
Chronic Diseases Associated with Malassezia Yeast


Abstract
:1. Introduction
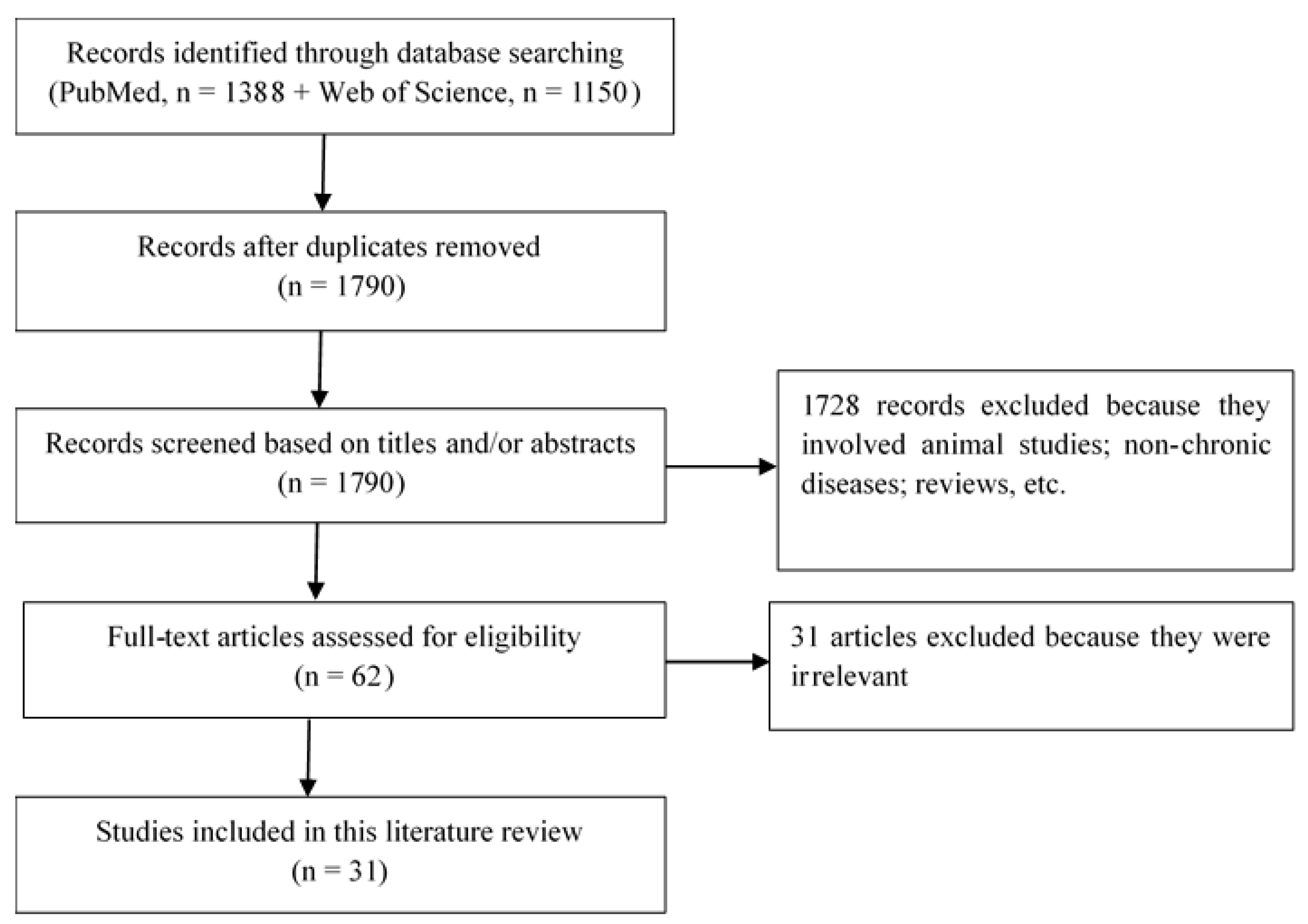
2. Materials and Methods
3. Results
3.1. Association between Malassezia and Psoriasis
3.2. Allergic Diseases Associated with Malassezia
3.2.1. Chronic Rhinosinusitis
3.2.2. Atopic Dermatitis and Asthma
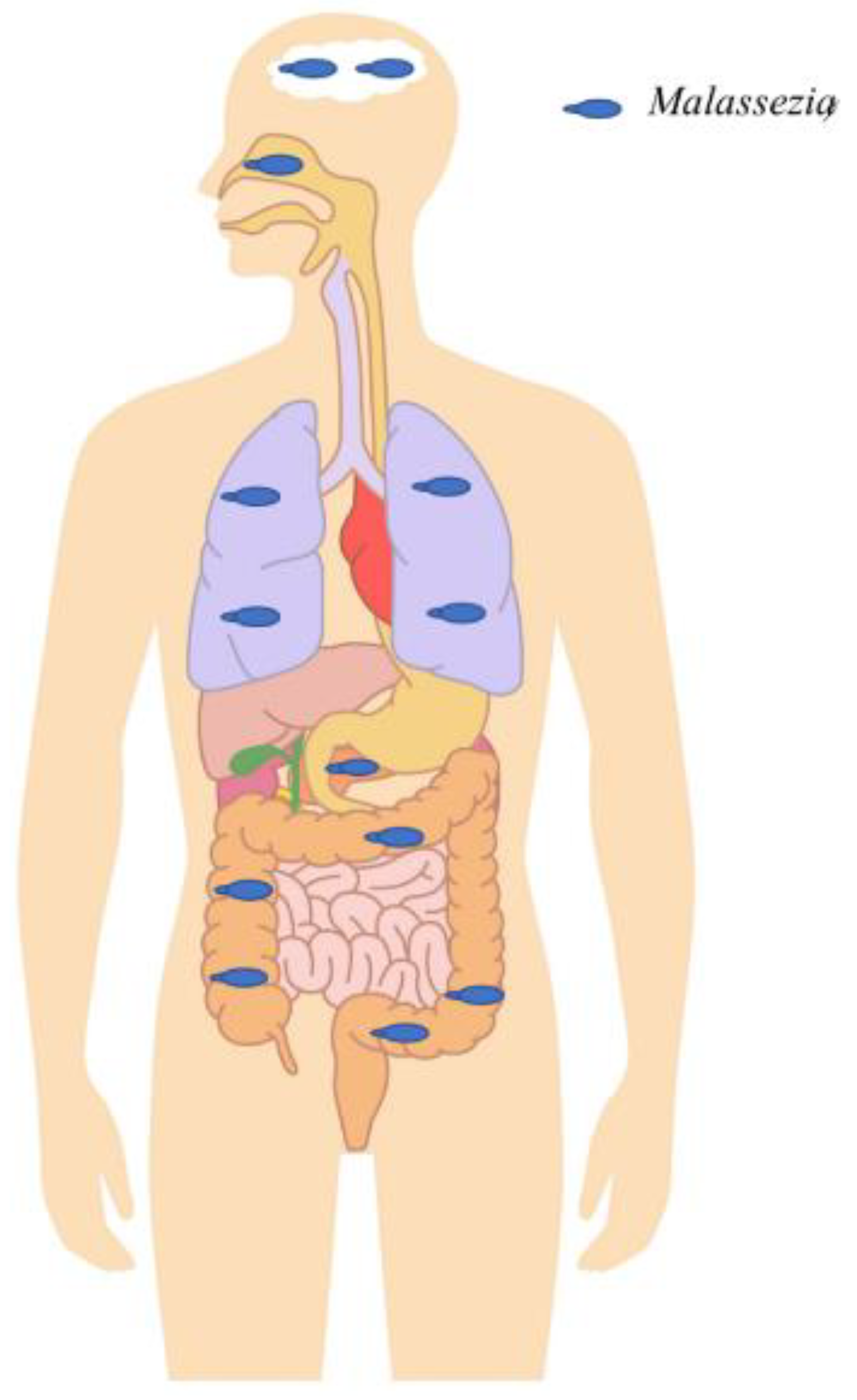
3.3. Cystic Fibrosis and Malassezia
3.4. Malassezia and HIV Infection
3.5. Malassezia in Enteric Diseases
3.5.1. Inflammatory Bowel Disease
3.5.2. Colorectal Cancer
3.6. Malassezia Neuroinfection in Neurodegenerative Diseases
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hirai, A.; Kano, R.; Makimura, K.; Duarte, E.R. Malassezia nana sp. nov., a Novel Lipid-Dependent Yeast Species Isolated from Animals. Int. J. Syst. Evol. Microbiol. 2004, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Akaza, N.; Akamatsu, H.; Sasaki, Y.; Takeoka, S.; Kishi, M.; Mizutani, H.; Sano, A.; Hirokawa, K.; Nakata, S.; Matsunaga, K. Cutaneous malassezia Microbiota of Healthy Subjects Differ by Sex, Body Part and Season. J. Dermatol. 2010, 786–792. [Google Scholar] [CrossRef] [PubMed]
- Findley, K.; Oh, J.; Yang, J.; Conlan, S.; Deming, C.; Meyer, J.A.; Schoenfeld, D.; Nomicos, E.; Park, M.; Sequencing, N.C. Human Skin Fungal Diversity. Nature 2013, 498, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Zhao, H.; Li, C.; Rajapakse, M.P.; Wong, W.C.; Xu, J.; Saunders, C.W.; Reeder, N.L.; Reilman, R.A.; Scheynius, A.; et al. Genus-Wide Comparative Genomics of Malassezia delineates Its Phylogeny, Physiology, and Niche Adaptation on Human Skin. PLoS Genet. 2015, 11, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Gaitanis, G.; Magiatis, P.; Hantschke, M.; Bassukas, I.D.; Velegraki, A. The Malassezia Genus in Skin and Systemic Diseases. Clin. Microbiol. Rev. 2012, 106–141. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.K.; Batra, R.; Bluhm, R.; Boekhout, T.; Dawson, T.L. Skin Diseases Associated with Malassezia Species. J. Am. Acad. Dermatol. 2004, 51, 785–798. [Google Scholar] [CrossRef]
- Chryssanthou, E.; Broberger, U.; Petrini, B. Malassezia pachydermatis fungaemia in a Neonatal Intensive Care Unit. Acta Paediatr. Int. J. Paediatr. 2001, 90, 323–327. [Google Scholar] [CrossRef]
- Al-Sweih, N.; Ahmad, S.; Joseph, L.; Khan, S.; Khan, Z. Malassezia pachydermatis fungemia in a Preterm Neonate Resistant to Fluconazole and Flucytosine. Med. Mycol. Case Rep. 2014, 5, 9–11. [Google Scholar] [CrossRef]
- Iatta, R.; Cafarchia, C.; Cuna, T.; Montagna, O.; Laforgia, N.; Gentile, O.; Rizzo, A.; Boekhout, T.; Otranto, D.; Montagna, M.T. Bloodstream Infections by Malassezia and Candida Species in Critical Care Patients. Med. Mycol. 2014, 264–269. [Google Scholar] [CrossRef] [Green Version]
- Dupuy, A.K.; David, M.S.; Li, L.; Heider, T.N.; Peterson, J.D.; Montano, E.A.; Dongari-Bagtzoglou, A.; Diaz, P.I.; Strausbaugh, L.D. Redefining the Human Oral Mycobiome with Improved Practices in Amplicon-Based Taxonomy: Discovery of Malassezia as a Prominent Commensal. PLoS ONE 2014, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Nash, A.K.; Auchtung, T.A.; Wong, M.C.; Smith, D.P.; Gesell, J.R.; Ross, M.C.; Stewart, C.J.; Metcalf, G.A.; Muzny, D.M.; Gibbs, R.A.; et al. The Gut Mycobiome of the Human Microbiome Project Healthy Cohort. Microbiome 2017, 5, 153. [Google Scholar] [CrossRef] [PubMed]
- Hoggard, M.; Zoing, M.; Biswas, K.; Taylor, M.W.; Douglas, R.G. The Sinonasal Mycobiota in Chronic Rhinosinusitis and Control Patients. Rhinology 2019, 57, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Pisa, D.; Alonso, R.; Carrasco, L. Parkinson’s Disease: A Comprehensive Analysis of Fungi and Bacteria in Brain Tissue. Int. J. Biol. Sci. 2020, 16, 1135–1152. [Google Scholar] [CrossRef] [PubMed]
