Fungal Treatment for the Valorization of Technical Soda Lignin

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Soda Lignin
2.2. Culture Conditions
2.2.1. Cultures on Lignin-Containing Plates
2.2.2. Mycelia Preparation for Liquid Cultures
2.2.3. Liquid Cultures on Lignin
2.3. DNA Extraction and Mycelial Dry Weight Determination on Lignin
2.4. Enzyme Activity
2.5. Proteomic Analysis
2.6. Analysis of the Soluble Lignin Fraction
2.6.1. Sample Preparation
2.6.2. HPSEC Analysis
2.6.3. LC–MS Analysis
2.7. Analysis of Insoluble Lignin Fraction
2.7.1. HPSEC Analysis
2.7.2. Quantitative 31P NMR and Samples Preparation
2.7.3. Thioacidolysis
3. Results and Discussion
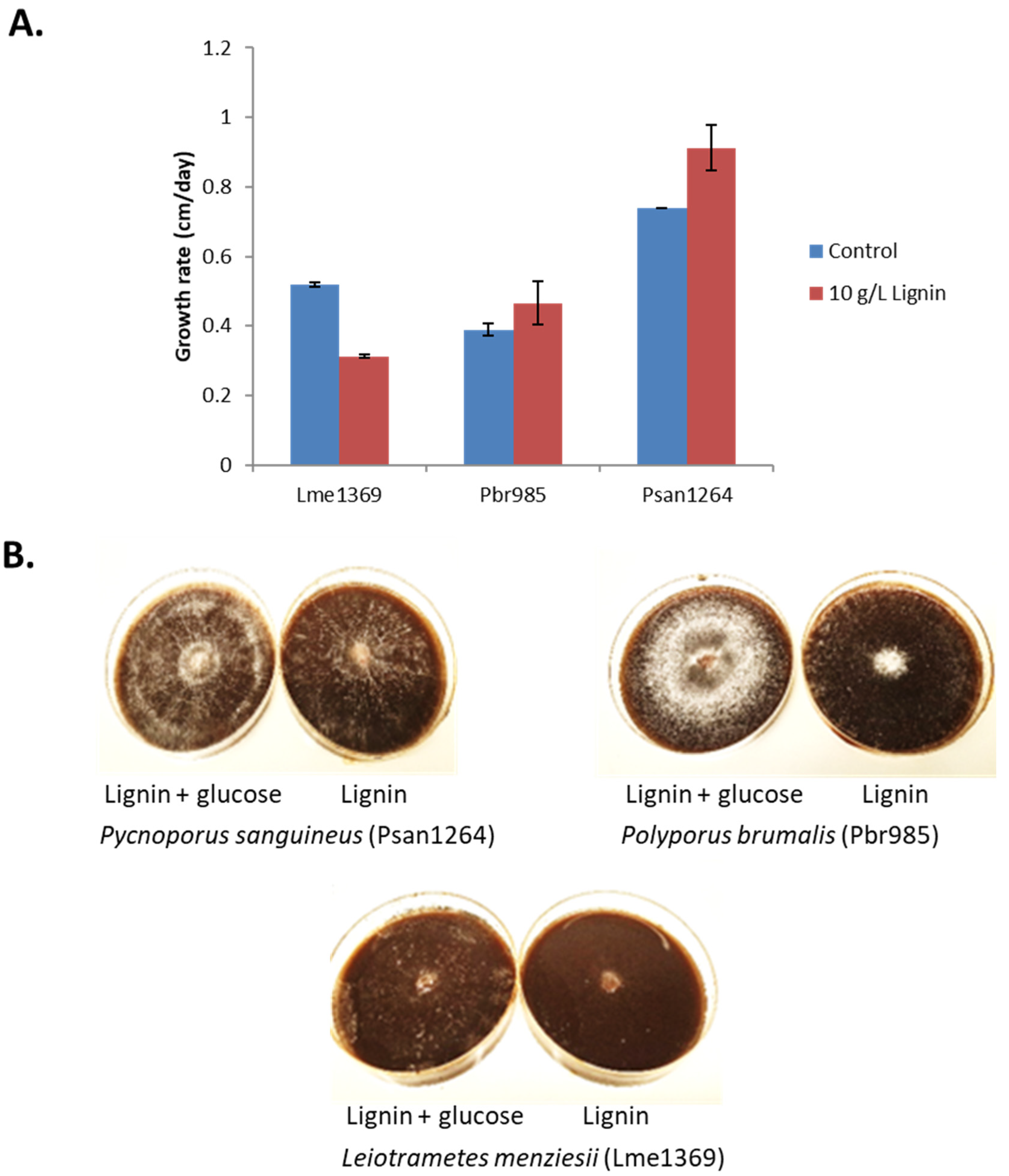
3.1. Growth in the Presence of Lignin
3.1.1. Growth on Lignin-Containing Plates
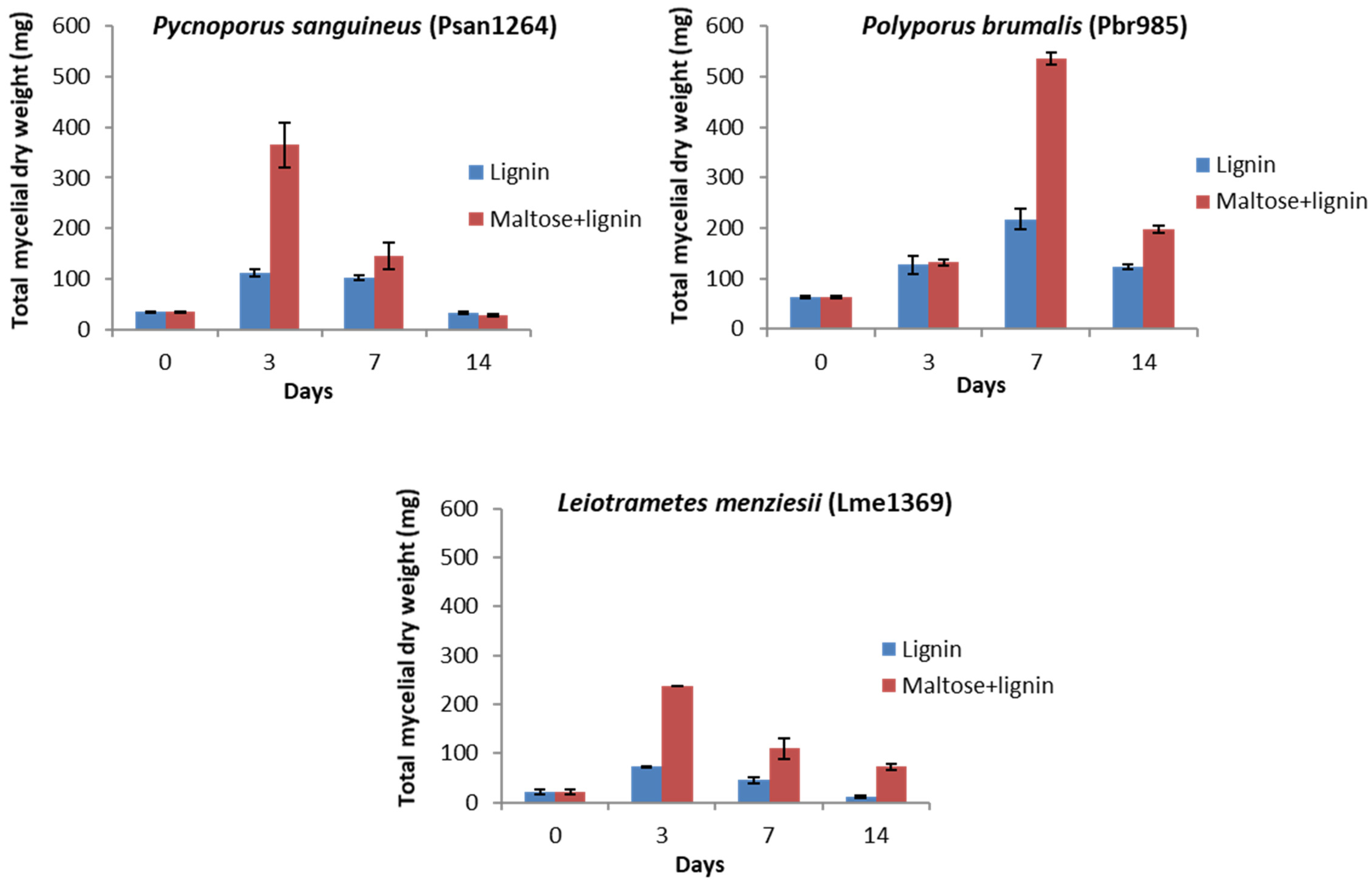
3.1.2. Liquid Cultures and Enzyme Activities
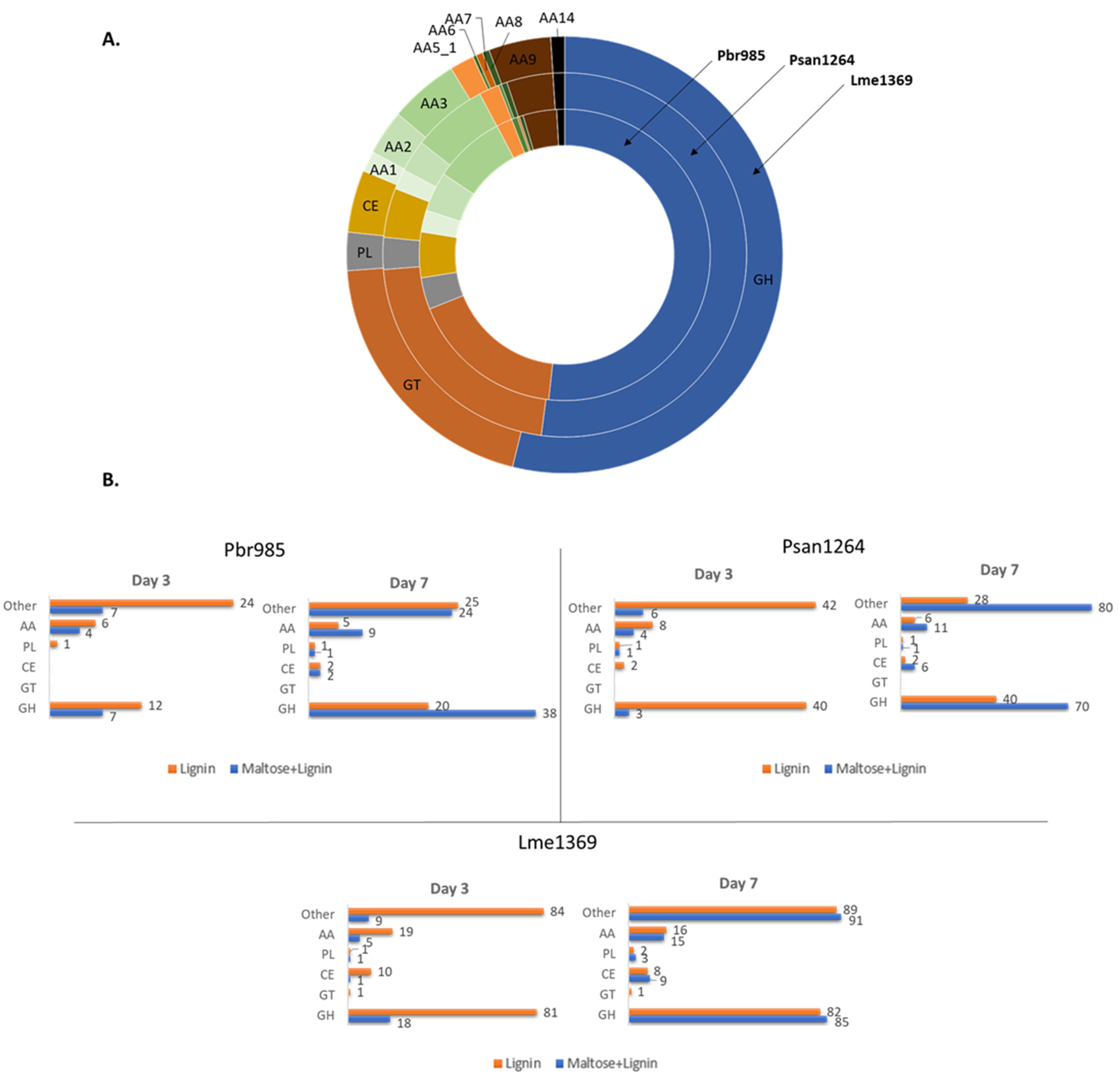
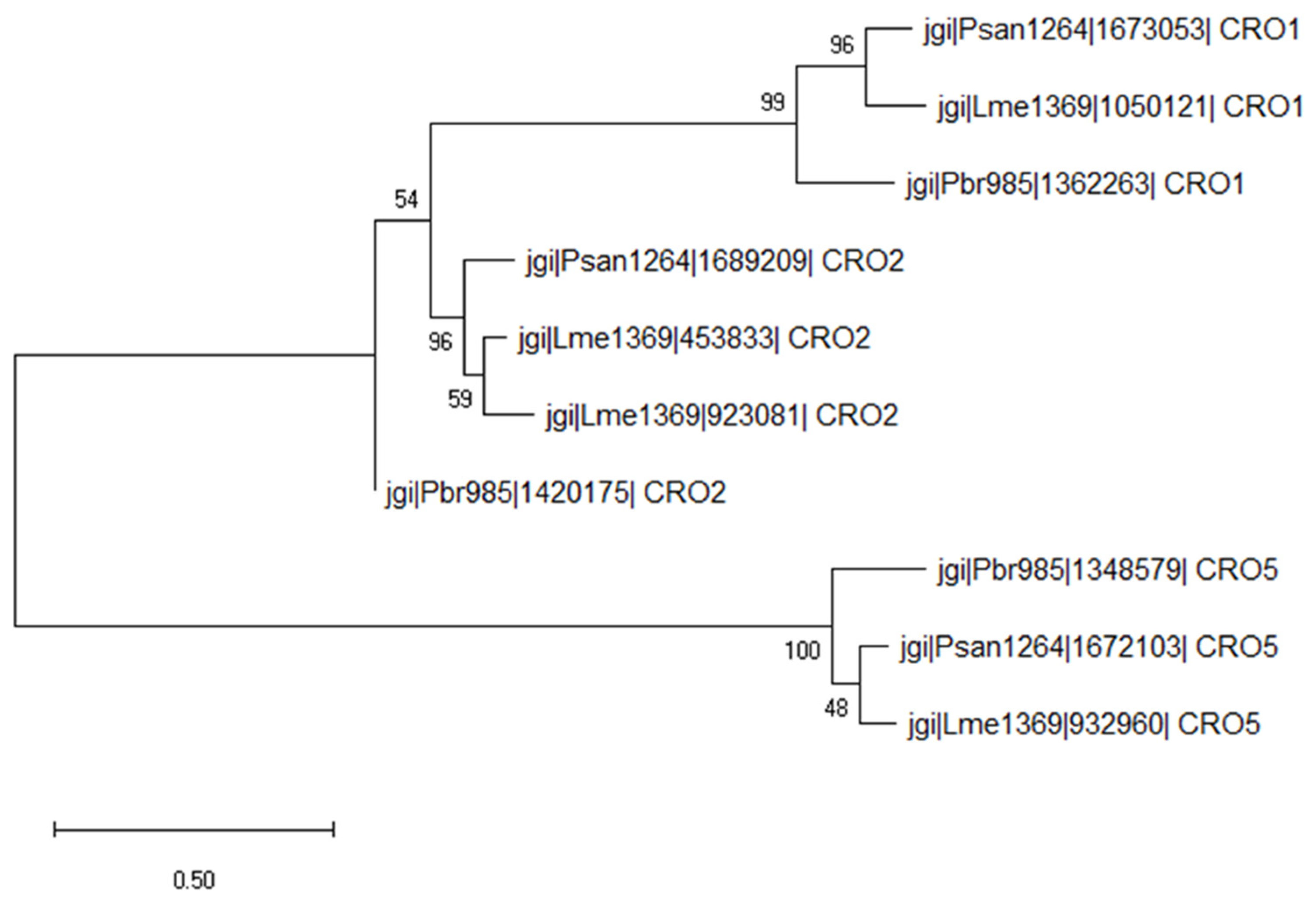
3.2. Proteomic Analysis
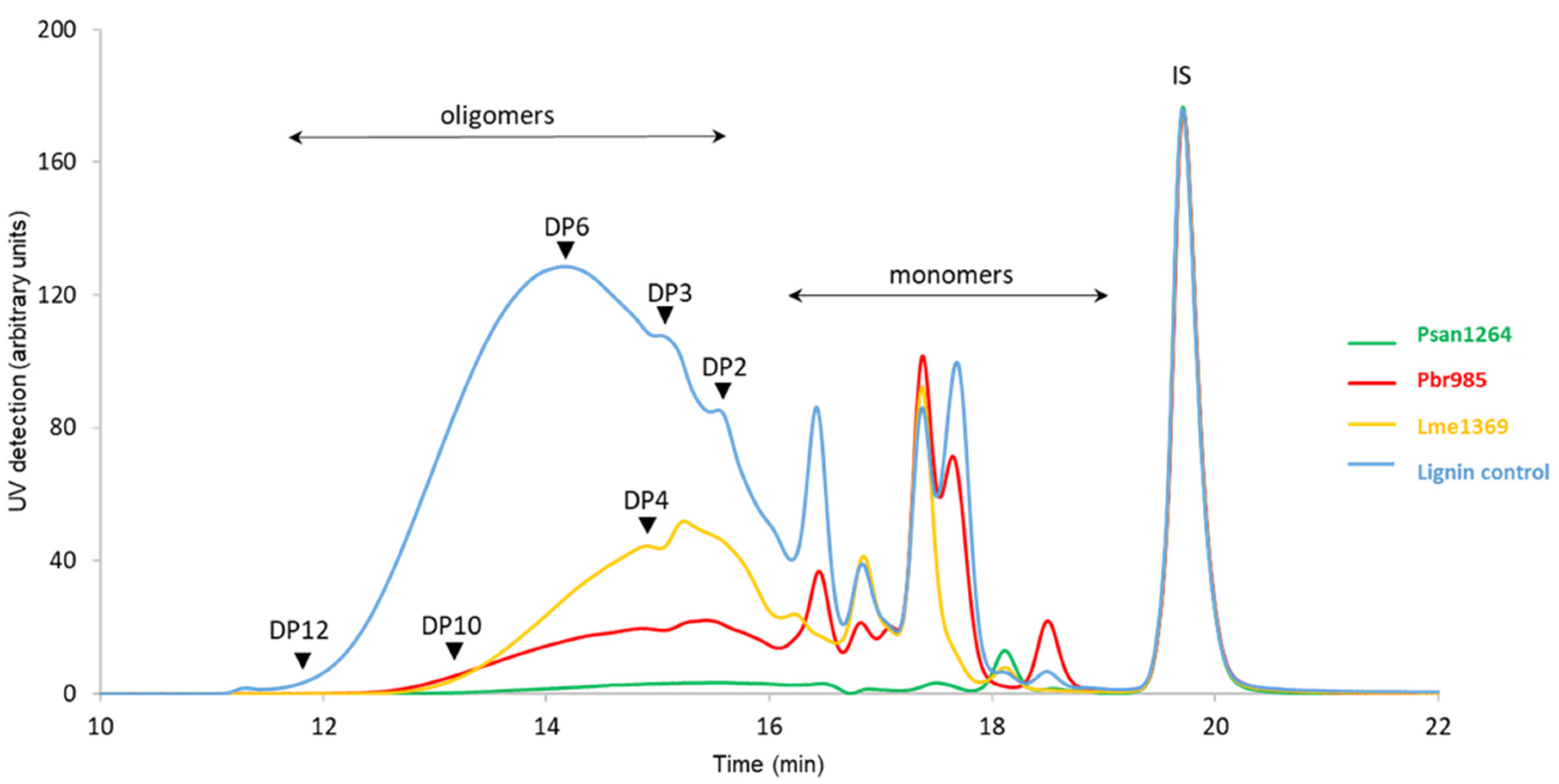
3.3. Analysis of Soluble Lignin Fraction after Fungal Treatment
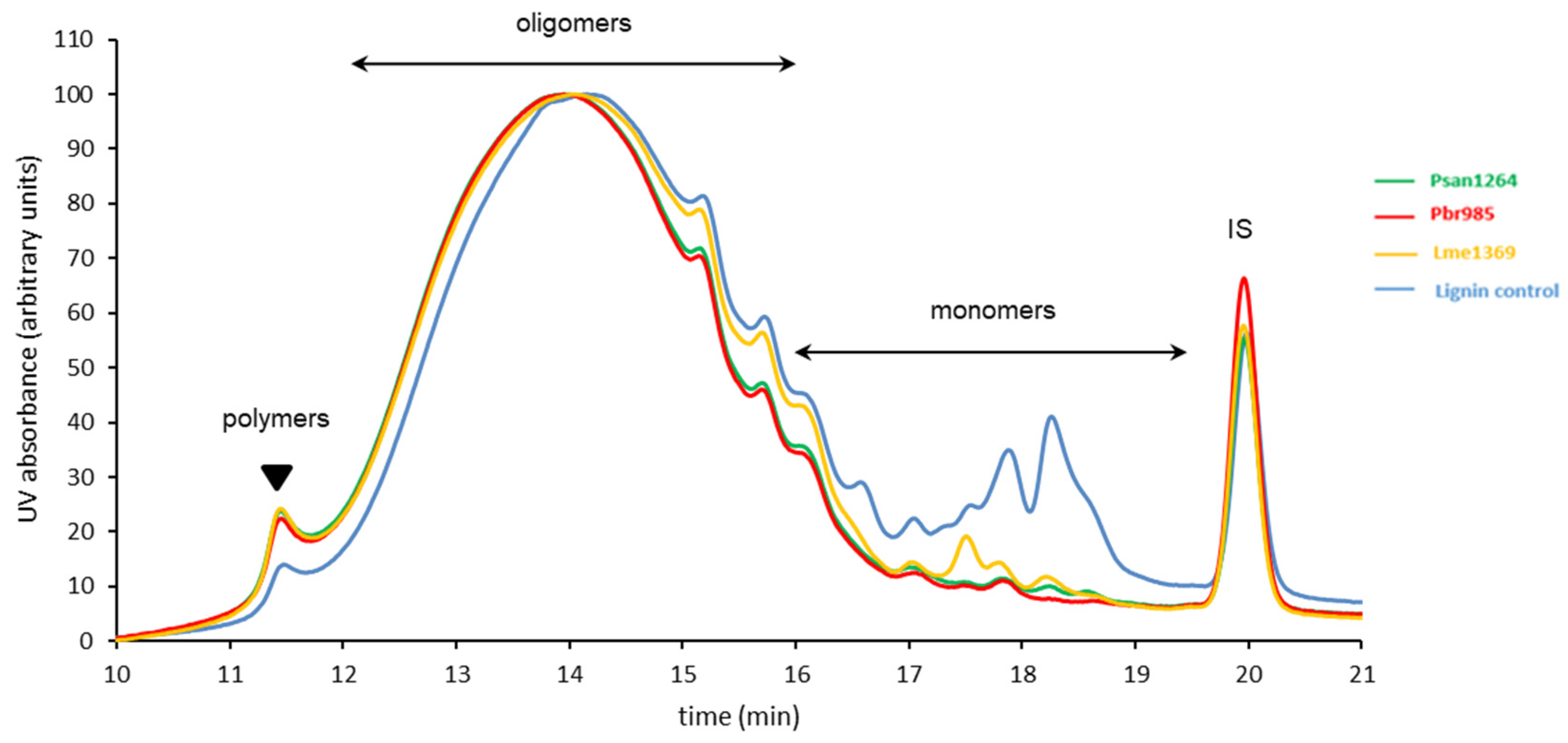
3.4. Analysis of Insoluble Residual Lignin
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Isikgor, F.H.; Becer, C.R. Lignocellulosic biomass: A sustainable platform for the production of bio-based chemicals and polymers. Polym. Chem. 2015, 6, 4497–4559. [Google Scholar] [CrossRef] [Green Version]
- Felby, C. Biomass Recalcitrance Deconstructing the Plant Cell Wall for Bioenergy. Edited by Michael E. Himmel. ChemSusChem 2009, 2, 593. [Google Scholar] [CrossRef]
- Baig, K.S.; Wu, J.; Turcotte, G. Future prospects of delignification pretreatments for the lignocellulosic materials to produce second generation bioethanol. Int. J. Energy Res. 2019, 43, 1411–1427. [Google Scholar] [CrossRef]
