Morphological Characterization of the In Vitro Mycorrhizae Formed between Four Terfezia Species (Pezizaceae) with Cistus salviifolius and Cistus ladanifer—Towards Desert Truffles Production in Acid Soils




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Material
2.2. Plant Material
2.3. Mycorrhizal Synthesis
2.4. Mycorrhizal Morphotyping and Colonization Assessment
2.5. Statistical Analysis
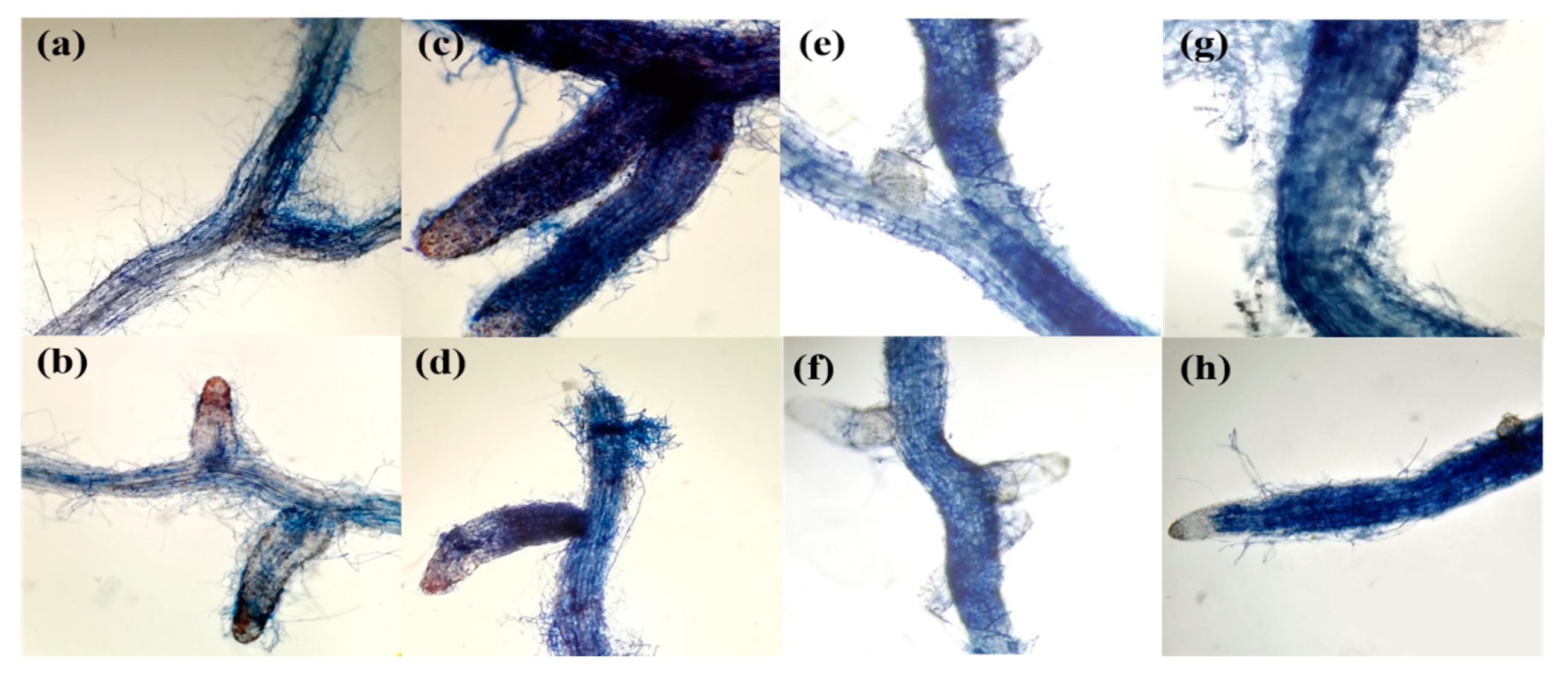
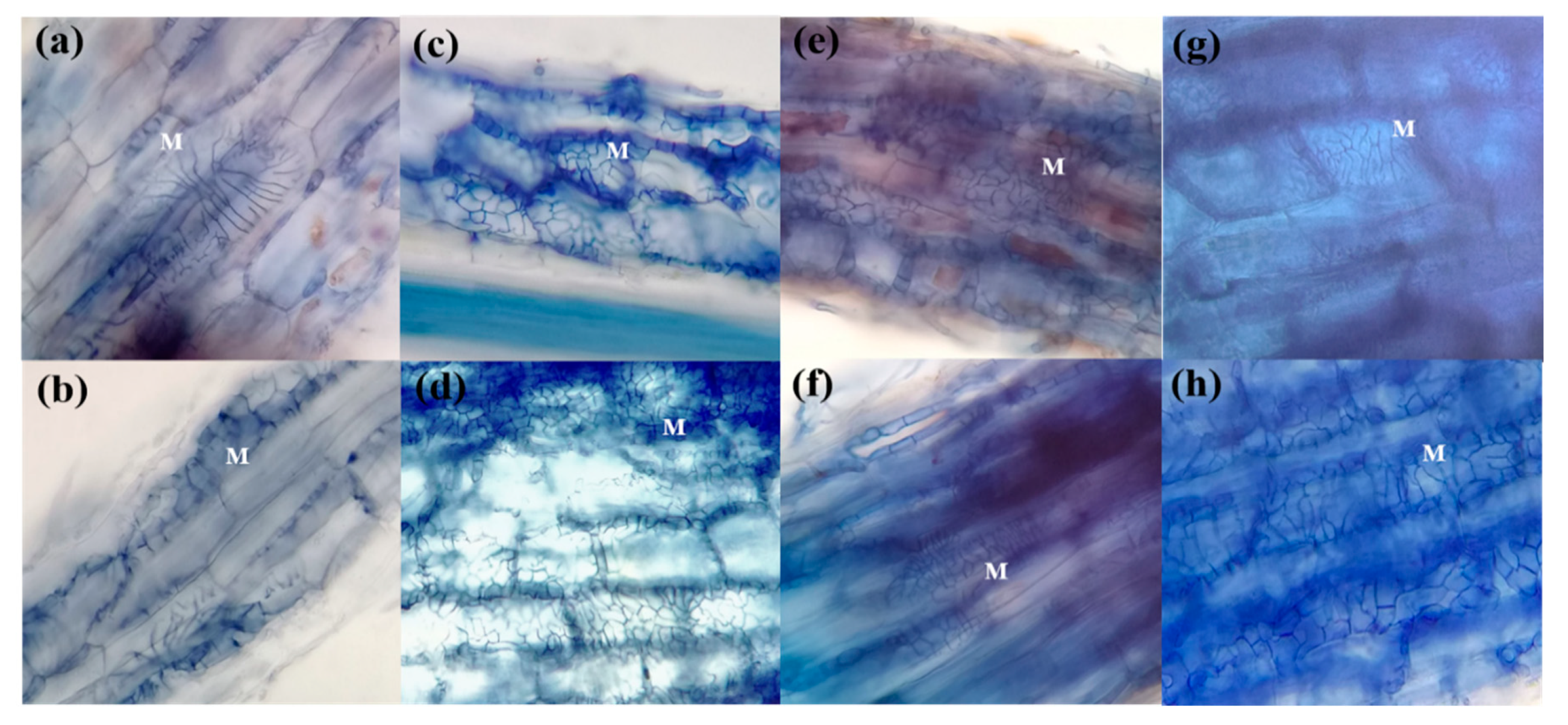
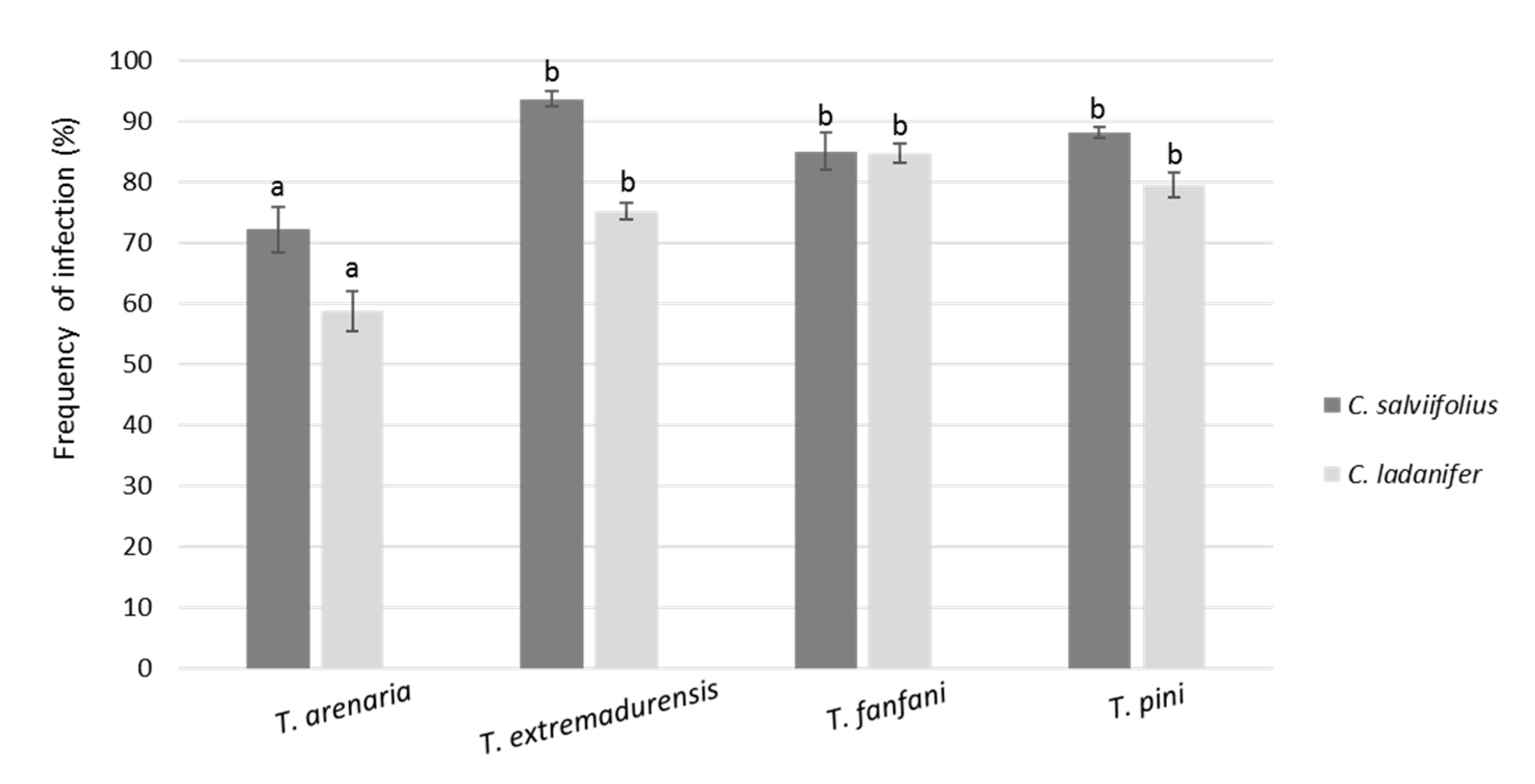
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moreno, G.; Alvarado, P.; Manjón, J.P. Hypogeous desert fungi. In Desert Truffles, Phylogeny, Physiology, Distribution and Domestication; Kagan-Zur, V., Roth-Bejerano, N., Sitrit, Y., Morte, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Morte, A.; Honrubia, M.; Gutiérrez, A. Biotechnology and cultivation of desert truffles. In Mycorrhiza: State of the Art Genetics and Molecular Biology, Eco-Function, Biotechnology, Eco-Physiology, Structure and Systematics; Varma, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Navarro-Ródenas, A.; Lozano-Carrillo, M.C.; Pérez-Gilabert, M.; Morte, A. Effect of water stress on in vitro mycelium cultures of two mycorrhizal desert truffles. Mycorrhiza 2011, 21, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Gilabert, M.; García-Carmona, F.; Morte, A. Enzymes in Terfezia claveryi ascocarps. In Desert Truffles, Phylogeny, Physiology, Distribution and Domestication; Kagan-Zur, V., Roth-Bejerano, N., Sitrit, Y., Morte, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Alsheikh, A.M. Taxonomy and Mycorrhizal Ecology of the Desert Truffles in the Genus. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 1994. [Google Scholar]