- Boehncke, W.H.; Schön, M.P. Psoriasis. Lancet 2015, 386, 983–994. [Google Scholar] [CrossRef]
- Amaya, M.; Tajima, M.; Okubo, Y.; Sugita, T.; Nishikawa, A.; Tsuboi, R. Molecular Analysis of Malassezia Microflora in the Lesional Skin of Psoriasis Patients. J. Dermatol. 2007, 34, 619–624. [Google Scholar] [CrossRef]
- Rudramurthy, S.M.; Honnavar, P.; Chakrabarti, A.; Dogra, S.; Singh, P.; Handa, S. Association of Malassezia Species with Psoriatic Lesions. Mycoses 2014, 57, 483–488. [Google Scholar] [CrossRef]
- Gomez-Moyano, E.; Crespo-Erchiga, V.; Martínez-Pilar, L.; Godoy Diaz, D.; Martínez-García, S.; Lova Navarro, M.; Vera Casaño, A. Do Malassezia Species Play a Role in Exacerbation of Scalp Psoriasis? J. Mycol. Med. 2014, 24, 87–92. [Google Scholar] [CrossRef]
- Stehlikova, Z.; Kostovcik, M.; Kostovcikova, K.; Kverka, M.; Juzlova, K.; Rob, F.; Hercogova, J.; Bohac, P.; Pinto, Y.; Uzan, A.; et al. Dysbiosis of Skin Microbiota in Psoriatic Patients: Co-occurrence of Fungal and Bacterial Communities. Front. Microbil. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Squiquera, L.; Galimberti, R.; Morelli, L.; Plotkin, L.; Milicich, R.; Kowalckzuk, A.; Leoni, J. Antibodies to Proteins from Pityrosporum ovale in the Sera from Patients with Psoriasis. Clin. Exp. Dermatol. 1994, 19, 289–293. [Google Scholar] [CrossRef]
- Mathov, I.; Plotkin, L.; Abatangelo, C.; Galimberti, R.; Squiquera, L. Antibodies from Patients with Psoriasis Recognize N -Acetylglucosamine Terminals in Glycoproteins from Pityrosporum ovale. Clin. Exp. Immunol. 1996, 105, 79–83. [Google Scholar] [CrossRef]
- Baker, B.S.; Garjoch, A.P.J.J.; Hardman, C.; Fry, L. Differential T-Cell Reactivity to the Round and Oval Forms of Pityrosporum in the Skin of Patients with Psoriasis. Br. J. Dermatol. 1997, 136, 319–325. [Google Scholar] [CrossRef]
- Baroni, A.; Paoletti, I.; Ruocco, E.; Agozzino, M.; Tufano, M.A.; Donnarumma, G. Possible Role of Malassezia Furfur in Psoriasis: Modulation of TGF-β1, Integrin, and HSP70 Expression in Human Keratinocytes and in the Skin of Psoriasis-Affected Patients. J. Cutan. Pathol. 2004, 31, 35–42. [Google Scholar] [CrossRef]
- Kanda, N.; Tani, K.; Enomoto, U.; Nakai, K.; Watanabe, S. The Skin Fungus-Induced Th1- and Th2-Related Cytokine, Chemokine and Prostaglandin E2 Production in Peripheral Blood Mononuclear Cells from Patients with Atopic Dermatitis and Psoriasis vulgaris. Clin. Exp. Allergy 2002, 2, 1243–1250. [Google Scholar] [CrossRef]
- Lober, C.W.; Belew, P.W.; Rosenberg, E.W.; Bale, G. Patch Tests with Killed Sonicated Microflora in Patients with Psoriasis. Arch. Dermatol. 1982, 118, 322–325. [Google Scholar] [CrossRef]
- Ebbens, F.A.; Georgalas, C.; Fokkens, W.J. Fungus as the Cause of Chronic Rhinosinusitis: The Case Remains Unproven. Curr. Opin. Otolaryngol. Head Neck Surg. 2009, 17, 43–49. [Google Scholar] [CrossRef]
- Sivasubramaniam, R.; Douglas, R. The Microbiome and Chronic Rhinosinusitis. World J. Otorhinolaryngol.-Head Neck Surg. 2018, 4, 216–221. [Google Scholar] [CrossRef]
- Cleland, E.J.; Bassiouni, A.; Bassioni, A.; Boase, S.; Dowd, S.; Vreugde, S.; Wormald, P.-J. The Fungal Microbiome in Chronic Rhinosinusitis: Richness, Diversity, Postoperative Changes and Patient Outcomes. Int. Forum Allergy Rhinol. 2014, 4, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Gelber, J.T.; Cope, E.K.; Goldberg, A.N.; Pletcher, S.D. Evaluation of Malassezia and Common Fungal Pathogens in Subtypes of Chronic Rhinosinusitis. Int. Forum Allergy Rhinol. 2016, 6, 950–955. [Google Scholar] [CrossRef]
