Influences of the Culturing Media in the Virulence and Cell Wall of Sporothrix schenckii, Sporothrix brasiliensis, and Sporothrix globosa

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Galleria mellonella Survival Assays
2.3. Ethics Statement
2.4. Analysis of Hemocyte Levels, Phenoloxidase, and Lactate Dehydrogenase Activities
2.5. Cell Wall Analysis
2.6. Analysis of Uptake by Hemocytes and Fungal-Secreted Protease and Lipase/Esterase Activities
2.7. Statistical Analysis
3. Results
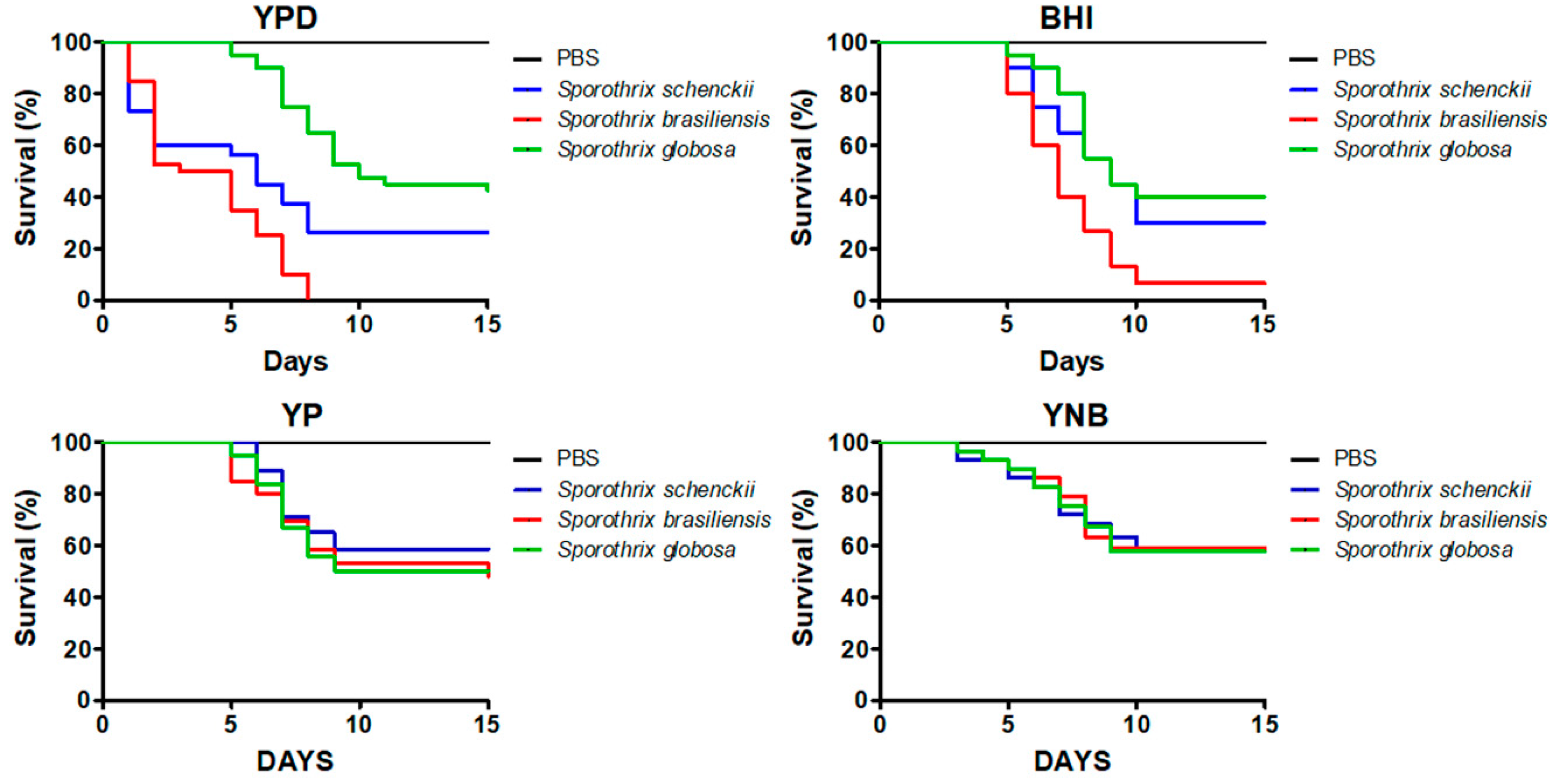
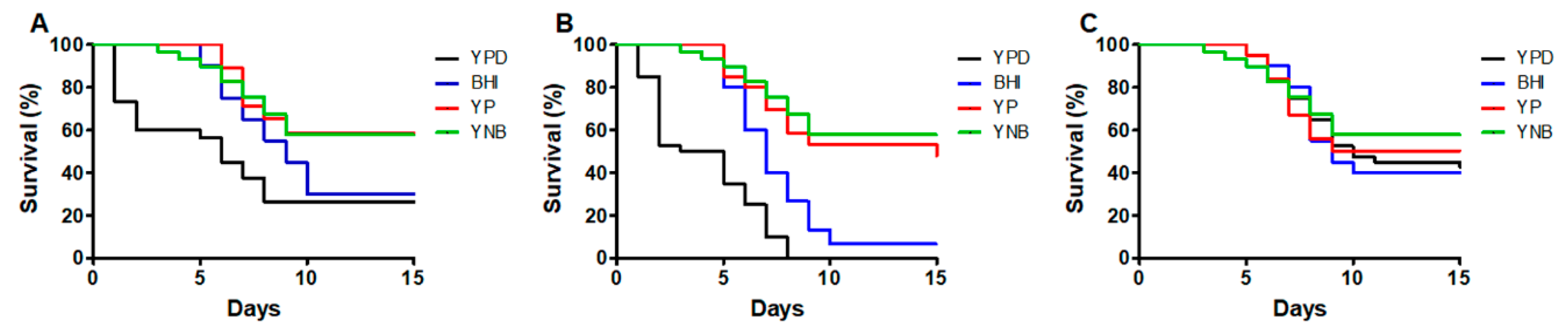
3.1. The Culture Medium Affects the Sporothrix spp. Ability to Kill Galleria mellonella Larvae
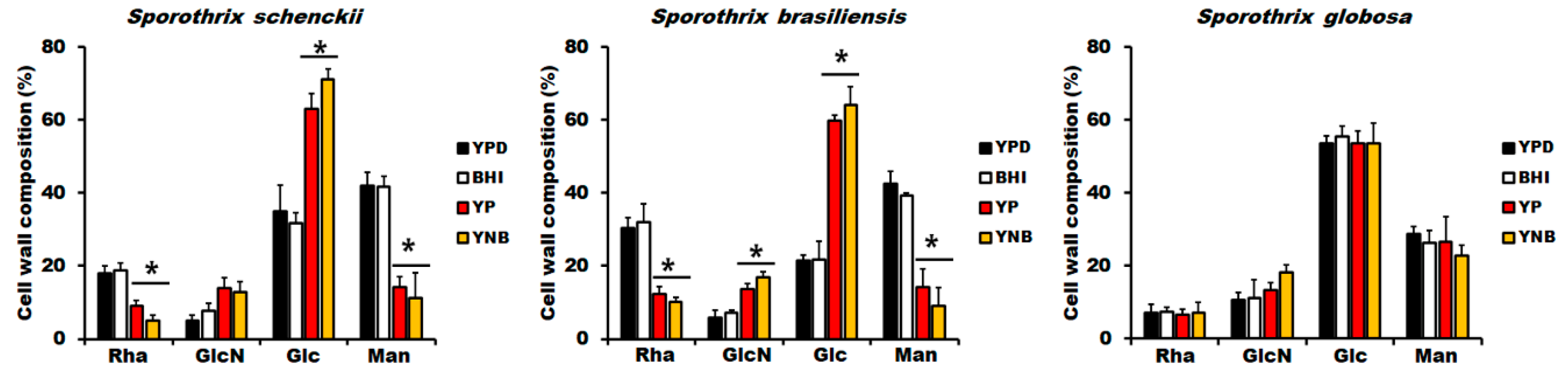
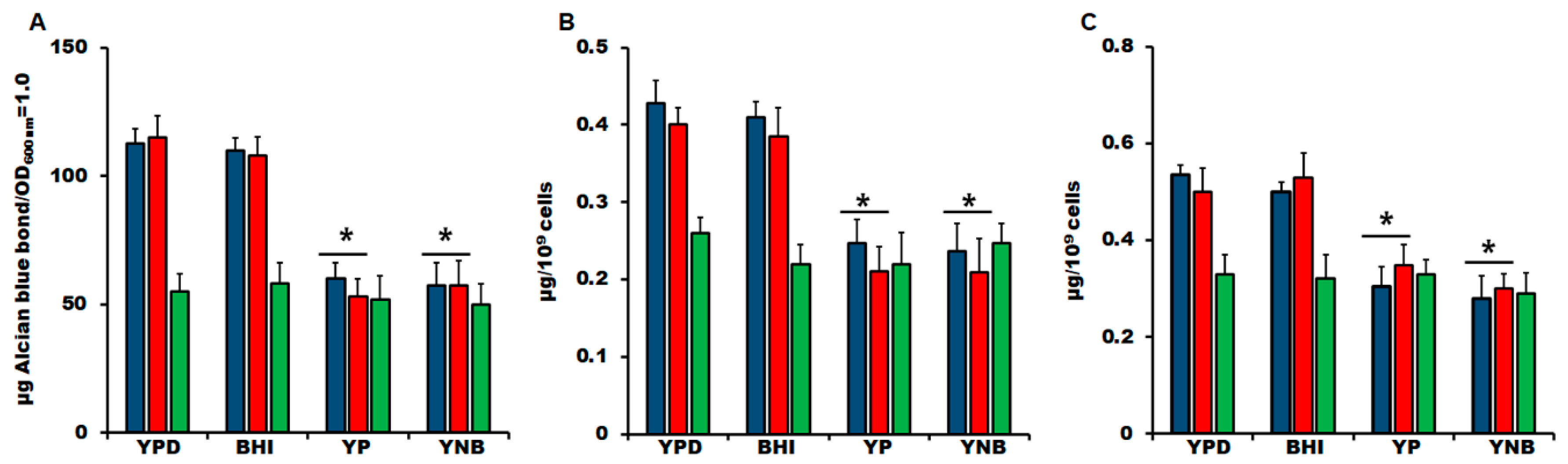
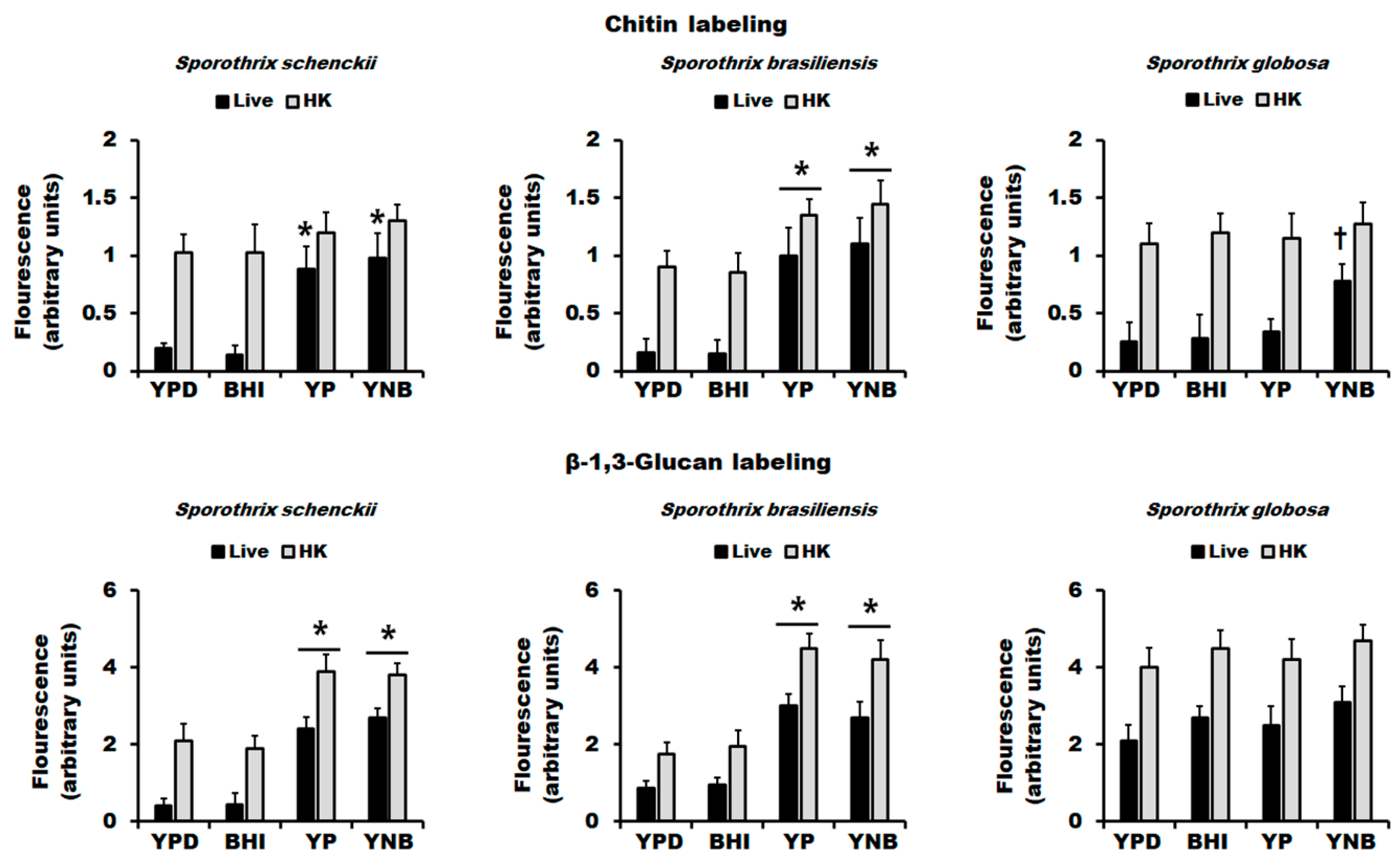
3.2. The Culture Medium Affects Sporothrix spp. Cell Wall Composition
3.3. Analysis of Secreted Protease and Lipase/Esterase Activity and Uptake by Hemocytes of Sporothrix schenckii, Sporothrix brasiliensis, and Sporothrix globosa Grown in Different Culture Media
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, M.C.; Gurr, S.J.; Cuomo, C.A.; Blehert, D.S.; Jin, H.; Stukenbrock, E.H.; Stajich, J.E.; Kahmann, R.; Boone, C.; Denning, D.W.; et al. Threats posed by the fungal kingdom to humans, wildlife, and agriculture. mBio 2020, 11, e00449-20. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Bezerra, L.M.; Mora-Montes, H.M.; Zhang, Y.; Nino-Vega, G.; Rodrigues, A.M.; de Camargo, Z.P.; de Hoog, S. Sporotrichosis between 1898 and 2017: The evolution of knowledge on a changeable disease and on emerging etiological agents. Med. Mycol. 2018, 56, 126–143. [Google Scholar] [CrossRef] [PubMed]
- Mora-Montes, H.M.; Dantas Ada, S.; Trujillo-Esquivel, E.; de Souza Baptista, A.R.; Lopes-Bezerra, L.M. Current progress in the biology of members of the Sporothrix schenckii complex following the genomic era. FEMS Yeast Res. 2015, 15, fov065. [Google Scholar] [CrossRef] [Green Version]
- López-Romero, E.; Reyes-Montes Mdel, R.; Pérez-Torres, A.; Ruiz-Baca, E.; Villagómez-Castro, J.C.; Mora-Montes, H.M.; Flores-Carreón, A.; Toriello, C. Sporothrix schenckii complex and sporotrichosis, an emerging health problem. Future Microbiol. 2011, 6, 85–102. [Google Scholar] [CrossRef]
- de Beer, Z.W.; Duong, T.A.; Wingfield, M.J. The divorce of Sporothrix and Ophiostoma: Solution to a problematic relationship. Stud. Mycol. 2016, 83, 165–191. [Google Scholar] [CrossRef] [Green Version]
- Arenas, R.; Sánchez-Cardenas, C.D.; Ramirez-Hobak, L.; Ruíz Arriaga, L.F.; Vega Memije, M.E. Sporotrichosis: From KOH to molecular biology. J. Fungi 2018, 4, 62. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, A.; Bonifaz, A.; Gutierrez-Galhardo, M.C.; Mochizuki, T.; Li, S. Global epidemiology of sporotrichosis. Med. Mycol. 2015, 53, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, A.M.; de Hoog, G.S.; de Camargo, Z.P. Sporothrix species causing outbreaks in animals and humans driven by animal-animal transmission. PLoS Pathog. 2016, 12, e1005638. [Google Scholar] [CrossRef]
- Etchecopaz, A.N.; Lanza, N.; Toscanini, M.A.; Devoto, T.B.; Pola, S.J.; Daneri, G.L.; Iovannitti, C.A.; Cuestas, M.L. Sporotrichosis caused by Sporothrix brasiliensis in Argentina: Case report, molecular identification and in vitro susceptibility pattern to antifungal drugs. J. Mycol. Med. 2020, 30, 100908. [Google Scholar] [CrossRef]
- Gremião, I.D.; Miranda, L.H.; Reis, E.G.; Rodrigues, A.M.; Pereira, S.A. Zoonotic epidemic of sporotrichosis: Cat to human transmission. PLoS Pathog. 2017, 13, e1006077. [Google Scholar] [CrossRef] [PubMed]
- Moussa, T.A.A.; Kadasa, N.M.S.; Al Zahrani, H.S.; Ahmed, S.A.; Feng, P.; Gerrits van den Ende, A.H.G.; Zhang, Y.; Kano, R.; Li, F.; Li, S.; et al. Origin and distribution of Sporothrix globosa causing sapronoses in Asia. J. Med. Microbiol. 2017, 66, 560–569. [Google Scholar] [CrossRef] [PubMed]
