Mayo-Prieto, S.; Gutiérrez, S.; Casquero, P.A. Self-Inhibitory Activity of Trichoderma Soluble Metabolites and Their Antifungal Effects on Fusarium oxysporum. J. Fungi 2020, 6, 176. https://doi.org/10.3390/jof6030176
Álvarez-García S, Mayo-Prieto S, Gutiérrez S, Casquero PA. Self-Inhibitory Activity of Trichoderma Soluble Metabolites and Their Antifungal Effects on Fusarium oxysporum. Journal of Fungi. 2020; 6(3):176. https://doi.org/10.3390/jof6030176
Chicago/Turabian StyleÁlvarez-García, Samuel, Sara Mayo-Prieto, Santiago Gutiérrez, and Pedro Antonio Casquero. 2020. "Self-Inhibitory Activity of Trichoderma Soluble Metabolites and Their Antifungal Effects on Fusarium oxysporum" Journal of Fungi 6, no. 3: 176. https://doi.org/10.3390/jof6030176
APA StyleÁlvarez-García, S., Mayo-Prieto, S., Gutiérrez, S., & Casquero, P. A. (2020). Self-Inhibitory Activity of Trichoderma Soluble Metabolites and Their Antifungal Effects on Fusarium oxysporum. Journal of Fungi, 6(3), 176. https://doi.org/10.3390/jof6030176