Hog1 Controls Lipids Homeostasis Upon Osmotic Stress in Candida albicans



Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Viability Assays
2.3. Staining with Propidium Iodide
2.4. Study of Lipid Content
2.5. Study of Endocytosis
2.6. Fluorescence Microscopy
3. Results
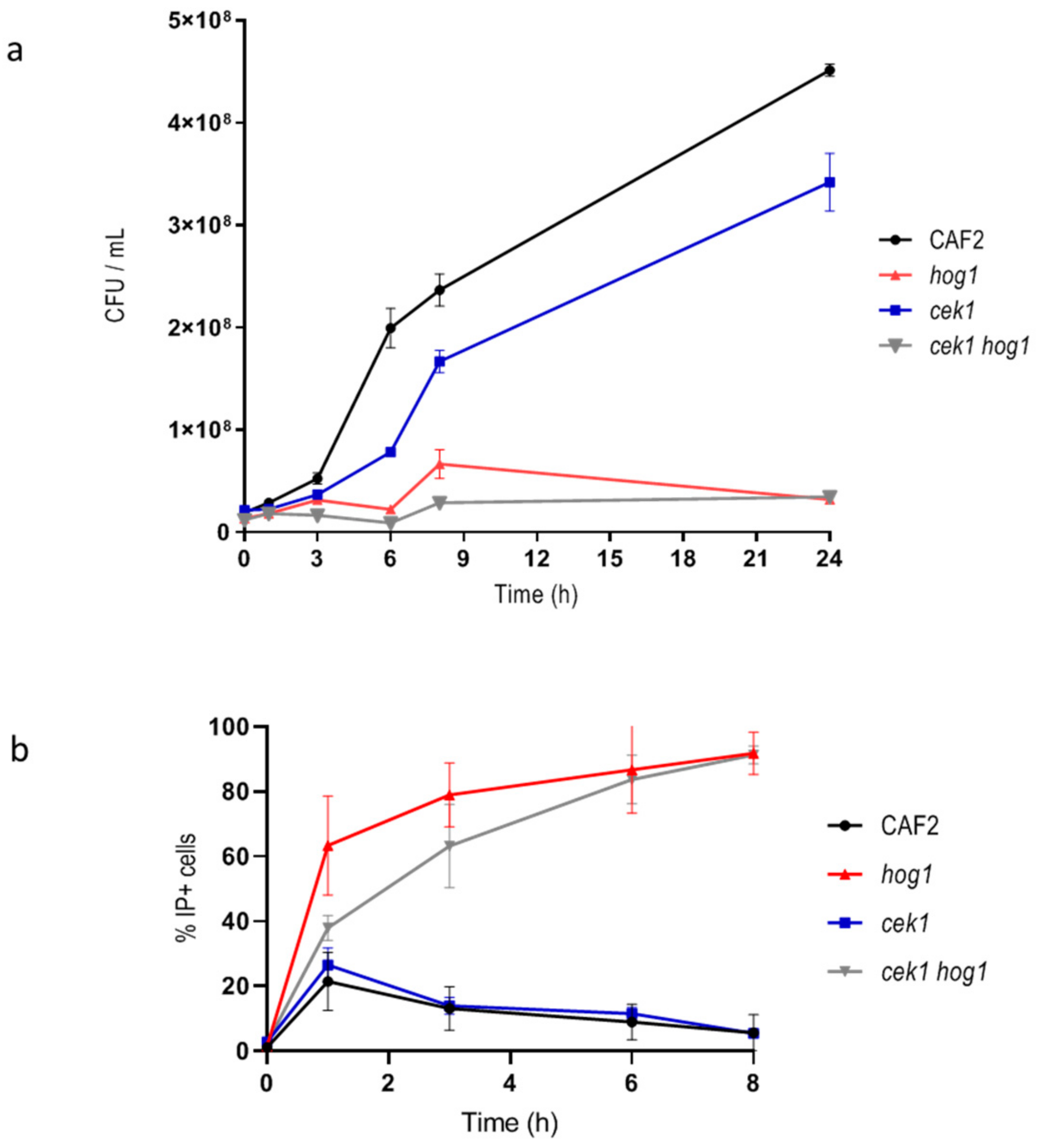
3.1. Hog1 Controls Cell Permeability upon Osmotic Stress
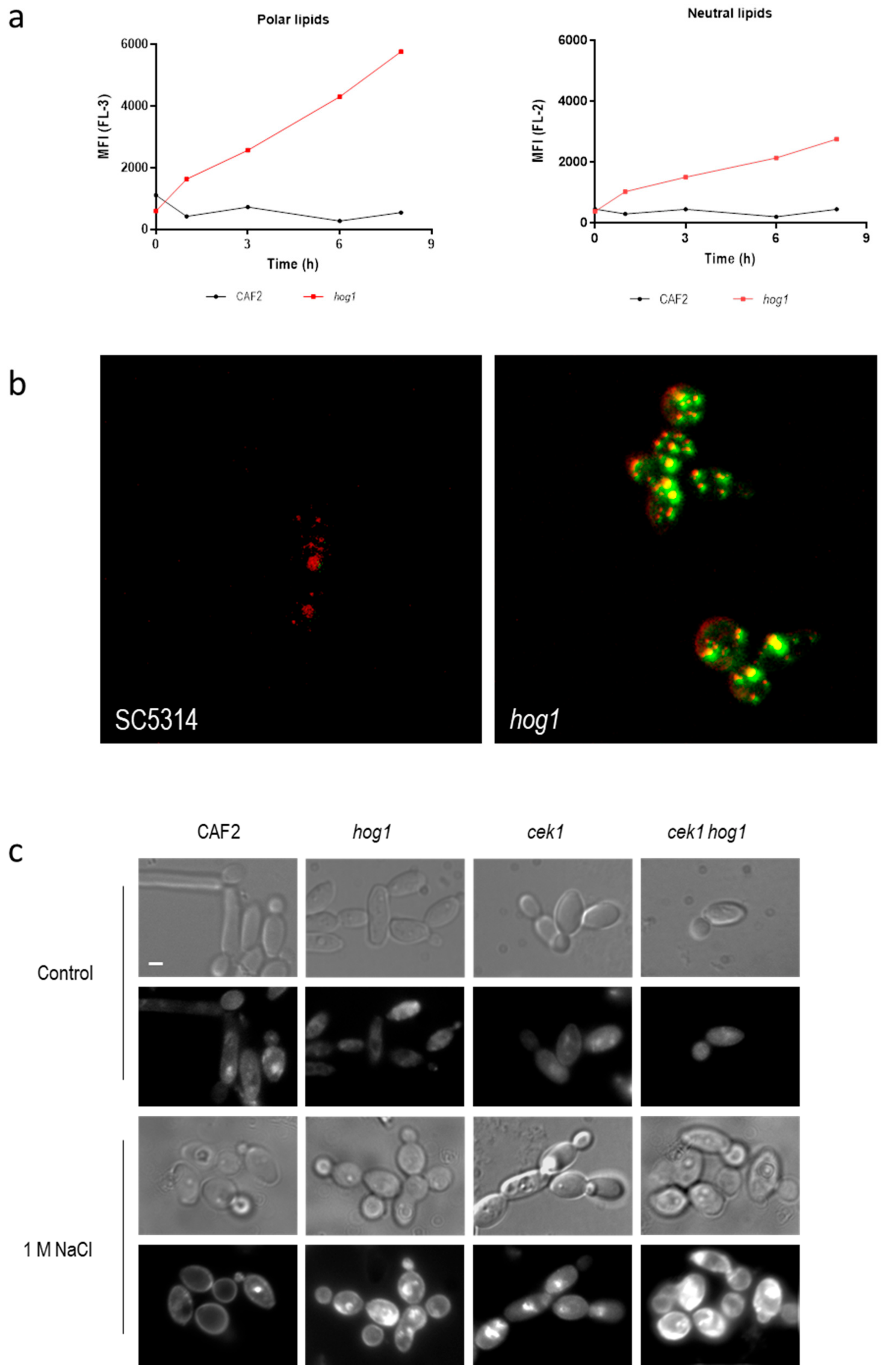
3.2. Hog1 Mediates Lipids Homeostasis Upon Osmotic Stress
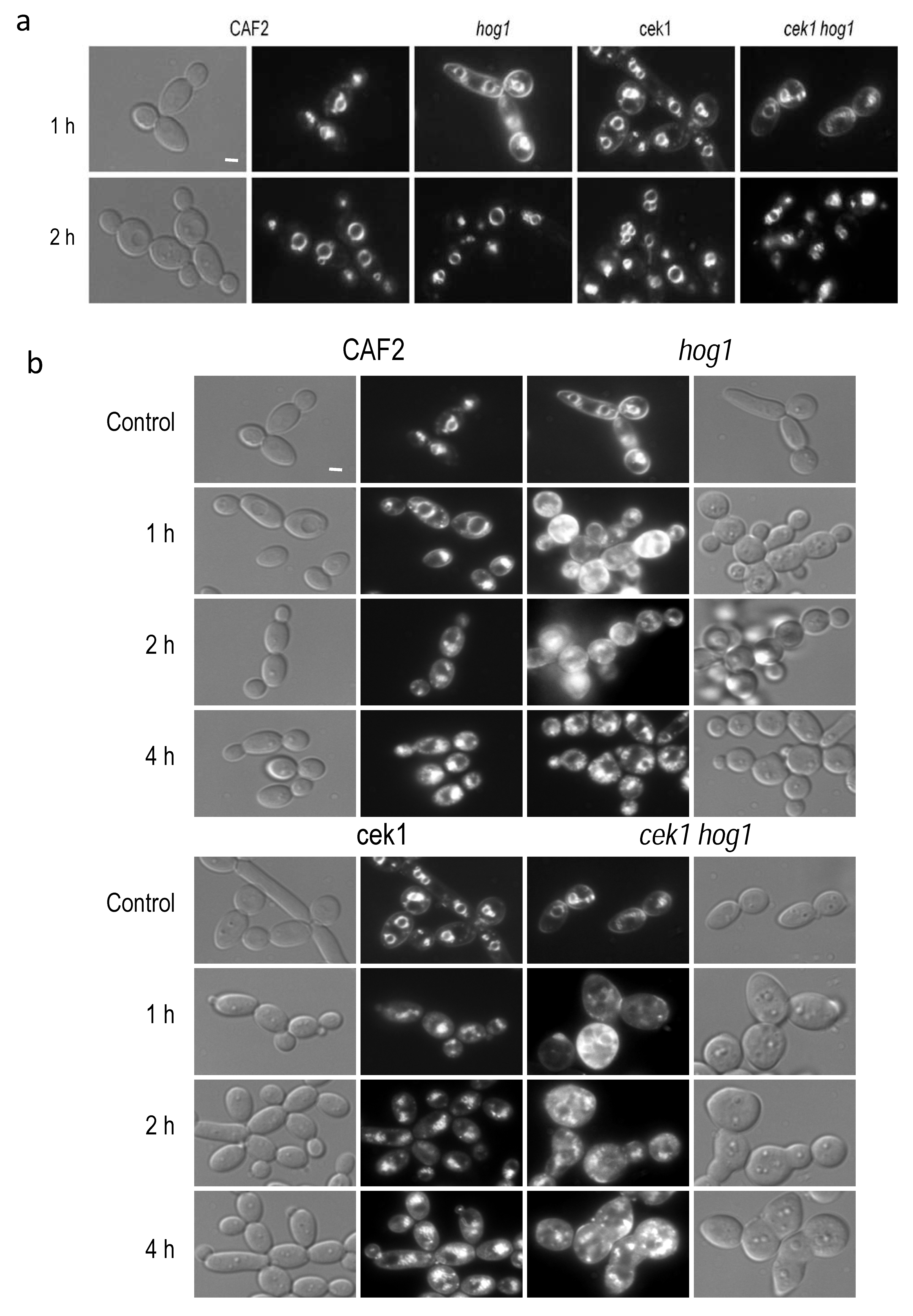
3.3. Osmotic Stress Delays the Endocytic Trafficking in hog1 Defective Mutants
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Odds, F.C. Pathogenesis of Candida infections. J. Am. Acad. Pharmacol. 1994, 31, S2–S5. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Diekema, D.J. Epidemiology of invasive mycoses in North America. Crit. Rev. Microbiol. 2010, 36, 1–53. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Diekema, D.J. Epidemiology of invasive candidiasis: A persistent public health problem. Clin. Microbiol. Rev. 2007, 20, 133–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcos-Zambrano, L.J.; Escribano, P.; Bouza, E.; Guinea, J. Erratum to “Use of molecular typing tools for the study of hospital outbreaks of candidemia”. Rev. Iberoam. Micol. 2016, 33, 261. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, S.; Chhillar, A.K.; Sharma, N.; Choudhary, P.; Punia, A.; Balhara, M.; Kaushik, K.; Parmar, V.S. Candida auris and Nosocomial infection. Curr. Drug Targets 2020, 21, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Day, A.M.; Quinn, J. Stress-Activated Protein Kinases in Human Fungal Pathogens. Front. Cell Infect. Microbiol. 2019, 9, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kultz, D. Phylogenetic and functional classification of mitogen- and stress-activated protein kinases. J. Mol. Evol. 1998, 46, 571–588. [Google Scholar] [CrossRef]
- Chen, R.E.; Thorner, J. Function and regulation in MAPK signaling pathways: Lessons learned from the yeast Saccharomyces cerevisiae. Biochim. Biophys. Acta 2007, 1773, 1311–1340. [Google Scholar] [CrossRef] [Green Version]