- Camacho, E.; León-Navarro, I.; Rodríguez-Brito, S.; Mendoza, M.; Niño-Vega, G.A. Molecular epidemiology of human sporotrichosis in Venezuela reveals high frequency of Sporothrix globosa. BMC Infect. Dis. 2015, 15, 94. [Google Scholar] [CrossRef]
- Martinez-Alvarez, J.A.; Perez-Garcia, L.A.; Mellado-Mojica, E.; Lopez, M.G.; Martinez-Duncker, I.; Lopes-Bezerra, L.M.; Mora-Montes, H.M. Sporothrix schenckii sensu stricto and Sporothrix brasiliensis are differentially recognized by human peripheral blood mononuclear cells. Front. Microbiol. 2017, 8, 843. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, M.M.; de Almeida, L.G.; Kubitschek-Barreira, P.; Alves, F.L.; Kioshima, E.S.; Abadio, A.K.; Fernandes, L.; Derengowski, L.S.; Ferreira, K.S.; Souza, R.C.; et al. Comparative genomics of the major fungal agents of human and animal sporotrichosis: Sporothrix schenckii and Sporothrix brasiliensis. BMC Genom. 2014, 15, 943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva-Bailão, M.G.; de Sousa Lima, P.; Evangelista de Oliveira, M.M.; Cardoso de Oliveira, L.; Almeida-Paes, R.; Borges, C.L.; Bailão, A.M.; Guedes Coelho, A.S.; Maria de Almeida Soares, C.; Zancopé-Oliveira, R.M. Comparative proteomics in the three major human pathogenic species of the genus Sporothrix. Microbes Infect. 2020. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, G.F.; dos Santos, P.O.; Rodrigues, A.M.; Sasaki, A.A.; Burger, E.; de Camargo, Z.P. Characterization of virulence profile, protein secretion and immunogenicity of different Sporothrix schenckii sensu stricto isolates compared with S. globosa and S. brasiliensis species. Virulence 2013, 4, 241–249. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Gao, W.; Giosa, D.; Criseo, G.; Zhang, J.; He, T.; Huang, X.; Sun, J.; Sun, Y.; Huang, J.; et al. Whole-genome sequencing and in silico analysis of two strains of Sporothrix globosa. Genome Biol. Evol. 2016, 8, 3292–3296. [Google Scholar] [CrossRef] [Green Version]
- Arrillaga-Moncrieff, I.; Capilla, J.; Mayayo, E.; Marimon, R.; Mariné, M.; Gené, J.; Cano, J.; Guarro, J. Different virulence levels of the species of Sporothrix in a murine model. Clin. Microbiol. Infect. 2009, 15, 651–655. [Google Scholar] [CrossRef] [Green Version]
- Marimon, R.; Serena, C.; Gené, J.; Cano, J.; Guarro, J. In vitro antifungal susceptibilities of five species of Sporothrix. Antimicrob. Agents Chemother. 2008, 52, 732–734. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, D.C.; Lopes, P.G.; Spader, T.B.; Mahl, C.D.; Tronco-Alves, G.R.; Lara, V.M.; Santurio, J.M.; Alves, S.H. Antifungal susceptibilities of Sporothrix albicans, S. brasiliensis, and S. luriei of the S. schenckii complex identified in Brazil. J. Clin. Microbiol. 2011, 49, 3047–3049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, M.M.E.; Almeida-Paes, R.; Corrêa-Moreira, D.; Borba, C.d.M.; Menezes, R.C.; Freitas, D.F.S.; do Valle, A.C.F.; Schubach, A.d.O.; Barros, M.B.d.L.; Nosanchuk, J.D.; et al. A case of sporotrichosis caused by different Sporothrix brasiliensis strains: Mycological, molecular, and virulence analyses. Mem. Inst. Oswaldo Cruz 2019, 114, e190260. [Google Scholar] [CrossRef] [PubMed]
- Batista-Duharte, A.; Téllez-Martínez, D.; Aparecida Jellmayer, J.; Leandro Portuondo Fuentes, D.; Campos Polesi, M.; Martins Baviera, A.; Zeppone Carlos, I. Repeated exposition to mercury (ii) chloride enhances susceptibility to S. schenckii sensu stricto infection in mice. J. Fingi 2018, 4, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Almeida, J.R.F.; Jannuzzi, G.P.; Kaihami, G.H.; Breda, L.C.D.; Ferreira, K.S.; de Almeida, S.R. An immunoproteomic approach revealing peptides from Sporothrix brasiliensis that induce a cellular immune response in subcutaneous sporotrichosis. Sci. Rep. 2018, 8, 4192. [Google Scholar] [CrossRef] [PubMed]