- Jung, W.H.; Croll, D.; Cho, J.H.; Kim, Y.R.; Lee, Y.W. Analysis of the Nasal Vestibule Mycobiome in Patients with Allergic Rhinitis. Mycoses 2015, 58, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Paulino, L.C.; Tseng, C.; Strober, B.E.; Blaser, M.J. Molecular Analysis of Fungal Microbiota in Samples from Healthy Human Skin and Psoriatic Lesions. J. Clin. Microbiol. 2006, 44, 2933–2941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bittar, F.; Gouriet, F.; Khelaifia, S.; Raoult, D.; Ranque, S. FastFung: A Novel Medium for the Culture and Isolation of Fastidious Fungal Species from Clinical Samples. J. Microbiol. Methods 2020, 180, 19–21. [Google Scholar] [CrossRef]
- Abdillah, A.; Khelaifia, S.; Raoult, D.; Bittar, F.; Ranque, S. Comparison of Three Skin Sampling Methods and Two Media for Culturing Malassezia Yeast. J. Fungi 2020, 6, 1–7. [Google Scholar] [CrossRef]
- Lee, K.; Zhang, I.; Kyman, S.; Kask, O.; Kathryn, E. Co-Infection of Malassezia sympodialis with Bacterial Pathobionts Pseudomonas aeruginosa or Staphylococcus aureus Leads to Distinct Sinonasal Inflammatory Responses in a Murine Acute Sinusitis Model. Front. Cell. Infect. Microbiol. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Roesner, L.M.; Ernst, M.; Chen, W.; Begemann, G.; Kienlin, P.; Raulf, M.K.; Lepenies, B.; Werfel, T. Human Thioredoxin, a Damage-Associated Molecular Pattern and Malassezia-Crossreactive Autoallergen, Modulates Immune Responses via the C-Type Lectin Receptors Dectin-1 and Dectin-2. Sci. Rep. 2019, 9, 11210. [Google Scholar] [CrossRef] [Green Version]
- Bieber, T. Atopic Dermatitis. Ann. Dermatol. 2010, 22, 125–137. [Google Scholar] [CrossRef] [Green Version]
- Sonesson, A.; Bartosik, J.; Christiansen, J.; Roscher, I.; Nilsson, F.; Schmidtchen, A.; Bäck, O. Sensitization to Skin-Associated Microorganisms in Adult Patients with Atopic Dermatitis is of Importance for Disease Severity. Acta Derm.-Venereol. 2013, 93, 340–345. [Google Scholar] [CrossRef] [Green Version]
- Kaga, M.; Sugita, T.; Nishikawa, A.; Wada, Y.; Hiruma, M.; Ikeda, S. Molecular Analysis of the Cutaneous Malassezia Microbiota from the Skin of Patients with Atopic Dermatitis of Different Severities. Mycoses 2009, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Yim, S.M.; Kim, J.Y.; Ko, J.H.; Lee, Y.W.; Choe, Y.B.; Ahn, K.J. Molecular Analysis of Malassezia Microflora on the Skin of the Patients with Atopic Dermatitis. Ann. Dermatol. 2010, 22, 41–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, E.; Tanaka, T.; Tajima, M.; Tsuboi, R.; Nishikawa, A.; Sugita, T. Characterization of the Skin Fungal Microbiota in Patients with Atopic Dermatitis and in Healthy Subjects. Microbiol. Immunol. 2011, 55, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Glatz, M.; Bosshard, P.; Schmid-Grendelmeier, P. The Role of Fungi in Atopic Dermatitis. Immunol. Allergy Clin. NA 2017, 37, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Scalabrin, D.M.F.; Bavbek, S.; Perzanowski, M.S.; Wilson, B.B.; Platts-Mills, T.A.E.; Wheatley, L.M. Use of Specific IgE in Assessing the Relevance of Fungal and Dust Mite Allergens to Atopic Dermatitis: A Comparison with Asthmatic and Nonasthmatic Control Subjects. J. Allergy Clin. Immunol. 1999, 104, 1273–1279. [Google Scholar] [CrossRef]
- Bayrou, O.; Pecquet, C.; Flahault, A.; Artigou, C.; Abuaf, N.; Leynadier, F. Head and Neck Atopic Dermatitis and Malassezia-Furfur-Specific IgE Antibodies. Dermatology 2005, 211, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Hiragun, M.; Hiragun, T.; Ishii, K.; Suzuki, H.; Tanaka, A.; Yanase, Y.; Mihara, S.; Haruta, Y.; Kohno, N.; Hide, M. Elevated Serum IgE against MGL_1304 in Patients with Atopic Dermatitis and Cholinergic Urticaria. Allergol. Int. 2014, 63, 83–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittermann, I.; Wikberg, G.; Johansson, C.; Lupinek, C.; Lundeberg, L.; Crameri, R.; Valenta, R.; Scheynius, A. IgE Sensitization Profiles Differ between Adult Patients with Severe and Moderate Atopic Dermatitis. PLoS ONE 2016, 11, e0156077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodská, P.; Panzner, P.; Pizinger, K.; Schmid-Grendelmeier, P. IgE-Mediated Sensitization to Malassezia in Atopic Dermatitis: More Common in Male Patients and in Head and Neck Type. Dermatitis 2014, 25, 120–126. [Google Scholar] [CrossRef]