- Bajwa, D.S.; Pourhashem, G.; Ullah, A.H.; Bajwa, S.G. A concise review of current lignin production, applications, products and their environmental impact. Ind. Crops Prod. 2019, 139, 111526. [Google Scholar] [CrossRef]
- Schutyser, W.; Renders, T.; Van den Bosch, S.; Koelewijn, S.F.; Beckham, G.T.; Sels, B.F. Chemicals from lignin: An interplay of lignocellulose fractionation, depolymerisation, and upgrading. Chem. Soc. Rev. 2018, 47, 852–908. [Google Scholar] [CrossRef] [PubMed]
- Stéphanie Laurichesse, L.A. Chemical modification of lignins: Towards biobased polymers. Prog. Polym. Sci. 2014, 39, 1266–1290. [Google Scholar] [CrossRef]
- Ragauskas, A.J.; Beckham, G.T.; Biddy, M.J.; Chandra, R.; Chen, F.; Davis, M.F.; Davison, B.H.; Dixon, R.A.; Gilna, P.; Keller, M.; et al. Lignin Valorization: Improving Lignin Processing in the Biorefinery. Science 2014, 344, 6185. [Google Scholar] [CrossRef] [PubMed]
- Boyle, C.D.; Kropp, B.R.; Reid, I.D. Solubilization and Mineralization of Lignin by White Rot Fungi. Appl. Environ. Microbiol. 1992, 58, 3217. [Google Scholar] [CrossRef] [Green Version]
- Salvachúa, D.; Katahira, R.; Cleveland, N.S.; Khanna, P.; Resch, M.G.; Black, B.A.; Purvine, S.O.; Zink, E.M.; Prieto, A.; Martínez, M.J.; et al. Lignin depolymerization by fungal secretomes and a microbial sink. Green Chem. 2016, 18, 6046–6062. [Google Scholar] [CrossRef] [Green Version]
- Van Erven, G.; Nayan, N.; Sonnenberg, A.S.M.; Hendriks, W.H.; Cone, J.W.; Kabel, M.A. Mechanistic insight in the selective delignification of wheat straw by three white-rot fungal species through quantitative (13)C-IS py-GC-MS and whole cell wall HSQC NMR. Biotechnol. Biofuels 2018, 11, 262. [Google Scholar] [CrossRef]
- Pollegioni, L.; Tonin, F.; Rosini, E. Lignin-degrading enzymes. FEBS J. 2015, 282, 1190–1213. [Google Scholar] [CrossRef] [PubMed]