- Della Terra, P.P.; Rodrigues, A.M.; Fernandes, G.F.; Nishikaku, A.S.; Burger, E.; de Camargo, Z.P. Exploring virulence and immunogenicity in the emerging pathogen Sporothrix brasiliensis. PLoS Negl. Trop. Dis. 2017, 11, e0005903. [Google Scholar] [CrossRef]
- Cruz Choappa, R.; Pérez Gaete, S.; Rodríguez Badilla, V.; Vieille Oyarzo, P.; Opazo Sanchez, H. Virulence of Sporothrix globosa in murine models. Rev. Argent. Microbiol. 2016, 48, 196–199. [Google Scholar] [CrossRef] [Green Version]
- Castro, R.A.; Kubitschek-Barreira, P.H.; Teixeira, P.A.; Sanches, G.F.; Teixeira, M.M.; Quintella, L.P.; Almeida, S.R.; Costa, R.O.; Camargo, Z.P.; Felipe, M.S.; et al. Differences in cell morphometry, cell wall topography and gp70 expression correlate with the virulence of Sporothrix brasiliensis clinical isolates. PLoS ONE 2013, 8, e75656. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, K.S.S.; Mathews, H.L.; Bezerra, L.M.L. Differences in virulence of Sporothrix schenckii conidia related to culture conditions and cell-wall components. J. Med Microbiol. 1999, 48, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Almeida-Paes, R.; de Oliveira, L.C.; Oliveira, M.M.E.; Gutierrez-Galhardo, M.C.; Nosanchuk, J.D.; Zancopé-Oliveira, R.M. Phenotypic characteristics associated with virulence of clinical isolates from the Sporothrix complex. Biomed. Res. Int. 2015, 2015, 212308. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.; Xiao, T.; Lin, J.; Wang, Y.; Chen, H.D. Relationships among genotypes, virulence and clinical forms of Sporothrix schenckii infection. Clin. Microbiol. Infect. 2006, 12, 1077–1081. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, P.A.; de Castro, R.A.; Nascimento, R.C.; Tronchin, G.; Torres, A.P.; Lazera, M.; de Almeida, S.R.; Bouchara, J.P.; Loureiro y Penha, C.V.; Lopes-Bezerra, L.M. Cell surface expression of adhesins for fibronectin correlates with virulence in Sporothrix schenckii. Microbiology 2009, 155, 3730–3738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clavijo-Giraldo, D.M.; Matinez-Alvarez, J.A.; Lopes-Bezerra, L.M.; Ponce-Noyola, P.; Franco, B.; Almeida, R.S.; Mora-Montes, H.M. Analysis of Sporothrix schenckii sensu stricto and Sporothrix brasiliensis virulence in Galleria mellonella. J. Microbiol. Methods 2016, 122, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Lozoya-Perez, N.E.; Casas-Flores, S.; de Almeida, J.R.F.; Martinez-Alvarez, J.A.; Lopez-Ramirez, L.A.; Jannuzzi, G.P.; Trujillo-Esquivel, E.; Estrada-Mata, E.; Almeida, S.R.; Franco, B.; et al. Silencing of OCH1 unveils the role of Sporothrix schenckii N-linked glycans during the host-fungus interaction. Infect. Drug Resist. 2019, 12, 67–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozoya-Perez, N.E.; Casas-Flores, S.; Martinez-Alvarez, J.A.; Lopez-Ramirez, L.A.; Lopes-Bezerra, L.M.; Franco, B.; Mora-Montes, H.M. Generation of Sporothrix schenckii mutants expressing the green fluorescent protein suitable for the study of host-fungus interactions. Fungal Biol. 2018, 122, 1023–1030. [Google Scholar] [CrossRef]
- Desalermos, A.; Fuchs, B.B.; Mylonakis, E. Selecting an invertebrate model host for the study of fungal pathogenesis. PLoS Pathog. 2012, 8, e1002451. [Google Scholar] [CrossRef] [Green Version]
- de Souza, P.C.; Morey, A.T.; Castanheira, G.M.; Bocate, K.P.; Panagio, L.A.; Ito, F.A.; Furlaneto, M.C.; Yamada-Ogatta, S.F.; Costa, I.N.; Mora-Montes, H.M.; et al. Tenebrio molitor (Coleoptera: Tenebrionidae) as an alternative host to study fungal infections. J. Microbiol. Methods 2015, 118, 182–186. [Google Scholar] [CrossRef] [Green Version]