- Lintu, P.; Savolainen, J.; Kortekangas-Savolainen, O.; Kalimo, K. Systemic Ketoconazole is an Effective Treatment of Atopic Dermatitis with IgE-Mediated Hypersensitivity to Yeasts. Allergy Eur. J. Allergy Clin. Immunol. 2001, 56, 512–517. [Google Scholar] [CrossRef]
- Yamasaki, S.; Matsumoto, M.; Takeuchi, O.; Matsuzawa, T.; Ishikawa, E.; Sakuma, M.; Tateno, H.; Uno, J.; Hirabayashi, J.; Mikami, Y.; et al. C-Type Lectin Mincle is an Activating Receptor for Pathogenic Fungus, Malassezia. Proc. Natl. Acad. Sci. USA 2009, 106, 1897–1902. [Google Scholar] [CrossRef] [Green Version]
- Sparber, F.; LeibundGut-Landmann, S. Host Responses to Malassezia spp. in the Mammalian Skin. Front. Immunol. 2017, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Johansson, H.J.; Vallhov, H.; Holm, T.; Gehrmann, U.; Andersson, A.; Johansson, C.; Blom, H.; Carroni, M.; Lehtiö, J.; Scheynius, A. Extracellular Nanovesicles Released from the Commensal Yeast Malassezia sympodialis are Enriched in Allergens and Interact with Cells in Human Skin. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Han, Y.; Sun, Y.; Jiang, H.; Liu, M.; Qi, R.; Gao, X. Extracellular Vesicles Derived from Malassezia Furfur Stimulate IL-6 Production in Keratinocytes as Demonstrated in In Vitro and In Vivo Models. J. Dermatol. Sci. 2019, 93, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Vallhov, H.; Johansson, C.; Veerman, R.E.; Scheynius, A. Extracellular Vesicles Released from the Skin Commensal Yeast Malassezia sympodialis Activate Human Primary Keratinocytes. Front. Cell. Infect. Microbiol. 2020, 10. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wang, S.; Zou, Z.; Tao, A.; Ai, Y. Activation Profile of THP-1 Derived Dendritic Cells Stimulated by Allergen Mal f 1 beyond its IgE-Binding Ability. Int. Immunopharmacol. 2018, 62, 139–146. [Google Scholar] [CrossRef]
- Ong, P.Y.; Ferdman, R.M.; Church, J.A. Association of Microbial IgE Sensitizations with Asthma in Young Children with Atopic Dermatitis. Ann. Allergy Asthma Immunol. 2012, 108, 212–213. [Google Scholar] [CrossRef]
- Van Woerden, H.C.; Gregory, C.; Brown, R.; Marchesi, J.R.; Hoogendoorn, B.; Matthews, I.P. Differences in Fungi Present in Induced Sputum Samples from Asthma Patients and Non-Atopic Controls: A Community Based Case Control Study. BMC Infect. Dis. 2013, 13, 1. [Google Scholar] [CrossRef] [Green Version]
- Al Bataineh, M.T.; Hamoudi, R.A.; Dash, N.R.; Ramakrishnan, R.K.; Almasalmeh, M.A.; Sharif, H.A.; Al-Hajjaj, M.S.; Hamid, Q. Altered Respiratory Microbiota Composition and Functionality Associated with Asthma Early in Life. BMC Infect. Dis. 2020, 20, 697. [Google Scholar] [CrossRef]
- Man, W.H.; Piters, W.A.A.D.S.; Bogaert, D. The Microbiota of the Respiratory Tract: Gatekeeper to Respiratory Health. Nat. Rev. Microbiol. 2017, 15, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Nagano, Y.; Elborn, S.; Millar, C.; Walker, J.M.; Goldsmith, C.E.; Rendall, J.; Moore, J.E. Comparison of Techniques to Examine the Diversity of Fungi in Adult Patients with Cystic Fibrosis. Med. Mycol. 2010, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Delhaes, L.; Monchy, S.; Fréalle, E.; Hubans, C.; Salleron, J.; Leroy, S.; Prevotat, A.; Wallet, F.; Wallaert, B.; Dei-Cas, E.; et al. The Airway Microbiota in Cystic Fibrosis: A Complex Fungal and Bacterial Community-Implications for Therapeutic Management. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Mounier, J.; Gouëllo, A.; Keravec, M.; Le Gal, S.; Pacini, G.; Debaets, S.; Nevez, G.; Rault, G.; Barbier, G.; Héry-Arnaud, G. Use of Denaturing High-Performance Liquid Chromatography (DHPLC) to Characterize the Bacterial and Fungal Airway Microbiota of Cystic Fibrosis Patients. J. Microbiol. 2014, 52, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Willger, S.D.; Grim, S.L.; Dolben, E.L.; Shipunova, A.; Hampton, T.H.; Morrison, H.G.; Filkins, L.M.; Toole, G.A.O.; Moulton, L.A.; Ashare, A.; et al. Characterization and Quantification of the Fungal Microbiome in Serial Samples from Individuals with Cystic Fibrosis. Microbiome 2014, 2, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Chotirmall, S.H.; O’Donoghue, E.; Bennett, K.; Gunaratnam, C.; O’Neill, S.J.; McElvaney, N.G. Sputum Candida Albicans Presages FEV 1 Decline and Hospital-Treated Exacerbations in Cystic Fibrosis. Chest 2010, 138, 1186–1195. [Google Scholar] [CrossRef]
- Masoud-Landgraf, L.; Badura, A.; Eber, E.; Feierl, G.; Marth, E.; Buzina, W. Modified Culture Method Detects a High Diversity of Fungal Species in Cystic Fibrosis Patients. Med. Mycol. 2014, 52, 179–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flemmer, A.W.; Yilmaz, E.; Mittal, R.; Müller-Edenborn, B.; Haas, A.; Holzinger, A.; Schulze, A. Malassezia Furfur Colonising the Respiratory Tract of Mechanically Ventilated Neonates. Z. Geburtshilfe Neonatol. 2008, 212, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Blaes, A.H.; Cavert, W.P.; Morrison, V.A. Malassezia: Is it a Pulmonary Pathogen in the Stem Cell Transplant Population? Transpl. Infect. Dis. 2009, 11, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Soret, P.; Vandenborght, L.E.; Francis, F.; Coron, N.; Enaud, R.; Avalos, M.; Schaeverbeke, T.; Berger, P.; Fayon, M.; Thiebaut, R.; et al. Respiratory Mycobiome and Suggestion of Inter-Kingdom Network during Acute Pulmonary Exacerbation in Cystic Fibrosis. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tragiannidis, A.; Bisping, G.; Koehler, G.; Groll, A.H. Minireview: Malassezia Infections in Immunocompromised Patients. Mycoses 2010, 53, 187–195. [Google Scholar] [CrossRef]
- Forrestel, A.K.; Kovarik, C.L.; Mosam, A.; Gupta, D.; Maurer, T.A.; Micheletti, R.G. Diffuse HIV-Associated Seborrheic Dermatitis-A Case Series. Int. J. STD AIDS 2016, 27, 1342–1345. [Google Scholar] [CrossRef] [PubMed]
- Pechère, M.; Remondat, C.; Bertrand, C.; Didierjean, L.; Saurat, J.H. A Simple Quantitative Culture of Malassezia spp. In HIV-Positive Persons. Dermatology 1995, 191, 348–349. [Google Scholar] [CrossRef]
- Moreno-Coutiño, G.; Sánchez-Cárdenas, C.D.; Bello-Hernández, Y.; Fernández-Martínez, R.; Arroyo-Escalante, S.; Arenas, R. Isolation of Malassezia Spp. in HIV-Positive Patients with and without Seborrheic Dermatitis. An. Bras. De Dermatol. 2019, 94. [Google Scholar] [CrossRef]
- Schechtman, R.; Hay, R.J.; Midgley, G. HIV Disease and Malassezia Yeasts: A Quantitative Study of Patients Presenting with Seborrhoeic Dermatitis. Br. J. Dermatol. 1995, 133, 694–698. [Google Scholar] [CrossRef]