- Diez, J.; Manjon, J.L.; Martin, F. Molecular phylogeny of the mycorrhizal desert truffles (Terfezia and Tirmania), host specificity and edaphic tolerance. Mycologia 2002, 94, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Fortas, Z.; Chevalier, G. Effet des conditions de culture sur la mycorhization de l’Helianthemum guttatum par trois espèces de terfez des genres Terfezia et Tirmania d’Algerie. Can. J. Bot. 1992, 70, 2453–2460. [Google Scholar] [CrossRef]
- Kagan-Zur, V.; Roth-Bejerano, N. Desert Truffles. Fungi 2008, 1, 32–37. [Google Scholar]
- Morte, A.; Andrino, A. Domestication: Preparation of mycorrhizal seedlings. In Desert Truffles, Phylogeny, Physiology, Distribution and Domestication; Kagan-Zur, V., Roth-Bejerano, N., Sitrit, Y., Morte, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Bordallo, J.J.; Rodríguez, A.; Kaounas, V.; Camello, F.; Honrubia, M.; Morte, A. Two new Terfezia species from southern Europe. Phytotaxa 2015, 230, 239–249. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Wingfield, M.J.; Lombard, L.; Roets, F.; Swart, W.J.; Alvarado, P.; Carnegie, A.J.; Moreno, G.; Luangsaard, J.; Thangavel, R.; et al. Fungal Planet Description Sheets: 951–1041. Persoonia 2019, 43, 223–425. [Google Scholar] [CrossRef]
- Kovacs, G.M.; Balazs, T.K.; Calonge, F.D.; Martín, M.P. The diversity of Terfezia desert truffles: New species and a highly variable species complex with intra-sporocarpic nrDNA ITS heterogeneity. Mycologia 2011, 103, 841–853. [Google Scholar] [CrossRef] [Green Version]
- Águeda, B.; Parladé, J.; de Miguel, A.M.; Martínez-Peña, F. Characterization and identification of field ectomycorrhizae of Boletus edulis and Cistus ladanifer. Mycologia 2006, 98, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Nuytinck, J.; Verbeken, A.; Leonardi, M.; Pacioni, G.; Rinaldi, A.C.; Comandini, O. Characterization of Lactarius tesquorum ectomycorrhizae on Cistus sp., and molecular phylogeny of related European Lactarius taxa. Mycologia 2004, 96, 272–282. [Google Scholar] [CrossRef]