- Lancefield, C.S.; Wienk, H.L.J.; Boelens, R.; Weckhuysen, B.M.; Bruijnincx, P.C.A. Identification of a diagnostic structural motif reveals a new reaction intermediate and condensation pathway in kraft lignin formation. Chem. Sci. 2018, 9, 6348–6360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constant, S.; Wienk, H.L.J.; Frissen, A.E.; de Peinder, P.; Boelens, R.; van Es, D.S.; Grisel, R.J.H.; Weckhuysen, B.M.; Huijgen, W.J.J.; Gosselink, R.J.A.; et al. New insights into the structure and composition of technical lignins: A comparative characterisation study. Green Chem. 2016, 18, 2651–2665. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Huang, J.; Yang, L.; Yue, F.; Lu, F. Revealing Structural Differences between Alkaline and Kraft Lignins by HSQC NMR. Ind. Eng. Chem. Res. 2019, 58, 5707–5714. [Google Scholar] [CrossRef]
- Guan, X.; Zhao, H.; Xu, Y.; Wang, Y. Transient expression of glyoxal oxidase from the Chinese wild grape Vitis pseudoreticulata can suppress powdery mildew in a susceptible genotype. Protoplasma 2011, 248, 415–423. [Google Scholar] [CrossRef]
- Zhou, S.; Raouche, S.; Grisel, S.; Navarro, D.; Sigoillot, J.C.; Herpoel-Gimbert, I. Solid-state fermentation in multi-well plates to assess pretreatment efficiency of rot fungi on lignocellulose biomass. Microb. Biotechnol. 2015, 8, 940–949. [Google Scholar] [CrossRef]
- Simeng, Z.; Raouche, S.; Grisel, S.; Sigoillot, J.-C.; Gimbert, I. Efficient biomass pretreatment using the White-rot Fungus Polyporus Brumalis. Fungal Genom. Biol. 2017, 7, 3. [Google Scholar]
- Miyauchi, S.; Rancon, A.; Drula, E.; Hage, H.; Chaduli, D.; Favel, A.; Grisel, S.; Henrissat, B.; Herpoël-Gimbert, I.; Ruiz-Dueñas, F.J.; et al. Integrative visual omics of the white-rot fungus Polyporus brumalis exposes the biotechnological potential of its oxidative enzymes for delignifying raw plant biomass. Biotechnol. Biofuels 2018, 11, 201. [Google Scholar] [CrossRef] [Green Version]
- Aracri, E.; Díaz Blanco, C.; Tzanov, T. An enzymatic approach to develop a lignin-based adhesive for wool floor coverings. Green Chem. 2014, 16, 2597–2603. [Google Scholar] [CrossRef]