- Navarro-García, F.; Sanchez, M.; Pla, J.; Nombela, C. Functional characterization of the MKC1 gene of Candida albicans, which encodes a mitogen-activated protein kinase homolog related to cell integrity. Mol. Cell. Biol. 1995, 15, 2197–2206. [Google Scholar] [CrossRef] [Green Version]
- Kumamoto, C.A. A contact-activated kinase signals Candida albicans invasive growth and biofilm development. Proc. Natl. Acad. Sci. USA 2005, 102, 5576–5581. [Google Scholar] [CrossRef] [Green Version]
- Navarro-García, F.; Eisman, B.; Fiuza, S.M.; Nombela, C.; Pla, J. The MAP kinase Mkc1p is activated under different stress conditions in Candida albicans. Microbiology 2005, 151, 2737–2749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díez-Orejas, R.; Molero, G.; Navarro-García, F.; Pla, J.; Nombela, C.; Sánchez-Pérez, M. Reduced virulence of Candida albicans MKC1 mutants: A role for a mitogen-activated protein kinase in pathogenesis. Infect. Immun. 1997, 65, 833–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiteway, M.; Dignard, D.; Thomas, D.Y. Dominant negative selection of heterologous genes: Isolation of Candida albicans genes that interfere with Saccharomyces cerevisiae mating factor-induced cell cycle arrest. Proc. Natl. Acad. Sci. USA 1992, 89, 9410–9414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csank, C.; Schröppel, K.; Leberer, E.; Harcus, D.; Mohamed, O.; Meloche, S.; Thomas, D.Y.; Whiteway, M. Roles of the Candida albicans mitogen-activated protein kinase homolog, Cek1p, in hyphal development and systemic candidiasis. Infect. Immun. 1998, 66, 2713–2721. [Google Scholar] [CrossRef] [Green Version]
- Román, E.; Nombela, C.; Pla, J. The Sho1 adaptor protein links oxidative stress to morphogenesis and cell wall biosynthesis in the fungal pathogen Candida albicans. Mol. Cell. Biol. 2005, 25, 10611–10627. [Google Scholar] [CrossRef] [Green Version]
- Román, E.; Cottier, F.; Ernst, J.F.; Pla, J. Msb2 signaling mucin controls activation of Cek1 mitogen-activated protein kinase in Candida albicans. Eukaryot. Cell 2009, 8, 1235–1249. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Chen, J.; Lane, S.; Liu, H. A conserved mitogen-activated protein kinase pathway is required for mating in Candida albicans. Mol. Microbiol. 2002, 46, 1335–1344. [Google Scholar] [CrossRef] [Green Version]