- Trevijano-Contador, N.; Zaragoza, O. Immune response of Galleria mellonella against human fungal pathogens. J. Fungi 2018, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Bergin, D.; Brennan, M.; Kavanagh, K. Fluctuations in haemocyte density and microbial load may be used as indicators of fungal pathogenicity in larvae of Galleria mellonella. Microb. Infect. 2003, 5, 1389–1395. [Google Scholar] [CrossRef] [Green Version]
- Mesa-Arango, A.C.; Forastiero, A.; Bernal-Martinez, L.; Cuenca-Estrella, M.; Mellado, E.; Zaragoza, O. The non-mammalian host Galleria mellonella can be used to study the virulence of the fungal pathogen Candida tropicalis and the efficacy of antifungal drugs during infection by this pathogenic yeast. Med Mycol. 2013, 51, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Brito, M.M.; Conceição-Silva, F.; Morgado, F.N.; Raibolt, P.S.; Schubach, A.; Schubach, T.P.; Schäffer, G.M.; Borba, C.M. Comparison of virulence of different Sporothrix schenckii clinical isolates using experimental murine model. Med. Mycol. 2007, 45, 721–729. [Google Scholar] [CrossRef] [Green Version]
- Odds, F.C.; Van Nuffel, L.; Gow, N.A.R. Survival in experimental Candida albicans infections depends on inoculum growth conditions as well as animal host. Microbiology 2000, 146 Pt 8, 1881–1889. [Google Scholar] [CrossRef] [Green Version]
- Gilmore, S.A.; Naseem, S.; Konopka, J.B.; Sil, A. N-acetylglucosamine (GlcNAc) triggers a rapid, temperature-responsive morphogenetic program in thermally dimorphic fungi. PLoS Genet. 2013, 9, e1003799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ene, I.V.; Adya, A.K.; Wehmeier, S.; Brand, A.C.; MacCallum, D.M.; Gow, N.A.R.; Brown, A.J.P. Host carbon sources modulate cell wall architecture, drug resistance and virulence in a fungal pathogen. Cell Microbiol. 2012, 14, 1319–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ene, I.V.; Cheng, S.-C.; Netea, M.G.; Brown, A.J.P. Growth of Candida albicans cells on the physiologically relevant carbon source lactate affects their recognition and phagocytosis by immune cells. Infect. Immun. 2013, 81, 238–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ene, I.V.; Heilmann, C.J.; Sorgo, A.G.; Walker, L.A.; de Koster, C.G.; Munro, C.A.; Klis, F.M.; Brown, A.J.P. Carbon source-induced reprogramming of the cell wall proteome and secretome modulates the adherence and drug resistance of the fungal pathogen Candida albicans. Proteomics 2012, 12, 3164–3179. [Google Scholar] [CrossRef] [Green Version]
- Lok, B.; Adam, M.A.A.; Kamal, L.Z.M.; Chukwudi, N.A.; Sandai, R.; Sandai, D. The assimilation of different carbon sources in Candida albicans: Fitness and pathogenicity. Med. Mycol. 2020. [Google Scholar] [CrossRef]
- Madrid, H.; Cano, J.; Gene, J.; Bonifaz, A.; Toriello, C.; Guarro, J. Sporothrix globosa, a pathogenic fungus with widespread geographical distribution. Rev. Iberoam. Micol. 2009, 26, 218–222. [Google Scholar] [CrossRef] [Green Version]
- Lopes-Bezerra, L.M.; Walker, L.A.; Nino-Vega, G.; Mora-Montes, H.M.; Neves, G.W.P.; Villalobos-Duno, H.; Barreto, L.; Garcia, K.; Franco, B.; Martinez-Alvarez, J.A.; et al. Cell walls of the dimorphic fungal pathogens Sporothrix schenckii and Sporothrix brasiliensis exhibit bilaminate structures and sloughing of extensive and intact layers. PLoS Negl. Trop. Dis. 2018, 12, e0006169. [Google Scholar] [CrossRef] [Green Version]
- Mora-Montes, H.M.; Robledo-Ortiz, C.I.; Gonzalez-Sanchez, L.C.; Lopez-Esparza, A.; Lopez-Romero, E.; Flores-Carreon, A. Purification and biochemical characterisation of endoplasmic reticulum alpha1,2-mannosidase from Sporothrix schenckiil. Mem. Inst. Oswaldo Cruz 2010, 105, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Carnero, L.C.; Clavijo-Giraldo, D.M.; Gomez-Gaviria, M.; Lozoya-Perez, N.E.; Tamez-Castrellon, A.K.; Lopez-Ramirez, L.A.; Mora-Montes, H.M. Early virulence predictors during the Candida species-Galleria mellonella Interaction. J. Fungi 2020, 6, 152. [Google Scholar] [CrossRef]