- Roth-Bejerano, N.; Navarro-Ródenas, A.; Gutiérrez, A. Types of mycorrhizal association. In Desert Truffles, Phylogeny, Physiology, Distribution and Domestication; Kagan-Zur, V., Roth-Bejerano, N., Sitrit, Y., Morte, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Zitouni-Haouar, F.H.; Fortas, Z.; Chevalier, G. Morphological characterization of mycorrhizae formed between three Terfezia species (desert truffles) and several Cistaceae and Aleppo pine. Mycorrhiza 2014, 24, 397–403. [Google Scholar] [CrossRef]
- Awameh, M.S. The response of Helianthemum salicifolium and H. ledifolium to infection by the desert truffle Terfezia boudieri. Mushroom. Sci. 1981, 11, 843–853. [Google Scholar]
- Gutiérrez, A.; Morte, A.; Honrubia, M. Morphological characterization of the mycorrhiza formed by Helianthemum almeriense Pau with Terfezia claveryi Chatin and Picoa lefebvrei (Pat.) Maire. Mycorrhiza 2003, 13, 299–307. [Google Scholar]
- Kagan-Zur, V.; Kuang, J.; Tabak, S.; Taylor, F.W.; Roth-Bejerano, N. Potential verification of a host plant for the desert truffle Terfezia pfeilii by molecular methods. Mycol. Res. 1999, 103, 1270–1274. [Google Scholar] [CrossRef]
- Slama, A.; Fortas, Z.; Boudabous, A.; Neffati, M. Cultivation of an edible desert truffle (Terfezia boudieri Chatin). Afr. J. Microbio. Res. 2010, 4, 2350–2356. [Google Scholar]
- Dexheimer, J.; Gerard, J.; Leduc, J.P.; Chevalier, G. Etude ultrastructurale comparee des associations symbiotiques mycorrhiziennes Helianthemum salicifolium-Terfezia claveryi et Helianthemum salicifolium-Terfezia leptoderma. Can. J. Bot. 1985, 63, 582–591. [Google Scholar] [CrossRef]
- Roth-Bejerano, N.; Livne, D.; Kagan-Zur, V. Helianthemum-Terfezia relations in different growth media. New Phytol. 1990, 114, 235–238. [Google Scholar] [CrossRef]