- Li, H. Fractionation and characterization of industrial lignins. Ind. Crops Prod 2014, 62, 67–76. [Google Scholar] [CrossRef]
- Majira, A.; Godon, B.; Foulon, L.; van der Putten, J.C.; Cézard, L.; Thierry, M.; Pion, F.; Bado-Nilles, A.; Pandard, P.; Jayabalan, T.; et al. Enhancing the Antioxidant Activity of Technical Lignins by Combining Solvent Fractionation and Ionic-Liquid Treatment. ChemSusChem 2019, 12, 4799–4809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesage-Meessen, L.; Haon, M.; Uzan, E.; Levasseur, A.; Piumi, F.; Navarro, D.; Taussac, S.; Favel, A.; Lomascolo, A. Phylogeographic relationships in the polypore fungus Pycnoporus inferred from molecular data. FEMS Microbiol. Lett. 2011, 325, 37–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, D.; Favel, A.; Chabrol, O.; Pontarotti, P.; Haon, M.; Lesage-Meessen, L. FunGene-DB: A web-based tool for Polyporales strains authentication. J. Biotechnol. 2012, 161, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Sayers, E.W.; Agarwala, R.; Bolton, E.E.; Brister, J.R.; Canese, K.; Clark, K.; Connor, R.; Fiorini, N.; Funk, K.; Hefferon, T.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2018, 47, D23–D28. [Google Scholar] [CrossRef] [Green Version]
- Welti, S.; Moreau, P.-A.; Favel, A.; Courtecuisse, R.; Haon, M.; Navarro, D.; Taussac, S.; Lesage-Meessen, L. Molecular phylogeny of Trametes and related genera, and description of a new genus Leiotrametes. Fungal Divers. 2012, 55, 47–64. [Google Scholar] [CrossRef] [Green Version]
- Dence, C.W. The Determination of Lignin. In Methods in Lignin Chemistry; Lin, S.Y., Dence, C.W., Eds.; Springer: Berlin/Heidelberg, Germany, 1992; pp. 33–61. [Google Scholar]
- Sipponen, M.H.; Lapierre, C.; Mechin, V.; Baumberger, S. Isolation of structurally distinct lignin-carbohydrate fractions from maize stem by sequential alkaline extractions and endoglucanase treatment. Bioresour. Technol. 2013, 133, 522–528. [Google Scholar] [CrossRef] [Green Version]