- Krzyściak, P.; Bakuła, Z.; Gniadek, A.; Garlicki, A.; Tarnowski, M.; Wichowski, M.; Jagielski, T. Prevalence of Malassezia Species on the Skin of HIV-Seropositive Patients. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hamad, I.; Abdallah, R.A.; Ravaux, I.; Mokhtari, S.; Tissot-Dupont, H.; Michelle, C.; Stein, A.; Lagier, J.C.; Raoult, D.; Bittar, F. Metabarcoding Analysis of Eukaryotic Microbiota in the Gut of HIV-Infected Patients. PLoS ONE 2018, 13, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suhr, M.J.; Banjara, N.; Hallen-Adams, H.E. Sequence-Based Methods for Detecting and Evaluating the Human Gut Mycobiome. Lett. Appl. Microbiol. 2015, 62, 209–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liguori, G.; Lamas, B.; Richard, M.L.; Brandi, G.; da Costa, G.; Hoffmann, T.W.; Di Simone, M.P.; Calabrese, C.; Poggioli, G.; Langella, P.; et al. Fungal Dysbiosis in Mucosa-Associated Microbiota of Crohn’s Disease Patients. J. Crohn’s Colitis 2016, 10, 296–305. [Google Scholar] [CrossRef]
- Frykman, P.K.; Nordenskjöld, A.; Kawaguchi, A.; Hui, T.T.; Granström, L.; Cheng, Z.; Tang, J.; Underhill, D.M.; Iliev, I.; Funari, V.A. Characterization of Bacterial and Fungal Microbiome in Children with Hirschsprung Disease with and without a History of Enterocolitis: A Multicenter Study. PLoS ONE 2015, 10, e0124172. [Google Scholar] [CrossRef] [Green Version]
- Quinton, J.F.; Sendid, B.; Reumaux, D.; Duthilleul, P.; Cortot, A.; Grandbastien, B.; Charrier, G.; Targan, S.R.; Colombel, J.F.; Poulain, D. Anti-Saccharomyces cerevisiae Manna Antibodies Combined with Antineutrophil Cytoplasmic Autoantibodies in Inflammatory Bowel Disease: Prevalence and Diagnostic Role. Gut 1998, 42, 788–791. [Google Scholar] [CrossRef] [Green Version]
- Rivas, M.A.; Beaudoin, M.; Gardet, A.; Stevens, C.; Zhang, C.K.; Boucher, G.; Ripke, S.; Ellinghaus, D.; Burtt, N.; Fennell, T.; et al. Deep Resequencing of GWAS Loci Identifies Independent Rare Variants Associated with Inflammatory Bowel Disease. Nat. Genet. 2011, 43, 1066–1073. [Google Scholar] [CrossRef] [Green Version]
- Sokol, H.; Leducq, V.; Aschard, H.; Pham, H.-P.; Jegou, S.; Landman, C.; Cohen, D.; Liguori, G.; Bourrier, A.; Nion-Larmurier, I.; et al. Fungal Microbiota Dysbiosis in IBD. Gut 2017, 66, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Jawhara, S.; Thuru, X.; Standaert-Vitse, A.; Jouault, T.; Mordon, S.; Sendid, B.; Desreumaux, P.; Poulain, D. Colonization of Mice by Candida albicans Is Promoted by Chemically Induced Colitis and Augments Inflammatory Responses through Galectin-3. J. Infect. Dis. 2008, 197, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Chiaro, T.R.; Soto, R.; Stephens, W.Z.; Kubinak, J.L.; Petersen, C.; Gogokhia, L.; Bell, R.; Delgado, J.C.; Cox, J.; Voth, W.; et al. A Member of the Gut Mycobiota Modulates Host Purine Metabolism Exacerbating Colitis in Mice. Sci. Transl. Med. 2017, 9. [Google Scholar] [CrossRef]
- Limon, J.J.; Tang, J.; Li, D.; Targan, S.R.; Mcgovern, D.P.B.; Underhill, D.M.; Limon, J.J.; Tang, J.; Li, D.; Wolf, A.J.; et al. Malassezia Is Associated with Crohn’s Disease and Exacerbates Colitis in Mouse Models. Cell Host Microbe 2019, 25, 377–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagnière, J.; Raisch, J.; Veziant, J.; Barnich, N.; Bonnet, R.; Buc, E.; Bringer, M.A.; Pezet, D.; Bonnet, M. Gut Microbiota Imbalance and Colorectal Cancer. World J. Gastroenterol. 2016, 22, 501–518. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Kong, C.; Li, H.; Huang, L.; Qu, X.; Qin, N.; Qin, H. Dysbiosis Signature of Mycobiota in Colon Polyp and Colorectal Cancer. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 2457–2468. [Google Scholar] [CrossRef]
- Coker, O.O.; Nakatsu, G.; Dai, R.Z.; Wu, W.K.K.; Wong, S.H.; Ng, S.C.; Chan, F.K.L.J.J.Y.S.; Yu, J. Enteric Fungal Microbiota Dysbiosis and Ecological Alterations in Colorectal Cancer. Gut 2018, 68. [Google Scholar] [CrossRef] [PubMed]
- Aykut, B.; Pushalkar, S.; Chen, R.; Li, Q.; Abengozar, R.; Kim, J.I.; Shadaloey, S.A.; Wu, D.; Preiss, P.; Verma, N.; et al. The Fungal Mycobiome Promotes Pancreatic Oncogenesis via Activation of MBL. Nature 2019, 574, 264–267. [Google Scholar] [CrossRef]