- Navarro-Ródenas, A.; Pérez-Gilabert, M.; Torrente, P.; Morte, A. The role of phosphorus in the ectendomycorrhiza continuum of desert truffle mycorrhizal plants. Mycorrhiza 2012, 22, 565–575. [Google Scholar] [CrossRef]
- Navarro-Ródenas, A.; Bárzana, G.; Nicolás, E.; Carra, A.; Schubert, A.; Morte, A. Expression analysis of aquaporins from desert truffle mycorrhizal symbiosis reveals a fine-tuned regulation under drought. Mol. Plant Microbe Interact. 2013, 26, 1068–1078. [Google Scholar] [CrossRef] [Green Version]
- Chevalier, G.; Riousset, L.; Dexheimer, J.; Dupre, C. Synthese mycorhizienne entre Terfezia leptoderma Tul. et diverses cistacés. Agronomie 1984, 4, 210–211. [Google Scholar]
- Leduc, F.P.; Dexheimer, J.; Chevalier, G. Etude ultrastructurale comparee des association de Terfezia leptoderma avec Helianthemum salicifolium, Cistus albidus, et C. salviifolius. In Mycorrhizae: Physiology and Genetics; Institut National de la Recherche Agronomique: Paris, France, 1986. [Google Scholar]
- Zaretsky, M.; Kagan-Zur, V.; Mills, D.; Roth-Bejerano, N. Analysis of mycorrhizal associations formed by Cistus incanus transformed root clones with Terfezia boudieri isolates. Plant Cell Rep. 2005, 25, 62–70. [Google Scholar] [CrossRef]
- Mattirolo, O. Gli ipogei di Sardegna e di Sicilia. Malpighia 1900, 14, 39–110. [Google Scholar]
- Bordallo, J.J.; Rodriguez, A.; Muñoz-Mohedano, J.M.; Suz, L.M.; Honrubia, M.; Morte, A. Morphological and molecular characterization of five new Terfezia species from the Iberian Peninsula. Mycotaxon 2013, 124, 189–208. [Google Scholar] [CrossRef]
- Louro, R.; Nobre, T.; Santos-Silva, C. Terfezia solaris-libera sp. nov., A New Mycorrhizal Species within the Spiny-Spored Lineages. J. Mycol. Mycol. Sci. 2020, 3, 1–13. [Google Scholar] [CrossRef]