- Galan-Diez, M.; Arana, D.M.; Serrano-Gomez, D.; Kremer, L.; Casasnovas, J.M.; Ortega, M.; Cuesta-Dominguez, A.; Corbi, A.L.; Pla, J.; Fernandez-Ruiz, E. Candida albicans beta-glucan exposure is controlled by the fungal CEK1-mediated mitogen-activated protein kinase pathway that modulates immune responses triggered through dectin-1. Infect. Immun. 2010, 78, 1426–1436. [Google Scholar] [CrossRef] [Green Version]
- Roman, E.; Correia, I.; Salazin, A.; Fradin, C.; Jouault, T.; Poulain, D.; Liu, F.T.; Pla, J. The Cek1mediated MAP kinase pathway regulates exposure of alpha1,2 and beta1,2mannosides in the cell wall of Candida albicans modulating immune recognition. Virulence 2016, 7, 558–577. [Google Scholar] [CrossRef] [Green Version]
- Correia, I.; Roman, E.; Prieto, D.; Eisman, B.; Pla, J. Complementary roles of the Cek1 and Cek2 MAP kinases in Candida albicans cell-wall biogenesis. Future Microbiol. 2016, 11, 51–67. [Google Scholar] [CrossRef]
- Herrero de Dios, C.; Alonso-Monge, R.; Pla, J. The lack of upstream elements of the Cek1 and Hog1 mediated pathways leads to a synthetic lethal phenotype upon osmotic stress in Candida albicans. Fungal Genet. Biol. 2014, 69, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Monge, R.; Navarro-García, F.; Román, E.; Negredo, A.I.; Eisman, B.; Nombela, C.; Pla, J. The Hog1 mitogen-activated protein kinase is essential in the oxidative stress response and chlamydospore formation in Candida albicans. Eukaryot. Cell 2003, 2, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.A.; Nicholls, S.; Morgan, B.A.; Brown, A.J.; Quinn, J. A conserved stress-activated protein kinase regulates a core stress response in the human pathogen Candida albicans. Mol. Biol. Cell 2004, 15, 4179–4190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero-de-Dios, C.; Day, A.M.; Tillmann, A.T.; Kastora, S.L.; Stead, D.; Salgado, P.S.; Quinn, J.; Brown, A.J.P. Redox regulation, rather than stress-induced phosphorylation, of a Hog1 mitogen-activated protein kinase modulates its nitrosative-stress-specific outputs. MBio 2018, 9, e02229-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urrialde, V.; Prieto, D.; Pla, J.; Alonso-Monge, R. The Pho4 transcription factor mediates the response to arsenate and arsenite in Candida albicans. Front. Microbiol. 