3.5. Taxonomy
Based on the results of multilocus phylogenetic analysis, ancestral inference, barcode gap, morphological data, and assessment of biogeography, the new taxonomic treatment of
Codinaea and similar fungi is presented. The current circumscription of
Codinaea is revised and several new genera, species, and combinations are proposed below. A key to
Codinaea, its segregates, and similar fungi is provided (
Table 2).
3.5.1. The Genus Codinaea
Codinaea Maire, Publ. Inst. Bot. Barcelona 3: 15. 1937.
= Menisporella Agnihothr., Proc. Natl. Acad. Sci. India B 56: 98. 1962.
= Codinaeopsis Morgan-Jones, Mycotaxon 4: 166. 1976.
= Bahusutrabeeja Subram. & Bhat, Can. J. Bot. 55: 204. 1977.
= Phialolunulospora Z.F. Yu & R.F. Castañeda, MycoKeys 76: 23. 2020.
Type species. Codinaea aristata Maire, Publ. Inst. Bot. Barcelona 3: 15. 1937.
Emended description: Colonies on natural substrate effuse, hairy, brown to black, composed of setae and conidiophores, mycelium semi-immersed or immersed. Anamorph. Setae present or absent, if present single or arise in groups from stromatic cells or knots of hyphal cells, erect, straight or flexuous, septate, pigmented, thick-walled, paler and thinner-walled toward the apex, unbranched, apex sterile, tapering or modified into a phialide with a terminal or several lateral openings. Conidiophores macronematous, mononematous, single or arise in fascicles from stromatic cells or knots of hyphal cells around the base of the setae, erect, straight or flexuous to slightly geniculate or undulate, unbranched or branched, occasionally with nodose or collar-like hyphae formed just below the septa, septate, smooth, pigmented, paler toward the apex, terminating into a phialide or with a sterile setiform extension. Conidiogenous cells integrated, terminal or discrete, lateral, mono- or polyphialidic, extending percurrently and sympodially, paler than the conidiophore; collarettes funnel-shaped. Conidia of two morphologically distinct types; macroconidia predominantly falcate to lunate, sometimes oblong-falcate, ellipsoid to ellipsoid-fusiform or broadly oblong, occasionally vermiform, slightly truncate at the base, with an inconspicuous basal scar, with a straight or gently curved setula at each end, sometimes setulae also inserted ventrally and dorsally, or also globose to pyriform with setulae distributed over the surface of the conidium, aseptate, hyaline, conidia accumulate in slimy fascicles; microconidia (formed occasionally and only in vitro) ellipsoidal to oblong, aseptate, hyaline, with a miniature setula. Teleomorph. Unknown.
Habitat and geographical distribution: Codinaea includes mainly saprobes that have been recorded from soil, decaying fruits, leaves, petioles, palm fronds, bark, wood, and roots, as well as leaf spots of a wide range of plant hosts. Some species have also been isolated as endophytes. In addition, C. fertilis has been recorded as a plant pathogen. The genus Codinaea has a wide geographic distribution with occurrence in the tropical and temperate climatic zones supported by literature, field records, and environmental sequence data from the GlobalFungi database.
Note: In the three-gene phylogeny (
Figure 2),
Codinaea is shown as a strongly supported monophyletic clade. Based on the results of the phylogenetic analysis, the generic concept of
Codinaea was emended and 14 species were accepted in the genus; the inclusion of 13 species was verified with DNA data. The teleomorph of
Codinaea is unknown. Although we were not successful in recollecting
C. aristata, we analyzed seven other species that match the
Codinaea s. str. Morphotype in all details, i.e., dark brown, thick-walled central seta or setae arranged in a fascicle with shorter, paler, and thinner-walled conidiophores bearing terminal phialides and falcate, aseptate, hyaline conidia with setulae at each end. All seven species of the
C. aristata morphotype clustered in the
Codinaea clade, namely
C. assamica, C. fertilis,
C. pandanicola,
C. paniculata,
C. phasma,
C. siamensis, and
C. terminalis.
Based on the arrangement of setae and conidiophores and their branching pattern, we distinguish four morphotypes C1–C4 (
Figure 8) in
Codinaea. In addition to species with the
Codinaea s. str. morphotype (C1), other lineages of species characterized by three other morphotypes were nested among them. Four species with unbranched, dark brown, thick-walled conidiophores that closely resemble the setae and terminate into a monophialide (C2: e.g.,
C. ellipsoidea and
C. lignicola); one species with conidiophores with a sterile setiform extension and integrated, terminal phialides borne in groups on short unilateral, branched stalks (C3:
C. amazonensis), and one species with branched conidiophores and discrete, lateral phialides borne on nodose hyphae or directly on conidiophores (C4:
C. gonytrichodes). The conidiophore variability found in
Codinaea compares well with that of
Chloridium (
Chl.) and
Menispora (
Figure 8 and
Figure 9) (see Discussion,
Section 4.1).