- Lapierre, C.; Voxeur, A.; Karlen, S.D.; Helm, R.F.; Ralph, J. Evaluation of Feruloylated and p-Coumaroylated Arabinosyl Units in Grass Arabinoxylans by Acidolysis in Dioxane/Methanol. J. Agric. Food Chem. 2018, 66, 5418–5424. [Google Scholar] [CrossRef]
- Zhou, S.; Grisel, S.; Herpoël-Gimbert, I.; Rosso, M.N. A PCR-based method to quantify fungal growth during pretreatment of lignocellulosic biomass. J. Microbiol. Methods 2015, 115, 67–70. [Google Scholar]
- Bourbonnais, R.; Paice, M.G.; Reid, I.D.; Lanthier, P.; Yaguchi, M. Lignin oxidation by laccase isozymes from Trametes versicolor and role of the mediator 2,2’-azinobis(3-ethylbenzthiazoline-6-sulfonate) in kraft lignin depolymerization. Appl. Environ. Microbiol. 1995, 61, 1876. [Google Scholar] [CrossRef] [Green Version]
- Kirk, T.K.; Croan, S.; Tien, M.; Murtagh, K.E.; Farrell, R.L. Production of multiple ligninases by Phanerochaete chrysosporium: Effect of selected growth conditions and use of a mutant strain. Enzyme Microb. Technol. 1986, 8, 27–32. [Google Scholar] [CrossRef]
- Paszczyński, A.; Huynh, V.-B.; Crawford, R. Enzymatic activities of an extracellular, manganese-dependent peroxidase from Phanerochaete chrysosporium. FEMS Microbiol. Lett. 1985, 29, 37–41. [Google Scholar] [CrossRef]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F.; et al. MycoCosm portal: Gearing up for 1000 fungal genomes. Nucleic Acids Res. 2013, 42, D699–D704. [Google Scholar] [CrossRef] [PubMed]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- International Symposium on Wood and Pulping. In Proceedings of the 12th International Symposium on Wood and Pulping Chemistry, Madison, WI, USA, 9–12 June 2003; Department of Forest Ecology and Management, University of Wisconsin-Madison: Madison, WI, USA, 2003.