- Itzhaki, R.F.; Wozniak, M.A.; Appelt, D.M.; Balin, B.J. Infiltration of the Brain by Pathogens Causes Alzheimer’s Disease. Neurobiol. Aging 2004, 25, 619–627. [Google Scholar] [CrossRef]
- Miklossy, J.; Kis, A.; Radenovic, A.; Miller, L.; Forro, L.; Martins, R.; Reiss, K.; Darbinian, N.; Darekar, P.; Mihaly, L.; et al. Beta-amyloid Deposition and Alzheimer’s Type Changes Induced by Borrelia spirochetes. Neurobiol. Aging 2006, 27, 228–236. [Google Scholar] [CrossRef] [Green Version]
- De Chiara, G.; Marcocci, M.E.; Sgarbanti, R.; Civitelli, L.; Ripoli, C.; Piacentini, R.; Garaci, E.; Grassi, C.; Palamara, A.T. Infectious Agents and Neurodegeneration. Mol. Neurobiol. 2012, 614–638. [Google Scholar] [CrossRef] [Green Version]
- Alonso, R.; Pisa, D.; Aguado, B.; Carrasco, L. Identification of Fungal Species in Brain Tissue from Alzheimer ’s Disease by Next-Generation Sequencing. J. Alzheimer. Dis. 2017, 58, 55–67. [Google Scholar] [CrossRef]
- Alonso, R.; Pisa, D.; Fernández-Fernández, A.M.; Carrasco, L. Infection of Fungi and Bacteria in Brain Tissue from Elderly Persons and Patients with Alzheimer’s Disease. Front. Aging Neurosci. 2018, 10, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Alonso, R.; Pisa, D.; Fernández-Fernández, A.M.; Rábano, A.; Carrasco, L. Fungal Infection in Neural Tissue of Patients with Amyotrophic Lateral Sclerosis. Neurobiol. Dis. 2017, 108, 249–260. [Google Scholar] [CrossRef]
- Alonso, R.; Fernández-Fernández, A.M.; Pisa, D.; Carrasco, L. Multiple Sclerosis and Mixed Microbial Infections. Direct Identification of Fungi and Bacteria in Nervous Tissue. Neurobiol. Dis. 2018, 117, 42–61. [Google Scholar] [CrossRef]
- Drummond, R.A.; Collar, A.L.; Swamydas, M.; Rodriguez, A.; Lim, J.K.; Mendez, L.M.; Fink, D.L.; Hsu, A.P.; Zhai, B.; Karauzum, H.; et al. CARD9-Dependent Neutrophil Recruitment Protects against Fungal Invasion of the Central Nervous System. PLoS Pathog. 2015, 11, e1005293. [Google Scholar] [CrossRef]
- Drummond, R.A.; Swamydas, M.; Oikonomou, V.; Zhai, B.; Dambuza, I.M.; Schaefer, B.C.; Bohrer, A.C.; Mayer-Barber, K.D.; Lira, S.A.; Iwakura, Y.; et al. CARD9+ Microglia Promote Antifungal Immunity via IL-1β- and CXCL1-Mediated Neutrophil Recruitment. Nat. Immunol. 2019, 20, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Bixler, S.L.; Mattapallil, J.J. Loss and Dysregulation of Th17 Cells during HIV Infection. Clin. Dev. Immunol. 2013, 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamad, I.; Ranque, S.; Azhar, E.I.; Yasir, M. Culturomics and Amplicon-based Metagenomic Approaches for the Study of Fungal Population in Human Gut Microbiota. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef]
- Mbareche, H.; Veillette, M.; Bilodeau, G.; Duchaine, C. Comparison of the Performance of ITS1 and ITS2 as Barcodes in Amplicon-Based Sequencing of Bioaerosols. PeerJ 2020, 2020, 1–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Psoriasis Patients (PP) and/or Controls | Samples | Methods | Malassezia | References |
|---|---|---|---|---|
| 22 PP vs. 30 healthy subjects (HS) | lesional and non-lesional skin | Nested PCR | High detection (64–96%) in PP Species diversity (3.7 ± 1.6 in PP vs. 2.8 ± 0.8 in HS) | [15] |
| 50 PP vs. 50 HS | lesional and non-lesional skin | Culture/ITS2 PCR-RFLP and D1/D2 sequencing | 68% of PP cases vs. 60% of HS In PP, more colonies isolated in psoriatic lesions (53 colonies) than in non-lesional skin (19), especially on the scalp (p = 0.03). High species richness in PP than HS | [16] |
| 34 PP vs. 25 HS | skin lesion and non-lesion | ITS1 metabarcoding | A significantly higher abundance of M. restricta and M. sympodialis in psoriatic lesions than HS | [18] |
| Patients | Sex | Age (Years) | Samples | Culture | Malassezia PCR |