Conidia also show some variability. Although they are always aseptate and hyaline, they vary in shape. They are mostly falcate to lunate, curved, occasionally vermiform but several species have also ellipsoidal-fusiform to more or less ellipsoidal conidia with setulae inserted at the apical and basal ends. The globose to pyriform conidia of C. dwaya with numerous setulae irregularly distributed over the surface represent a character that is unusual in Codinaea. The falcate conidia of C. phasma form a simple setula at each end under natural conditions, however, in culture, they become irregularly ellipsoidal and have 3(–5) setulae inserted also on the ventral and dorsal sides.
Interestingly, the ex-type strains of
Bahusutrabeeja dwaya CBS 261.77 [
120] and
Phialolunulospora vermispora CGMCC 3.19632 [
50], and a non-type strain of
Codinaeopsis gonytrichodes CBS 593.93 [
42] clustered in this clade. The two former species form simple conidiophores terminating into a monophialide and correspond to the C2 morphotype, while
C. gonytrichodes is the only representative of the C4 morphotype. Based on the phylogenetic evidence,
Bahusutrabeeja,
Codinaeopsis, and
Phialolunulospora are proposed as generic synonyms of
Codinaea. Although
Codinaeopsis and
Phialolunulospora are monotypic genera,
Bahusutrabeeja accommodates five other species, but only two of them resemble
C. dwaya.
Several other morphologically similar species, whose living cultures or DNA sequence data are not available for study, are discussed below. These taxa should be considered candidates for inclusion in
Codinaea, however, such relationships need to be supported by molecular data. Species such as
Dictyochaeta gyrosetula [
24],
D. intermedia [
121],
D. plovercovensis [
122],
D. tilikfrei [
123], and
D. vittata [
24] correspond to the
Codinaea s. str. morphotype (C1). The C2 morphotype also occurs in
C. aquatica [
124],
C. leomaiae [
125],
C. tropicalis [
126],
D. fimbriaspora [
44],
D. multifimbriata [
34],
D. multisetula, and
D. renispora [
44], which often have ellipsoidal to reniform or drop-shaped conidia with two or more setulae. Morphotype C4 with discrete phialides borne on nodose hyphae was also recorded in
D. pahangensis [
21] and
Dictyochaetopsis polysetosa [
127]. A synopsis table with diagnostic features of accepted species of
Codinaea is provided in the
Supplementary File: Table S4.
Codinaea amazonensis (Matsushima) Réblová & Hern.-Restr.,
comb. nov. MycoBank MB 842191 (
Figure 10 and
Figure 11).
Basionym. Menispora amazonensis Matsush., Matsush. Mycol. Mem. 7: 57. 1993.
Culture characteristics: On CMD: colonies 33–35 mm diam, circular, flat, margin entire, cobwebby, beige-brown, beige toward the margin, reverse isabelline. On MLA: colonies 65–67 mm diam, circular, raised, margin entire, velvety-lanose, floccose, furrowed, zonate, smoke grey centrally, dark grey toward the margin, white-grey at the margin, with a grey-brown outer zone of submerged growth, reverse dark grey-brown. On OA: colonies 57–58 mm diam after 28 d, circular, raised, margin entire, velvety to cobwebby, mucoid at the margin, furrowed, zonate, with an intermediate zone of sparse growth, beige-brown, olivaceous brown at the margin, reverse dark brown. On PCA: colonies 58–60 mm diam, circular, slightly raised, flat at the margin, margin entire to curled, cobwebby, floccose locally, mucoid toward the periphery, beige-brown, dark brown with an orange tinge at the margin, reverse dark brown. Sporulation was sparse on CMD, OA, and PCA, moderate on MLA.