- Nobre, T.; Gomes, L.; Rei, F. Uncovered variability in olive moth (Prays oleae) questions species monophyly. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990. [Google Scholar]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinf. 2017, bbx118, 1e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- INMG. The Climate of Portugal: Normais Climatológicas of the Region of Alentejo and Algarve, Corresponding to 1951–1980; INMG: Lisboa, Portugal, 1991. [Google Scholar]
- Rivas-Martínez, S. Advances in Geobotany-Opening Speech of the Academic Year of the Real National Academy of Pharmacy of the Year 2005; Royal National Academy of Pharmacy—Institute of Spain: Madrid, Spain, 2005. [Google Scholar]
- Louro, R.; Peixe, A.; Santos-Silva, C. New insights on Cistus salviifolius L. Micropropagation. J. Bot. Sci. 2017, 6, 10–14. [Google Scholar]
- Agerer, R. Colour Atlas of Ectomycorrhizae; Einhorn Verlag: Schwabisch Gmünd, Germany, 1987–2002. [Google Scholar]
- Agerer, R. Characterization of ectomycorrhiza. In Techniques for the Study of Mycorrhiza; Norris, J.R., Read, D.J., Varma, A.K., Eds.; Academic Press: London, UK, 1991. [Google Scholar]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Brundrett, M.; Bougher, N.; Dell, B.; Grove, T.; Malajczuk, N. Working with Mycorrhizas in Forestry and Agriculture; ACIAR Monograph 32; Australian Centre for International Agricultural Research: Canberra, Australia, 1996.
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. Mesure du taux de mycorhization VA d’un système radiculaire. Recherche de méthodes d’estimation ayant une signification fonctionnelle. In Physiological and Genetical Aspects of Mycorrhizae; Gianinazzi-Pearson, V., Gianinazzi, S., Eds.; Institut National de la Recherche Agronomique: Paris, France, 1986. [Google Scholar]
- IBM Corp. IBM SPSS Statistics for Windows; Version 24.0.; IBM Corp.: Armonk, NY, USA, 2016. [Google Scholar]