- Granata, A.; Argyropoulos, D.S. 2-Chloro-4,4,5,5-tetramethyl-1,3,2-dioxaphospholane, a Reagent for the Accurate Determination of the Uncondensed and Condensed Phenolic Moieties in Lignins. J. Agric. Food Chem. 1995, 43, 1538–1544. [Google Scholar] [CrossRef]
- Lapierre, C. Application of New Methods for the Investigation of Lignin Structure. In Forage Cell Wall Struct Dig; Jung, H.G., Buxton, D.R., Hatfield, R.D., Ralph, J., Eds.; Wiley: Madison, WI, USA, 1993; Chapter 6; pp. 133–166. [Google Scholar]
- Fricker, M.; Boddy, L.; Bebber, D. Network Organisation of Mycelial Fungi. In Biology of the Fungal Cell; Springer: Berlin/Heidelberg, Germany, 2007; Volume 8, pp. 309–330. [Google Scholar]
- Sullivan, G.; Henry, E.D. Occurrence and distribution of phenoxazinone pigments in the genus Pycnoporus. J. Pharm Sci. 1971, 60, 1097–1098. [Google Scholar] [CrossRef]
- Eggert, C.; Temp, U.; Dean, J.F.; Eriksson, K.E. Laccase-mediated formation of the phenoxazinone derivative, cinnabarinic acid. FEBS Lett. 1995, 376, 202–206. [Google Scholar] [CrossRef] [Green Version]
- Thurston, C.F. The structure and function of fungal laccases. Microbiology 1994, 140, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Xie, N.; Chapeland-Leclerc, F.; Silar, P.; Ruprich-Robert, G. Systematic gene deletions evidences that laccases are involved in several stages of wood degradation in the filamentous fungus Podospora anserina. Environ. Microbiol. 2014, 16, 141–161. [Google Scholar] [CrossRef]
- Elisashvili, V.; Kachlishvili, E.; Penninckx, M. Effect of growth substrate, method of fermentation, and nitrogen source on lignocellulose-degrading enzymes production by white-rot basidiomycetes. J. Ind. Microbiol. Biotechnol. 2008, 35, 1531–1538. [Google Scholar] [CrossRef] [PubMed]
- Bi, R.; Spadiut, O.; Brumer, H.; Henriksson, G. Isolation and identification of microorganisms from soil able to live on lignin as acarbon source and to produce enzymes which cleave the β-o-4 bond in a lignin model compound. Cell Chem. Technol. 2012, 46, 227–242. [Google Scholar]
- Bi, R.; Lawoko, M.; Henriksson, G. Phoma herbarum, a soil fungus able to grow on natural lignin and synthetic lignin (DHP) as sole carbon source and cause lignin degradation. J. Ind. Microbiol. Biotechnol. 2016, 43, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Bi, R.; Huang, S.; Henriksson, G. Isolation of exceedingly low oxygen consuming fungal strains able to utilize lignin as carbon source. Cell Chem. Technol. 2016, 50, 811–817. [Google Scholar]
- Moore, D.; Robson, G.D.; Trinci, A.P.J. 21st Century Guidebook to Fungi; University Press: Cambridge, UK, 2000. [Google Scholar]
- Eggert, C.; Temp, U.; Eriksson, K.-E. The ligninolytic system of the white rot fungus Pycnoporus cinnabarinus: Purification and characterization of the laccase. Appl. Environ. Microbiol. 1996, 62, 1151–1158. [Google Scholar] [CrossRef] [Green Version]
- Pointing, S.B.; Jones, E.B.G.; Vrijmoed, L.L.P. Optimization of Laccase Production by Pycnoporus sanguineus in Submerged Liquid Culture. Mycologia 2000, 92, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Esposito, E.; Innocentini-Mei, L.H.; Ferraz, A.; Canhos, V.P.; Durán, N. Phenoloxidases and hydrolases from Pycnoporus sanguineus (UEC-2050 strain): Applications. J. Biotechnol. 1993, 29, 219–228. [Google Scholar] [CrossRef]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [Green Version]
- Levasseur, A.; Drula, E.; Lombard, V.; Coutinho, P.; Henrissat, B. Expansion of the enzymatic repertoire of the CAZy database to integrate auxiliary redox enzymes. Biotechnol. Biofuels 2013, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Gasch, A.P. Comparative genomics of the environmental stress response in ascomycete fungi. Yeast 2007, 24, 961–976. [Google Scholar] [CrossRef]
- Janusz, G.; Pawlik, A.; Sulej, J.; Swiderska-Burek, U.; Jarosz-Wilkolazka, A.; Paszczynski, A. Lignin degradation: Microorganisms, enzymes involved, genomes analysis and evolution. FEMS Microbiol. Rev. 2017, 41, 941–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanden Wymelenberg, A.; Sabat, G.; Mozuch, M.; Kersten, P.J.; Cullen, D.; Blanchette, R.A. Structure, organization, and transcriptional regulation of a family of copper radical oxidase genes in the lignin-degrading basidiomycete Phanerochaete chrysosporium. Appl. Environ. Microbiol. 2006, 72, 4871–4877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kersten, P.; Cullen, D. Copper radical oxidases and related extracellular oxidoreductases of wood-decay Agaricomycetes. Fungal Genet. Biol. 2014, 72, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Daou, M.; Piumi, F.; Cullen, D.; Record, E.; Faulds, C.B. Heterologous Production and Characterization of Two Glyoxal Oxidases from Pycnoporus cinnabarinus. Appl. Environ. Microbiol. 2016, 82, 4867–4875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adav, S.S.; Ravindran, A.; Sze, S.K. Quantitative proteomic analysis of lignocellulolytic enzymes by Phanerochaete chrysosporium on different lignocellulosic biomass. J. Proteom 2012, 75, 1493–1504. [Google Scholar] [CrossRef] [PubMed]