Colonies on MLA effuse, hairy, mycelium composed of branched, septate, hyaline to subhyaline hyphae 1.5–2 μm diam. Anamorph. Setae absent. Conidiophores 260–320 μm long, 4.5–6.5 μm wide near the base, macronematous, single, erect, septate, smooth, dark brown, slightly paler toward the apex, with a sterile setiform extension, apex acute, pale brown, conidiophores with unilateral phialide-bearing stalks, which arise just below the septa, 19–36 μm long including the phialides, 3.5–5 μm wide at the base, stalks are curved upwards, composed of a cylindrical basal cell and several compact, densely aggregated cells bearing 1–15 phialides, basal cell is medium to dark brown, other cells are pale brown to subhyaline. Conidiogenous cells 8.5–13.5 × 3–4.5(–5) μm tapering to 1–2 μm below the collarette, discrete, lateral, arise on stalks or on conidiophores, sometimes stalked or sessile phialides arise on vegetative mycelium, mostly monophialidic, occasionally with 1–2 lateral openings, extending percurrently and sympodially, lageniform, pale brown, paler than the conidiophore; collarettes 2.5–4 μm wide, funnel-shaped, subhyaline. Conidia 9–11.5 × 1.5–2(–2.5) μm (mean ± SD = 9.9 ± 0.7 × 2.0 ± 0.2 μm), falcate, tapering toward both ends, slightly truncate at the base with an inconspicuous scar, aseptate, hyaline, with straight or gently curved setula at each end 5.5–9 μm long, inserted terminally at the apex, subterminally at the base, conidia accumulate in pale ochre fascicles. Teleomorph. Unknown.
Specimen examined: BRAZIL, Mata Avenca-Santa Rita, on rotten leaf, 10 September 1997, J. Guarro (culture MUCL 41171).
Habitat and geographical distribution: According to literature, it is a rare species isolated from soil and decaying leaves of
Quercus dentata and other unidentified hosts in South America and Asia: Brazil and Japan [
5,
128,
129]. Czeczuga and Orłowska [
130] reported this species (as
Menispora amazonensis) in the water from melting ice collected from branches of coniferous trees in Poland. However, given the similarity of
C. amazonensis to
Menispora tortuosa [
131,
132,
133], which is common in the temperate zone of Europe and North America, and the known distribution of
C. amazonensis in tropical and subtropical areas, it is possible that the fungus from melting snow may have been misidentified.
According to GlobalFungi, the identical sequences were found in one soil sample from the forest in South America (Amazonian river basin, Brazil) [
117], which is in agreement with the original data and where the type was collected [
128]. The location has a tropical climate (MAT avg. 25 °C, MAP avg. 2803 mm).
Notes:
Codinaea amazonensis was originally described from a decaying leaf in the Amazon river basin in Peru [
128]. However, no type or authentic material is available. Our strain MUCL 41171, originally published under the name
Codinaea gonytrichodes [
129], matches the protologue and original illustration of
C. amazonensis in all details.
This species differs from other
Codinaea species in the branching pattern of the conidiophores, which have a sterile extension. In addition, short stalks arise unilaterally almost the entire length of the conidiophores; they consist of several compact cells that bear a group of phialides.
Codinaea amazonensis superficially resembles
C. gonytrichodes, which forms collar-like and nodose hyphae at the conidiophore septa, from which arise discrete phialides and occasionally branches and setae. Interestingly, the fungus reported as
C. gonytrichodes from Japan and described and illustrated by Matsushima [
5], is in fact
C. amazonensis [
128].
Codinaea aristata Maire, Publ. Inst. Bot. Barcelona 3: 15. 1937. (
Figure 1).
≡ Dictyochaeta aristata (Maire) Aramb. & Cabello, Mycotaxon 34: 681. 1989. (Nom. inval., Art. 41.4).
≡ Dictyochaeta aristata (Maire) Whitton, McKenzie & K.D. Hyde, Fungal Divers. 4: 136. 2000.
Typification: SPAIN, Catalonia, Sant Miquel de Cladells, on decaying stem of
Rubus sp. lying on the ground, date unknown, R. Maire (
holotype not available). Lectotype illustration: Maire, Fungi Catalaunici: Series altera. Contributions a l’étude de la flore mycologique de la Catalogne. Publ. Inst. Bot. Barcelona 3: 15,
Figure 1. 1937 (
lectotype illustration designated here MBT 10004618,
Figure 1).