- Martinez-Alvarez, J.A.; Garcia-Carnero, L.C.; Kubitschek-Barreira, P.H.; Lozoya-Perez, N.E.; Belmonte-Vazquez, J.L.; de Almeida, J.R.; Gomez-Infante, A.d.J.; Curty, N.; Villagomez-Castro, J.C.; Pena-Cabrera, E.; et al. Analysis of some immunogenic properties of the recombinant Sporothrix schenckii Gp70 expressed in Escherichia coli. Future Microbiol. 2019, 14, 397–410. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Yi, Y.; Lv, Y.; Li, M.; Wang, J.; Qiu, L. The lipopolysaccharide (LPS) of Photorhabdus luminescens TT01 can elicit dose- and time-dependent immune priming in Galleria mellonella larvae. J. Invertebr. Pathol. 2015, 127, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Bidla, G.; Hauling, T.; Dushay, M.S.; Theopold, U. Activation of insect phenoloxidase after injury: Endogenous versus foreign elicitors. J. Innate Immun. 2009, 1, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Gaviria, M.; Lozoya-Perez, N.E.; Staniszewska, M.; Franco, B.; Nino-Vega, G.A.; Mora-Montes, H.M. Loss of Kex2 affects the Candida albicans cell wall and interaction with innate immune cells. J. Fungi 2020, 6, 57. [Google Scholar] [CrossRef] [PubMed]
- Mora-Montes, H.M.; Bates, S.; Netea, M.G.; Diaz-Jimenez, D.F.; Lopez-Romero, E.; Zinker, S.; Ponce-Noyola, P.; Kullberg, B.J.; Brown, A.J.; Odds, F.C.; et al. Endoplasmic reticulum alpha-glycosidases of Candida albicans are required for N glycosylation, cell wall integrity, and normal host-fungus interaction. Eukaryot. Cell 2007, 6, 2184–2193. [Google Scholar] [CrossRef] [Green Version]
- Estrada-Mata, E.; Navarro-Arias, M.J.; Perez-Garcia, L.A.; Mellado-Mojica, E.; Lopez, M.G.; Csonka, K.; Gacser, A.; Mora-Montes, H.M. Members of the Candida parapsilosis complex and Candida albicans are differentially recognized by human peripheral blood mononuclear cells. Front. Microbiol. 2016, 6, 1527. [Google Scholar] [CrossRef] [Green Version]
- Hobson, R.P.; Munro, C.A.; Bates, S.; MacCallum, D.M.; Cutler, J.E.; Heinsbroek, S.E.M.; Brown, G.D.; Odds, F.C.; Gow, N.A.R. Loss of Cell Wall Mannosylphosphate in Candida albicans Does Not Influence Macrophage Recognition. J. Biol. Chem. 2004, 279, 39628–39635. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Arias, M.J.; Defosse, T.A.; Dementhon, K.; Csonka, K.; Mellado-Mojica, E.; Dias Valerio, A.; Gonzalez-Hernandez, R.J.; Courdavault, V.; Clastre, M.; Hernandez, N.V.; et al. Disruption of protein mannosylation affects Candida guilliermondii cell wall, immune sensing, and virulence. Front. Microbiol. 2016, 7, 1951. [Google Scholar] [CrossRef]
- Mora-Montes, H.M.; McKenzie, C.; Bain, J.M.; Lewis, L.E.; Erwig, L.P.; Gow, N.A. Interactions between macrophages and cell wall oligosaccharides of Candida albicans. Methods Mol. Biol. 2012, 845, 247–260. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Graham, L.M.; Tsoni, S.V.; Willment, J.A.; Williams, D.L.; Taylor, P.R.; Gordon, S.; Dennehy, K.; Brown, G.D. Soluble dectin-1 as a tool to detect beta-glucans. J. Immunol. Methods 2006, 314, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Marakalala, M.J.; Vautier, S.; Potrykus, J.; Walker, L.A.; Shepardson, K.M.; Hopke, A.; Mora-Montes, H.M.; Kerrigan, A.; Netea, M.G.; Murray, G.I.; et al. Differential adaptation of Candida albicans in vivo modulates immune recognition by dectin-1. PLoS Pathog. 2013, 9, e1003315. [Google Scholar] [CrossRef]