- Kersten, P.J. Glyoxal oxidase of Phanerochaete chrysosporium: Its characterization and activation by lignin peroxidase. Proc. Natl. Acad. Sci. USA 1990, 87, 2936–2940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, D.; Urresti, S.; Lafond, M.; Johnston, E.M.; Derikvand, F.; Ciano, L.; Berrin, J.-G.; Henrissat, B.; Walton, P.H.; Davies, G.J.; et al. Structure-function characterization reveals new catalytic diversity in the galactose oxidase and glyoxal oxidase family. Nat. Commun. 2015, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Beeson, W.T.; Vu, V.V.; Span, E.A.; Phillips, C.M.; Marletta, M.A. Cellulose Degradation by Polysaccharide Monooxygenases. Annu. Rev. Biochem. 2015, 84, 923–946. [Google Scholar] [CrossRef]
- Kittl, R.; Kracher, D.; Burgstaller, D.; Haltrich, D.; Ludwig, R. Production of four Neurospora crassa lytic polysaccharide monooxygenases in Pichia pastoris monitored by a fluorimetric assay. Biotechnol. Biofuels 2012, 5, 79. [Google Scholar] [CrossRef] [Green Version]
- Kracher, D.; Scheiblbrandner, S.; Felice, A.K.G.; Breslmayr, E.; Preims, M.; Ludwicka, K.; Haltrich, D.; Eijsink, V.G.H.; Ludwig, R. Extracellular electron transfer systems fuel cellulose oxidative degradation. Science 2016, 352, 1098. [Google Scholar] [CrossRef]
- Li, F.; Ma, F.; Zhao, H.; Zhang, S.; Wang, L.; Zhang, X.; Yu, H. A Lytic Polysaccharide Monooxygenase from a White-Rot Fungus Drives the Degradation of Lignin by a Versatile Peroxidase. Appl. Environ. Microbiol. 2019, 85, e02803–e02818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belinky, P.A.; Flikshtein, N.; Lechenko, S.; Gepstein, S.; Dosoretz, C.G. Reactive oxygen species and induction of lignin peroxidase in Phanerochaete chrysosporium. Appl. Environ. Microbiol. 2003, 69, 6500–6506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levasseur, A.; Lomascolo, A.; Chabrol, O.; Ruiz-Duenas, F.J.; Boukhris-Uzan, E.; Piumi, F.; Kues, U.; Ram, A.F.; Murat, C.; Haon, M.; et al. The genome of the white-rot fungus Pycnoporus cinnabarinus: A basidiomycete model with a versatile arsenal for lignocellulosic biomass breakdown. BMC Genom. 2014, 15, 486. [Google Scholar] [CrossRef] [PubMed]
- Arantes, V.; Goodell, B. Current Understanding of Brown-Rot Fungal Biodegradation Mechanisms: A Review. In Deterioration and Protection of Sustainable Biomaterials; American Chemical Society: Washington, DC, USA, 2014; Volume 1158, pp. 3–21. [Google Scholar]
- Niemenmaa, O.; Uusi-Rauva, A.; Hatakka, A. Demethoxylation of [O14CH3]-labelled lignin model compounds by the brown-rot fungi Gloeophyllum trabeum and Poria (Postia) placenta. Biodegradation 2007, 19, 555. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Griffin, K.; Langeveld, S.; Frommhagen, M.; Underlin, E.N.; Kabel, M.A.; de Vries, R.P.; Dilokpimol, A. Functional Validation of Two Fungal Subfamilies in Carbohydrate Esterase Family 1 by Biochemical Characterization of Esterases From Uncharacterized Branches. Front. Bioeng Biotechnol. 2020, 8, 694. [Google Scholar] [CrossRef]
- Jeffries, T.W. Biodegradation of lignin-carbohydrate complexes. Biodegradation 1990, 1, 163–176. [Google Scholar] [CrossRef]
- McSweeney, C.S.; Dulieu, A.; Katayama, Y.; Lowry, J.B. Solubilization of lignin by the ruminal anaerobic fungus Neocallimastix patriciarum. Appl. Environ. Microbiol. 1994, 60, 2985. [Google Scholar] [CrossRef] [Green Version]
- Biondo, C.; Beninati, C.; Delfino, D.; Oggioni, M.; Mancuso, G.; Midiri, A.; Bombaci, M.; Tomaselli, G.; Teti, G. Identification and cloning of a cryptococcal deacetylase that produces protective immune responses. Infect. Immun. 2002, 70, 2383–2391. [Google Scholar] [CrossRef] [Green Version]
- Mosbech, C.; Holck, J.; Meyer, A.; Agger, J.W. Enzyme kinetics of fungal glucuronoyl esterases on natural lignin-carbohydrate complexes. Appl. Microbiol. Biotechnol. 2019, 103, 4065–4075. [Google Scholar] [CrossRef] [Green Version]
- Christgau, S.; Kofod, L.V.; Halkier, T.; Andersen, L.N.; Hockauf, M.; Dörreich, K.; Dalbøge, H.; Kauppinen, S. Pectin methyl esterase from Aspergillus aculeatus: Expression cloning in yeast and characterization of the recombinant enzyme. Biochem. J. 1996, 319, 705–712. [Google Scholar] [CrossRef] [Green Version]
- Moraes, E.C.; Alvarez, T.M.; Persinoti, G.F.; Tomazetto, G.; Brenelli, L.B.; Paixão, D.A.A.; Ematsu, G.C.; Aricetti, J.A.; Caldana, C.; Dixon, N.; et al. Lignolytic-consortium omics analyses reveal novel genomes and pathways involved in lignin modification and valorization. Biotechnol. Biofuels 2018, 11, 75. [Google Scholar] [CrossRef] [PubMed]
- Mani, P.; Fidal Kumar, T.V.; Keshavarz, T.; Chandra, S.T.; Kyazze, G. The Role of Natural Laccase Redox Mediators in Simultaneous Dye Decolorization and Power Production in Microbial Fuel Cells. Energies 2018, 11, 3455. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Strain | Condition | Incubation Time (Day) | ||