- Zambonelli, A.; Iotti, M.; Hall, I.R. Current status of truffle cultivation: Recent results and future perspectives. Ital. J. Mycol. 2015, 44, 31–40. [Google Scholar]
- Morte, A.; Andrino, A.; Honrubia, M.; Navarro-Ródenas, A. Terfezia cultivation in arid and semiarid soils. In Edible Ectomycorrhizal Mushrooms; Zambonelli, A., Bonito, G.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Santos-Silva, C.; Louro, R. Desert truffle cultivation in acid soils – Terfezia arenaria mass production. New Phytol. unpublished, manuscript in preparation.






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Louro, R.; Natário, B.; Santos-Silva, C. Morphological Characterization of the In Vitro Mycorrhizae Formed between Four Terfezia Species (Pezizaceae) with Cistus salviifolius and Cistus ladanifer—Towards Desert Truffles Production in Acid Soils. J. Fungi 2021, 7, 35. https://doi.org/10.3390/jof7010035
Louro R, Natário B, Santos-Silva C. Morphological Characterization of the In Vitro Mycorrhizae Formed between Four Terfezia Species (Pezizaceae) with Cistus salviifolius and Cistus ladanifer—Towards Desert Truffles Production in Acid Soils. Journal of Fungi. 2021; 7(1):35. https://doi.org/10.3390/jof7010035
Chicago/Turabian StyleLouro, Rogério, Bruno Natário, and Celeste Santos-Silva. 2021. "Morphological Characterization of the In Vitro Mycorrhizae Formed between Four Terfezia Species (Pezizaceae) with Cistus salviifolius and Cistus ladanifer—Towards Desert Truffles Production in Acid Soils" Journal of Fungi 7, no. 1: 35. https://doi.org/10.3390/jof7010035
APA StyleLouro, R., Natário, B., & Santos-Silva, C. (2021). Morphological Characterization of the In Vitro Mycorrhizae Formed between Four Terfezia Species (Pezizaceae) with Cistus salviifolius and Cistus ladanifer—Towards Desert Truffles Production in Acid Soils. Journal of Fungi, 7(1), 35. https://doi.org/10.3390/jof7010035