- Mora-Montes, H.M.; Netea, M.G.; Ferwerda, G.; Lenardon, M.D.; Brown, G.D.; Mistry, A.R.; Kullberg, B.J.; O’Callaghan, C.A.; Sheth, C.C.; Odds, F.C.; et al. Recognition and blocking of innate immunity cells by Candida albicans chitin. Infect. Immun. 2011, 79, 1961–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Garcia, L.A.; Csonka, K.; Flores-Carreon, A.; Estrada-Mata, E.; Mellado-Mojica, E.; Nemeth, T.; Lopez-Ramirez, L.A.; Toth, R.; Lopez, M.G.; Vizler, C.; et al. Role of protein glycosylation in Candida parapsilosis cell wall integrity and host interaction. Front. Microbiol. 2016, 7, 306. [Google Scholar] [CrossRef]
- Hernandez-Chavez, M.J.; Franco, B.; Clavijo-Giraldo, D.M.; Hernandez, N.V.; Estrada-Mata, E.; Mora-Montes, H.M. Role of protein phosphomannosylation in the Candida tropicalis-macrophage interaction. FEMS Yeast Res. 2018, 18. [Google Scholar] [CrossRef]
- Gonzalez-Hernandez, R.J.; Jin, K.; Hernandez-Chavez, M.J.; Diaz-Jimenez, D.F.; Trujillo-Esquivel, E.; Clavijo-Giraldo, D.M.; Tamez-Castrellon, A.K.; Franco, B.; Gow, N.A.R.; Mora-Montes, H.M. Phosphomannosylation and the functional analysis of the extended Candida albicans MNN4-like gene family. Front. Microbiol. 2017, 8, 2156. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, S.N.; Medoff, G.; Kobayashi, G.S.; Kwan, C.N.; Schlessinger, D. Antifungal properties of polymyxin B and its potentiation of tetracycline as an antifungal agent. Antimicrob. Agents Chemother. 1972, 2, 36–40. [Google Scholar] [CrossRef] [Green Version]
- Mora-Montes, H.M.; Lopez-Romero, E.; Zinker, S.; Ponce-Noyola, P.; Flores-Carreon, A. Hydrolysis of Man9GlcNAc2 and Man8GlcNAc2 oligosaccharides by a purified alpha-mannosidase from Candida albicans. Glycobiology 2004, 14, 593–598. [Google Scholar] [CrossRef]
- Bates, S.; Hughes, H.B.; Munro, C.A.; Thomas, W.P.H.; MacCallum, D.M.; Bertram, G.; Atrih, A.; Ferguson, M.A.J.; Brown, A.J.P.; Odds, F.C.; et al. Outer chain N-glycans are required for cell wall integrity and virulence of Candida albicans. J. Biol. Chem. 2006, 281, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Bates, S.; MacCallum, D.M.; Bertram, G.; Munro, C.A.; Hughes, H.B.; Buurman, E.T.; Brown, A.J.P.; Odds, F.C.; Gow, N.A.R. Candida albicans Pmr1p, a secretory pathway P-type Ca2+/Mn2+-ATPase, is required for glycosylation and virulence. J. Biol. Chem. 2005, 280, 23408–23415. [Google Scholar] [CrossRef] [Green Version]
- Mora-Montes, H.M.; Bates, S.; Netea, M.G.; Castillo, L.; Brand, A.; Buurman, E.T.; Diaz-Jimenez, D.F.; Jan Kullberg, B.; Brown, A.J.; Odds, F.C.; et al. A multifunctional mannosyltransferase family in Candida albicans determines cell wall mannan structure and host-fungus interactions. J. Biol. Chem. 2010, 285, 12087–12095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gow, N.A.R.; Netea, M.G.; Munro, C.A.; Ferwerda, G.; Bates, S.; Mora-Montes, H.M.; Walker, L.; Jansen, T.; Jacobs, L.; Tsoni, V.; et al. Immune recognition of Candida albicans beta-glucan by dectin-1. J. Infect. Dis. 2007, 196, 1565–1571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, M.; Clemons, K.V.; Stevens, D.A. Heat-killed yeast as a pan-fungal vaccine. Methods Mol. Biol. 2017, 1625, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Yu, R.; Zhen, D.; Tao, S.; Schmidt, M.; Han, L. β-1,3-Glucan-induced host phospholipase D activation is involved in Aspergillus fumigatus internalization into type II human pneumocyte A549 cells. PLoS ONE 2011, 6, e21468. [Google Scholar] [CrossRef] [Green Version]
- Heinsbroek, S.E.M.; Taylor, P.R.; Martinez, F.O.; Martinez-Pomares, L.; Brown, G.D.; Gordon, S. Stage-specific sampling by pattern recognition receptors during Candida albicans phagocytosis. PLoS Pathog. 2008, 4, e1000218. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Chavez, M.J.; Clavijo-Giraldo, D.M.; Novak, A.; Lozoya-Perez, N.E.; Martinez-Alvarez, J.A.; Salinas-Marin, R.; Hernandez, N.V.; Martinez-Duncker, I.; Gacser, A.; Mora-Montes, H.M. Role of protein mannosylation in the Candida tropicalis-host Interaction. Front. Microbiol. 2019, 10, 2743. [Google Scholar] [CrossRef] [Green Version]