2015, 6, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Monge, R.; Navarro-García, F.; Molero, G.; Díez-Orejas, R.; Gustin, M.; Pla, J.; Sánchez, M.; Nombela, C. Role of the mitogen-activated protein kinase Hog1p in morphogenesis and virulence of Candida albicans. J. Bacteriol. 1999, 181, 3058–3068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arana, D.M.; Alonso-Monge, R.; Du, C.; Calderone, R.; Pla, J. Differential susceptibility of mitogen-activated protein kinase pathway mutants to oxidative-mediated killing by phagocytes in the fungal pathogen Candida albicans. Cell. Microbiol. 2007, 9, 1647–1659. [Google Scholar] [CrossRef]
- Prieto, A.D.; Román, E.; Correia, I.; Pla, J. The HOG pathway is critical for the colonization of the mouse gastrointestinal tract by Candida albicans. PLoS ONE 2014, 9, e87128. [Google Scholar] [CrossRef] [Green Version]
- Enjalbert, B.; Smith, D.A.; Cornell, M.J.; Alam, I.; Nicholls, S.; Brown, A.J.; Quinn, J. Role of the Hog1 stress-activated protein kinase in the global transcriptional response to stress in the fungal pathogen Candida albicans. Mol. Biol. Cell 2006, 17, 1018–1032. [Google Scholar] [CrossRef] [Green Version]
- Fonzi, W.A.; Irwin, M.Y. Isogenic strain construction and gene mapping in Candida albicans. Genetics 1993, 134, 717–728. [Google Scholar]
- American Public Health Association. 9215 Heterotrophic Plate Count. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 2017. [Google Scholar] [CrossRef]
- Kwolek-Mirek, M.; Zadrag-Tecza, R. Comparison of methods used for assessing the viability and vitality of yeast cells. FEMS Yeast Res. 2014, 14, 1068–1079. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, J.M.; Alvarez, A.; Nombela, C.; Sánchez, M. Flow cytometric analysis of Saccharomyces cerevisiae autolytic mutants and protoplasts. Yeast 1992, 8, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, M.R.; Smant, W.; de Boer, W.; Dijksterhuis, J. Filipin is a reliable in situ marker of ergosterol in the plasma membrane of germinating conidia (spores) of Penicillium discolor and stains intensively at the site of germ tube formation. J. Microbiol. Methods 2008, 74, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Maayan, I.; Engelberg, D. The yeast MAPK Hog1 is not essential for immediate survival under osmostress. FEBS Lett. 2009, 583, 2015–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- San José, C.; Alonso-Monge, R.; Pérez-Díaz, R.M.; Pla, J.; Nombela, C. The mitogen-activated protein kinase homolog HOG1 gene controls glycerol accumulation in the pathogenic fungus Candida albicans. J. Bacteriol. 1996, 178, 5850–5852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pina-Vaz, C.; Sansonetty, F.; Rodrigues, A.G.; Costa-Oliveira, S.; Tavares, C.; Martinez-de-Oliveira, J. Cytometric approach for a rapid evaluation of susceptibility of Candida strains to antifungals. Clin. Microbiol. Infect. 2001, 7, 609–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, E.; Vale-Silva, L.; Cavaleiro, C.; Salgueiro, L. Antifungal activity of the clove essential oil from Syzygium aromaticum on Candida, Aspergillus and dermatophyte species. J. Med. Microbiol. 2009, 58, 1454–1462. [Google Scholar] [CrossRef]
- Safranyos, R.G.; Caveney, S. Rates of diffusion of fluorescent molecules via cell-to-cell membrane channels in a developing tissue. J. Cell Biol. 1985, 100, 736–747. [Google Scholar] [CrossRef] [Green Version]
- Greenspan, P.; Fowler, S.D. Spectrofluorometric studies of the lipid probe, nile red. J. Lipid Res. 1985, 26, 781–789. [Google Scholar]
- Leszczynska, A.; Burzynska, B.; Plochocka, D.; Kaminska, J.; Zimnicka, M.; Kania, M.; Kiliszek, M.; Wysocka-Kapcinska, M.; Danikiewicz, W.; Szkopinska, A. Investigating the effects of statins on cellular lipid metabolism using a yeast expression system. PLoS ONE 2009, 4, e8499. [Google Scholar] [CrossRef] [Green Version]
- Grossmann, G.; Opekarova, M.; Malinsky, J.; Weig-Meckl, I.; Tanner, W. Membrane potential governs lateral segregation of plasma membrane proteins and lipids in yeast. EMBO J. 2007, 26, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Walther, T.C.; Brickner, J.H.; Aguilar, P.S.; Bernales, S.; Pantoja, C.; Walter, P. Eisosomes mark static sites of endocytosis. Nature 2006, 439, 998–1003. [Google Scholar] [CrossRef] [PubMed]