|---|---|---|---|---|---|
| Patient 1 | Female | 77 | Sinus | Negative | Positive |
| Patient 2 | Female | 27 | Sinus | Negative | Positive |
| Patient 3 | Male | 52 | Sinus | Negative | Positive |
| Patient 4 | Male | 56 | Sinus | Negative | Positive |
| Patient 5 | Female | 44 | Sinus | Negative | Positive |
| Patient 6 | Female | 53 | Sinus | Negative | Positive |
| Patient 7 | Female | 13 | Sinus 1 | Negative | Positive |
| Sinus 2 | Negative | Positive | |||
| Patient 8 | Male | 48 | Sinus | Negative | Negative |
| Patient 9 | Male | 25 | Sinus 1 | Negative | Positive |
| Sinus 2 | Negative | Positive | |||
| Patient 10 | Female | 53 | Sinus | Negative | NA |
| Patient 11 | Female | 72 | Sinus 1 | Negative | NA |
| Sinus 2 | Negative | NA |
| Patients and/or Controls | Samples | Methods | Malassezia | References |
|---|---|---|---|---|
| 23 CRS vs. 11 controls | Sinonasal swabs | 18S pyrosequencing | 100% of all sinus samples Relative abundance of 50.09% in CRS patients vs. 57.5% in 11 controls | [27] |
| 21 CRS vs. seven controls | Sinus brushings | qPCR | 68% of all samples with no prevalence variation among the groups (p > 0.99) M. restricta (46%) than M. globosa (14%, p = 0.029) | [28] |
| 106 CRS vs. 38 controls | Mucosal swabs | ITS2 metabarcoding | 100% of subjects with an abundance of 86% of the sequences | [12] |
| 56 AD vs. 32 controls | lesional and non-lesional skin | qPCR | High Malassezia colonization in patients with severe AD, ≈ two- to fivefold that in mild and moderate AD patients and healthy subjects (p < 0.05) | [37] |
| 106 head and neck AD (HNAD) vs. 61 controls | Blood | Anti-Malassezia IgE assays (Pharmacia CAP System) | Significant correlation between M. furfur IgE levels and severity in HNAD patients (p < 0.0001) | [42] |
| 63 AD vs. 23 controls | Blood | Anti-M. globosa IgE by Enzyme-linked immunosorbent assay (ELISA) | High significantly IgE and correlated with severity in AD patients compared to normal controls (p < 0.001) | [43] |
| 53 severe AD and 126 AD moderate vs. 140 controls | Blood | Anti-M. sympodialis IgE (ImmunoCAP™) | 62% of severe AD compared to 39% of moderate AD (p < 0.01), No positive controls | [44] |
| 74 HNAD vs. 99 Non-HNAD | Blood | Anti-Malassezia IgE (ImmunoCAP™) | Significantly higher levels of IgE in HNAD patients than non-HAND (p < 0.001) | [45] |
| 73 AD, 156 asthmatic and 212 control patients | Blood | Anti-M. furfur IgE Fluoroimmuno assay (CAP system) | AD (53%), asthmatic (1%) and non-asthmatic control subjects (0.5%) | [41] |
| 30 asthma patients vs. 13 controls | Sputum | 18S pyrosequencing | Only in asthma patients with a percentage of reads from 0.012% to 21.651% | [54] |
| 21 asthma patients vs. 19 controls | Sputum | ITS2 metabarcoding | Mean abundance of 67.69%, 27.04%, 2.02% and 2.94% in pediatric asthma, adult asthma, healthy adult, and healthy pediatric participants, respectively. Significant abundance of Malassezia in the airways of asthmatic patients receiving steroid therapy combined with leukotriene receptor antagonists. | [55] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdillah, A.; Ranque, S. Chronic Diseases Associated with Malassezia Yeast. J. Fungi 2021, 7, 855. https://doi.org/10.3390/jof7100855
Abdillah A, Ranque S. Chronic Diseases Associated with Malassezia Yeast. Journal of Fungi. 2021; 7(10):855. https://doi.org/10.3390/jof7100855
Chicago/Turabian StyleAbdillah, Abdourahim, and Stéphane Ranque. 2021. "Chronic Diseases Associated with Malassezia Yeast" Journal of Fungi 7, no. 10: 855. https://doi.org/10.3390/jof7100855
APA StyleAbdillah, A., & Ranque, S. (2021). Chronic Diseases Associated with Malassezia Yeast. Journal of Fungi, 7(10), 855. https://doi.org/10.3390/jof7100855