- Luther, K.; Torosantucci, A.; Brakhage, A.A.; Heesemann, J.; Ebel, F. Phagocytosis of Aspergillus fumigatus conidia by murine macrophages involves recognition by the dectin-1 beta-glucan receptor and Toll-like receptor 2. Cell Microbiol. 2007, 9, 368–381. [Google Scholar] [CrossRef]
- Erwig, L.P.; Gow, N.A. Interactions of fungal pathogens with phagocytes. Nat. Rev. Microbiol. 2016, 14, 163–176. [Google Scholar] [CrossRef]
- Kim, C.H.; Shin, Y.P.; Noh, M.Y.; Jo, Y.H.; Han, Y.S.; Seong, Y.S.; Lee, I.H. An insect multiligand recognition protein functions as an opsonin for the phagocytosis of microorganisms. J. Biol. Chem. 2010, 285, 25243–25250. [Google Scholar] [CrossRef] [Green Version]
- Whitten, M.M.; Tew, I.F.; Lee, B.L.; Ratcliffe, N.A. A novel role for an insect apolipoprotein (apolipophorin III) in beta-1,3-glucan pattern recognition and cellular encapsulation reactions. J. Immunol. 2004, 172, 2177–2185. [Google Scholar] [CrossRef] [Green Version]
- Tamez-Castrellon, A.K.; Romeo, O.; Garcia-Carnero, L.C.; Lozoya-Perez, N.E.; Mora-Montes, H.M. Virulence factors in Sporothrix schenckii, one of the causative agents of sporotrichosis. Curr. Protein Pept. Sci. 2020, 21, 295–312. [Google Scholar] [CrossRef] [PubMed]
- Tachibana, T.; Matsuyama, T.; Mitsuyama, M. Characteristic infectivity of Sporothrix schenckii to mice depending on routes of infection and inherent fungal pathogenicity. Med Mycol. 1998, 36, 21–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trujillo-Esquivel, E.; Martinez-Alvarez, J.A.; Clavijo-Giraldo, D.M.; Hernandez, N.V.; Flores-Martinez, A.; Ponce-Noyola, P.; Mora-Montes, H.M. The Sporothrix schenckii gene encoding for the ribosomal protein L6 has constitutive and stable expression and works as an endogenous control in gene expression analysis. Front. Microbiol. 2017, 8, 1676. [Google Scholar] [CrossRef] [PubMed]
- Marimon, R.; Cano, J.; Gené, J.; Sutton, D.A.; Kawasaki, M.; Guarro, J. Sporothrix brasiliensis, S. globosa, and S. mexicana, three new Sporothrix species of clinical interest. J. Clin. Microbiol. 2007, 45, 3198–3206. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.J.P.; Brown, G.D.; Netea, M.G.; Gow, N.A.R. Metabolism impacts upon Candida immunogenicity and pathogenicity at multiple levels. Trends Microbiol. 2014, 22, 614–622. [Google Scholar] [CrossRef] [Green Version]
- Ciesielczuk, H.; Betts, J.; Phee, L.; Doumith, M.; Hope, R.; Woodford, N.; Wareham, D.W. Comparative virulence of urinary and bloodstream isolates of extra-intestinal pathogenic Escherichia coli in a Galleria mellonella model. Virulence 2015, 6, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Wand, M.E.; McCowen, J.W.I.; Nugent, P.G.; Sutton, J.M. Complex interactions of Klebsiella pneumoniae with the host immune system in a Galleria mellonella infection model. J. Med Microbiol. 2013, 62, 1790–1798. [Google Scholar] [CrossRef]
- Matha, V.; Mracek, Z. Changes in haemocyte counts in Galleria mellonella (L.) (Lepidoptera: Galleriidae) larvae infected with Steinernema sp. (Nematoda: Steinernematidae). Nematologica 1983, 30, 86–89. [Google Scholar] [CrossRef]
- McKenzie, C.G.; Koser, U.; Lewis, L.E.; Bain, J.M.; Mora-Montes, H.M.; Barker, R.N.; Gow, N.A.; Erwig, L.P. Contribution of Candida albicans cell wall components to recognition by and escape from murine macrophages. Infect. Immun. 2010, 78, 1650–1658. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Uscanga, B.; François, J.M. A study of the yeast cell wall composition and structure in response to growth conditions and mode of cultivation. Lett. Appl. Microbiol. 2003, 37, 268–274. [Google Scholar] [CrossRef]