- Vida, T.A.; Emr, S.D. A new vital stain for visualizing vacuolar membrane dynamics and endocytosis in yeast. J. Cell Biol. 1995, 128, 779–792. [Google Scholar] [CrossRef] [PubMed]
- Vogel, B.F.; Hansen, L.T.; Mordhorst, H.; Gram, L. The survival of Listeria monocytogenes during long term desiccation is facilitated by sodium chloride and organic material. Int. J. Food Microbiol. 2010, 140, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Saito, H.; Posas, F. Response to hyperosmotic stress. Genetics 2012, 192, 289–318. [Google Scholar] [CrossRef] [Green Version]
- De Nadal, E.; Posas, F. Osmostress-induced gene expression—A model to understand how stress-activated protein kinases (SAPKs) regulate transcription. FEBS J. 2015, 282, 3275–3285. [Google Scholar] [CrossRef]
- Roman, E.; Correia, I.; Prieto, D.; Alonso, R.; Pla, J. The HOG MAPK pathway in Candida albicans: More than an osmosensing pathway. Int. Microbiol. 2020, 23, 23–29. [Google Scholar] [CrossRef]
- Ma, D.; Li, R. Current understanding of HOG-MAPK pathway in Aspergillus fumigatus. Mycopathologia 2013, 175, 13–23. [Google Scholar] [CrossRef]
- Ha, G.S.; El-Dalatony, M.M.; Kim, D.H.; Salama, E.S.; Kurade, M.B.; Roh, H.S.; El-Fatah Abomohra, A.; Jeon, B.H. Biocomponent-based microalgal transformations into biofuels during the pretreatment and fermentation process. Bioresour. Technol. 2020, 302, 122809. [Google Scholar] [CrossRef]
- Venkata Subhash, G.; Rohit, M.V.; Devi, M.P.; Swamy, Y.V.; Venkata Mohan, S. Temperature induced stress influence on biodiesel productivity during mixotrophic microalgae cultivation with wastewater. Bioresour. Technol. 2014, 169, 789–793. [Google Scholar] [CrossRef]