|---|---|---|---|---|---|
| 3 | 7 | 14 | |||
| Activity (nkat/mg dry mycelia) | |||||
| ABTS Oxidation | Pbr985 | Maltose (20 g/L) | 1.47 ± 0.09 | 26.61 ± 8.05 | 7.42 ± 0.33 |
| Maltose (2.5 g/L) + Lignin (15 g/L) | 4.22 ± 0.29 | 34.94 ± 2.57 | 226.42 ± 32.72 | ||
| Lignin (15 g/L) | 0.49 ± 0.24 | 14.34 ± 1.89 | 37.36 ± 5.59 | ||
| Psan1264 | Maltose (20 g/L) | 35.56 ± 1.24 | 32.18 ± 2.79 | 25.38 ± 0.74 | |
| Maltose (2.5 g/L) + Lignin (15 g/L) | 20.74 ± 1.61 | 768.67 ± 151.03 | 3175.82 ± 320.50 | ||
| Lignin (15 g/L) | 13.97 ± 0.12 | 32.52 ± 3.26 | 112.49 ± 13.09 | ||
| Lme1369 | Maltose (20 g/L) | nd | nd | nd | |
| Maltose (2.5 g/L) + Lignin (15 g/L) | 34.16 ± 2.49 | 66.18 ± 0.93 | 73.08 ± 5.62 | ||
| Lignin (15 g/L) | nd | nd | 6.52 ± 1.31 | ||
| Veratryl Alcohol Oxidation | Pbr985 | Maltose (20 g/L) | nd | nd | 0.11 ± 0.15 |
| Maltose (2.5 g/L) + Lignin (15 g/L) | 0.97 ± 0.06 | 1.19 ± 0.09 | 2.40 ± 0.1 | ||
| Lignin (15 g/L) | nd | 1.91 ± 0.37 | nd | ||
| Psan1264 | Maltose (20 g/L) | nd | nd | nd | |
| Maltose (2.5 g/L) + Lignin (15 g/L) | nd | 0.25 ± 0.19 | 0.36 ± 0.1 | ||
| Lignin (15 g/L) | nd | 2.27 ± 0.52 | 1.97 ± 0.1 | ||
| Lme1369 | Maltose (20 g/L) | nd | nd | nd | |
| Maltose (2.5 g/L) + Lignin (15 g/L) | nd | 0.38 ± 0.04 | 1.94 ± 0.28 | ||
| Lignin (15 g/L) | nd | nd | 0.09 ± 0.1 | ||
| Pbr985 | Psan1264 | Lme1369 | ||||||
|---|---|---|---|---|---|---|---|---|
| Protein ID | CAZy | Day of Growth | Protein ID | CAZy | Day of Growth | Protein ID | CAZy | Day of Growth |
| 657909 | AA1_1 | 3 and 7 | 1583166 | AA1_1 | 3, 7 and 14 | 924031 | AA1_1 | 3 and 7 |
| 224137 | AA1_1 | 3, 7 and 14 | 1560767 | AA1_1 | 3 | 1044554 | AA1_1 | 3, 7 and 14 |
| 1420175 | AA5_1 | 3, 7 and 14 | 1672751 | AA8-AA3_1 | 3, 7 and 14 | 916190 | AA8-AA3_1 | 3, 7 and 14 |
| 1362263 | AA5_1 | 3 and 7 | 1574363 | AA3_2 | 3, 7 and 14 | 927121 | AA3_2 | 3 and 7 |
| 1348579 | AA5_1 | 3 | 1648421 | AA3_4 | 7 and 14 | 924050 * | AA3_2 | 3 |
| 1388977 | AA7 | 3, 7 and 14 | 1672103 | AA5_1 | 3, 7 and 14 | 1048304 | AA3_2 | 7 |
| 1689209 | AA5_1 | 3, 7 and 14 | 437597 * | AA3_3 | 3, 7 and 14 | |||
| 1673053 * | AA5_1 | 3 | 453833 | AA5_1 | 3, 7 and 14 | |||
| 1600019 * | AA9 | 3 | 1050121 * | AA5_1 | 3 and 7 | |||
| 1677933 * | CBM1-CE1 | 3 | 923081 | AA5_1 | 3, 7 and 14 | |||
| 1565370 | CE4 | 3 and 7 | 932960 | AA5_1 | 3 and 7 | |||
| 1577721 | CE8 | 7 and 14 | 1057183 * | AA9 | 3 and 7 | |||
| 1050920 | AA9 | 3 and 7 | ||||||
| 1050025 | AA9 | 3 and 7 | ||||||
| 497583 | AA9 | 3 | ||||||
| 95318 | AA9 | 3, 7 and 14 | ||||||
| 1046503 * | AA9 | 3 | ||||||
| 923917 * | AA9 | 3 | ||||||
| 931673 | AA9-CBM1 | 3 and 7 | ||||||
| 1106760 * | AA9-CBM1 | 3 | ||||||
| 932017 * | AA14 | 7 | ||||||
| 1042341 | CE1-CBM1 | 3 and 7 | ||||||
| 693655 * | CE1-CBM1 | 3 | ||||||
| 946461 | CE4 | 3 and 7 | ||||||
| 908440 | CE4 | 3 and 7 | ||||||
| 754929 * | CE4 | 3 | ||||||
| 1018647 | CE8 | 3, 7 and 14 | ||||||
| 46074 | CE8 | 7 and 14 | ||||||
| 921049 | CE15-CBM1 | 3 | ||||||
| 250112 | CE16 | 3 and 7 | ||||||
| 1049410 | CE16 | 3 and 7 | ||||||
| 1046069 | CE16-CBM1 | 3 and 7 | ||||||
| Rt (min) | Phenolic Compounds | Lme1369 | Psan1264 | Pbr985 |
|---|---|---|---|---|
| 1.9 | p-OH benzaldehyde | -- | - | ~ |
| 2.3 | Syringic acid | ~ | -- | -- |
| 2.8 | Vanillin | -- | - | - |
| 3.4 | p-Coumaric acid | -- | -- | - |
| 3.8 | Syringaldehyde | -- | -- | ~ |
| 4.7 | Ferulic acid | -- | -- | -- |
| 5.0 | Acetosytingone | ~ | -- | ~ |
| Strain | PheOH (mmol·g−1) a | Thioacidolysis b | |
|---|---|---|---|
| Total Yield (µmol·g−1) | S/G Ratio | ||
| PB1000 | 2.68 | 104 ± 4 | 0.97 ± 0.01 |
| Control | 2.90 | 132 ± 9 | 0.93 ± 0.05 |
| Pbr985 | 2.42 | 96 ± 1 | 0.91 ± 0.02 |
| Psan1264 | 2.48 | 79 ± 7 | 0.85 ± 0.02 |
| Lme1369 | 2.85 | 120 ± 4 | 0.90 ± 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daou, M.; Farfan Soto, C.; Majira, A.; Cézard, L.; Cottyn, B.; Pion, F.; Navarro, D.; Oliveira Correia, L.; Drula, E.; Record, E.; et al. Fungal Treatment for the Valorization of Technical Soda Lignin. J. Fungi 2021, 7, 39. https://doi.org/10.3390/jof7010039
Daou M, Farfan Soto C, Majira A, Cézard L, Cottyn B, Pion F, Navarro D, Oliveira Correia L, Drula E, Record E, et al. Fungal Treatment for the Valorization of Technical Soda Lignin. Journal of Fungi. 2021; 7(1):39. https://doi.org/10.3390/jof7010039
Chicago/Turabian StyleDaou, Mariane, Clementina Farfan Soto, Amel Majira, Laurent Cézard, Betty Cottyn, Florian Pion, David Navarro, Lydie Oliveira Correia, Elodie Drula, Eric Record, and et al. 2021. "Fungal Treatment for the Valorization of Technical Soda Lignin" Journal of Fungi 7, no. 1: 39. https://doi.org/10.3390/jof7010039
APA StyleDaou, M., Farfan Soto, C., Majira, A., Cézard, L., Cottyn, B., Pion, F., Navarro, D., Oliveira Correia, L., Drula, E., Record, E., Raouche, S., Baumberger, S., & Faulds, C. B. (2021). Fungal Treatment for the Valorization of Technical Soda Lignin. Journal of Fungi, 7(1), 39. https://doi.org/10.3390/jof7010039