- Giosa, D.; Felice, M.R.; Giuffre, L.; Aiese Cigliano, R.; Paytuvi-Gallart, A.; Lo Passo, C.; Barresi, C.; D’Alessandro, E.; Huang, H.; Criseo, G.; et al. Transcriptome-wide expression profiling of Sporothrix schenckii yeast and mycelial forms and the establishment of the Sporothrix Genome DataBase. Microb. Genom. 2020. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Colony-Forming Units (×105) a | Cytotoxicity (%) b | Hemocytes (×106)/mL c | Phenoloxidase d |
|---|---|---|---|---|
| PBS e | 0.0 ± 0 | 13.1 ± 3.7 | 2.9 ± 0.6 | 0.7 ± 0.2 |
| Sporothrix schenckii | ||||
| YPD f | 2.2 ± 0.4 | 77.4 ± 9.8 * | 7.8 ± 0.8 * | 2.8 ± 0.3 * |
| BHI f | 2.4 ± 0.2 | 52.7 ± 12.4 * | 5.8 ± 0.5 * | 2.0 ± 0.3 * |
| YP f | 1.9 ± 0.2 | 32.4 ± 11.4 † | 3.7 ± 0.5 † | 1.3 ± 0.2 † |
| YNB f | 2.2 ± 0.2 | 36.7 ± 9.7 † | 3.9 ± 0.3 † | 1.3 ± 0.4 † |
| Sporothrix brasiliensis | ||||
| YPD | 2.3 ± 0.3 | 89.4 ± 7.6 * | 9.2 ± 0.5 * | 3.5 ± 0.3 * |
| BHI | 1.8 ± 0.3 | 60.1 ± 12.7 * | 5.1 ± 0.9 * | 2.6 ± 0.4 * |
| YP | 2.4 ± 0.4 | 42.4 ± 12.4 † | 3.9 ± 0.3 † | 1.6 ± 0.5 † |
| YNB | 2.2 ± 0.4 | 37.0 ± 9.6 † | 4.4 ± 0.6 † | 1.5 ± 0.6 † |
| Sporothrix globosa | ||||
| YPD | 2.2 ± 0.3 | 33.4 ± 10.4 | 3.9 ± 0.7 | 1.5 ± 0.3 |
| BHI | 2.0 ± 0.4 | 34.8 ± 12.4 | 4.1 ± 0.8 | 1.2 ± 0.4 |
| YP | 2.4 ± 0.5 | 37.5 ± 8.8 | 4.2 ± 0.4 | 1.5 ± 0.6 |
| YNB | 1.9 ± 0.5 | 40.4 ± 11.1 | 3.6 ± 0.6 | 1.2 ± 0.2 |
| Organism | Uptake by Hemocytes (×103 Cells) | Uptake by Hemocytes (×103 cells) + Laminarin a | Protease Activity b | Lipase/Esterase Activy c |
|---|---|---|---|---|
| PBS e | 0.0 ± 0 | 0.0 ± 0 | 0.01 ± 0.005 | 0.0 ± 0.0 |
| Sporothrix schenckii | ||||
| YPD d | 6.5 ± 0.6 | 3.8 ± 0.8 † | 357.4 ± 89.7 | 38.2 ± 13.5 |
| BHI d | 7.4 ± 0.8 | 3.2 ± 0.6 † | 387.1 ± 76.1 | 42.7± 8.9 |
| YP d | 12.9 ± 0.7 * | 2.9 ± 1.0 † | 366.7 ± 97.4 | 33.7± 11.4 |
| YNB d | 13.4 ± 0.4 * | 3.3 ± 0.8 † | 438.7 ± 77.5 | 44.5 ± 10.0 |
| Sporothrix brasiliensis | ||||
| YPD | 3.8 ± 0.2 | 2.4 ± 0.6 † | 402.8 ± 92.1 | 35.2 ± 6.8 |
| BHI | 4.4 ± 0.6 | 2.5 ± 0.8 † | 387.2 ± 47.8 | 33.6 ± 9.1 |
| YP | 10.4 ± 0.7 * | 3.1 ± 1.1 † | 364.1 ± 67.0 | 38.4 ± 8.8 |
| YNB | 11.2 ± 0.9 * | 2.5 ± 1.3 † | 442.1 ± 57.9 | 40.1 ± 10.4 |
| Sporothrix globosa | ||||
| YPD | 11.4 ± 0.8 | 3.9 ± 0.8 † | 357 ± 99.2 | 40.5 ± 11.4 |
| BHI | 12.2 ± 0.7 | 2.4 ± 1.2 † | 380 ± 95.4 | 35.2 ± 9.7 |
| YP | 13.4 ± 0.8 | 4.1 ± 0.9 † | 411 ± 62.7 | 33.7 ± 10.4 |
| YNB | 12.1 ± 0.6 | 3.7 ± 1.0 † | 435 ± 41.8 | 44.1 ± 8.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lozoya-Pérez, N.E.; Clavijo-Giraldo, D.M.; Martínez-Duncker, I.; García-Carnero, L.C.; López-Ramírez, L.A.; Niño-Vega, G.A.; Mora-Montes, H.M. Influences of the Culturing Media in the Virulence and Cell Wall of Sporothrix schenckii, Sporothrix brasiliensis, and Sporothrix globosa. J. Fungi 2020, 6, 323. https://doi.org/10.3390/jof6040323
Lozoya-Pérez NE, Clavijo-Giraldo DM, Martínez-Duncker I, García-Carnero LC, López-Ramírez LA, Niño-Vega GA, Mora-Montes HM. Influences of the Culturing Media in the Virulence and Cell Wall of Sporothrix schenckii, Sporothrix brasiliensis, and Sporothrix globosa. Journal of Fungi. 2020; 6(4):323. https://doi.org/10.3390/jof6040323
Chicago/Turabian StyleLozoya-Pérez, Nancy E., Diana M. Clavijo-Giraldo, Iván Martínez-Duncker, Laura C. García-Carnero, Luz A. López-Ramírez, Gustavo A. Niño-Vega, and Héctor M. Mora-Montes. 2020. "Influences of the Culturing Media in the Virulence and Cell Wall of Sporothrix schenckii, Sporothrix brasiliensis, and Sporothrix globosa" Journal of Fungi 6, no. 4: 323. https://doi.org/10.3390/jof6040323
APA StyleLozoya-Pérez, N. E., Clavijo-Giraldo, D. M., Martínez-Duncker, I., García-Carnero, L. C., López-Ramírez, L. A., Niño-Vega, G. A., & Mora-Montes, H. M. (2020). Influences of the Culturing Media in the Virulence and Cell Wall of Sporothrix schenckii, Sporothrix brasiliensis, and Sporothrix globosa. Journal of Fungi, 6(4), 323. https://doi.org/10.3390/jof6040323