- Manzanares-Estreder, S.; Espi-Bardisa, J.; Alarcon, B.; Pascual-Ahuir, A.; Proft, M. Multilayered control of peroxisomal activity upon salt stress in Saccharomyces cerevisiae. Mol. Microbiol. 2017, 104, 851–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Monge, R.; Carvaihlo, S.; Nombela, C.; Rial, E.; Pla, J. The Hog1 MAP kinase controls respiratory metabolism in the fungal pathogen Candida albicans. Microbiology 2009, 155, 413–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slessareva, J.E.; Routt, S.M.; Temple, B.; Bankaitis, V.A.; Dohlman, H.G. Activation of the phosphatidylinositol 3-kinase Vps34 by a G protein alpha subunit at the endosome. Cell 2006, 126, 191–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappell, S.D.; Dohlman, H.G. Selective regulation of MAP kinase signaling by an endomembrane phosphatidylinositol 4-kinase. J. Biol. Chem. 2011, 286, 14852–14860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schink, K.O.; Tan, K.W.; Stenmark, H. Phosphoinositides in control of membrane dynamics. Annu. Rev. Cell Dev. Biol. 2016, 32, 143–171. [Google Scholar] [CrossRef] [PubMed]
- Huotari, J.; Helenius, A. Endosome maturation. EMBO J. 2011, 30, 3481–3500. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Name in the Text | Reference |
|---|---|---|---|
| CAF2 | URA3/ura3Δ::imm434 | CAF2 (wt) | [30] |
| HI3-21 | CAI4 hog1::hisG/hog1::hisG-URA3-hisG | hog1 | [28] |
| CK43B-16 | CAI4 cek1::hisG/cek1::hisG-URA3-hisG | cek1 | [14] |
| E5 | CAI4 cek1::hisG-URA3-hisG/cek1::hisG hog1::hisG/hog1::hisG ARD1/ard1::FRT pSAP2-FLP-URA3 | cek1 hog1 | [27] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrero-de-Dios, C.; Román, E.; Pla, J.; Alonso-Monge, R. Hog1 Controls Lipids Homeostasis Upon Osmotic Stress in Candida albicans. J. Fungi 2020, 6, 355. https://doi.org/10.3390/jof6040355
Herrero-de-Dios C, Román E, Pla J, Alonso-Monge R. Hog1 Controls Lipids Homeostasis Upon Osmotic Stress in Candida albicans. Journal of Fungi. 2020; 6(4):355. https://doi.org/10.3390/jof6040355
Chicago/Turabian StyleHerrero-de-Dios, Carmen, Elvira Román, Jesús Pla, and Rebeca Alonso-Monge. 2020. "Hog1 Controls Lipids Homeostasis Upon Osmotic Stress in Candida albicans" Journal of Fungi 6, no. 4: 355. https://doi.org/10.3390/jof6040355
APA StyleHerrero-de-Dios, C., Román, E., Pla, J., & Alonso-Monge, R. (2020). Hog1 Controls Lipids Homeostasis Upon Osmotic Stress in Candida albicans. Journal of Fungi, 6(4), 355. https://doi.org/10.3390/jof6040355