Habitat and geographical distribution: Saprobe on a stem of Rubus sp. in Europe, Spain.
Notes: Maire [
1] described
C. aristata on a decaying stem of
Rubus sp. lying on the ground close to the torrent near Sant Miquel de Cladells in Catalonia, Spain. The species was characterized with setae 250 μm long and longer, sterile, arising singly or in a group of two, erect, dark brown, thick-walled, several-septate, smooth, unbranched, sterile at the apex; conidiophores up to 100 μm long, in bundles with setae, pale brown, thin-walled, flexuous, 1–2-septate, tapering apically; conidia 12–14 × 2 μm, falcate, aseptate, hyaline, with a setula at each end, 4–6 μm long at the apical end and ca. 1 μm long at the basal end, soon evanescent.
Because the holotype or other authentic material of
C. aristata could not be found [
2] and other collections or cultures of this species are not available, the illustration accompanying the protologue [
1] is designated here as a lectotype (
Figure 1).
Codinaea aristata resembles
C. fertilis [
2] and
C. siamensis [
47], but differs in conidia with shorter setulae of a different length.
Codinaea assamica (Agnihothr.) S. Hughes & W.B. Kendr., N. Z. J. Bot. 6: 334. 1968. (
Figure 12 and
Figure 13).
Basionym. Menisporella assamica Agnihothr., Proc. Indian natn Sci. Acad., Part B. Biol. Sci. 56: 99. 1962.
≡ Dictyochaeta assamica (Agnihothr.) Aramb., Cabello & Mengasc., Darwiniana 28: 297. 1988. (Nom. inval., Art. 41.4).
= Codinaea acaciae Crous & M.J. Wingf., Persoonia 34: 181. 2015.
Culture characteristics: On CMD: colonies 75–78 mm diam, circular, flat, margin fimbriate, sparsely lanose to cobwebby, funiculose at the inoculation block, olivaceous beige, grey-brown centrally, aerial mycelium with numerous colorless exudates, reverse olivaceous beige. On MLA: colonies 48–50 mm diam, circular, raised, margin fimbriate, lanose, grey-brown, olivaceous brown at the margin, reverse dark brown. On OA: colonies 69–70 mm diam, raised, margin fimbriate, lanose, floccose, grey with irregular white patches, dark olivaceous grey at the margin, reverse dark grey. On PCA: colonies 70–72 mm diam, circular, flat, margin fimbriate, cobwebby, lanose at the inoculation block, zonate, pale brown at the centre becoming olivaceous beige toward the periphery, dark olivaceous brown at the margin, reverse of the same colors. Sporulation was abundant on MLA, moderate on PCA and OA, absent on CMD.
Colonies on SNA with pine needles effuse, hairy, brown, mycelium composed of branched, septate, subhyaline to pale brown hyphae 1.5–2.5 μm diam. Anamorph. Setae 273–360 μm long, 5–7 μm wide near the swollen base, arise solitary or in groups of two from conspicuous dark brown stromatic cells, erect, straight or flexuous, septate, smooth, unbranched, dark brown, thick-walled, paler and thinner-walled toward the apex, apex pale brown to subhyaline, sterile when young, broadly rounded, later with a terminal or 1–3 lateral phialidic openings. Conidiophores 42–125 μm long, 3.5–4.5(–5) μm wide near the base, macronematous, arise in fascicles of 3–9 from stromatic cells around the base of the setae, unbranched, erect, straight or flexuous or geniculate to undulate, septate, smooth, pale to medium brown, gradually paler toward the apex. Conidiogenous cells 14.5–27 × 3.5–4.5(–5) μm tapering to 1.5–2 μm below collarette, integrated, terminal, mono- or polyphialidic with 1–3 lateral openings while internally septa can be formed, extending percurrently and sympodially, cylindrical, slightly swollen bellow the collarette, pale brown, subhyaline at the apex; collarettes 3.5–4.5 μm wide, 2–2.5 μm deep, funnel-shaped, subhyaline to pale brown. Conidia 14–18 × 2.5–3.5 μm (mean ± SD = 15.9 ± 0.9 × 2.9 ± 0.3 μm), falcate, tapering toward both ends, narrowly rounded apically, slightly truncate at the base with an inconspicuous scar, aseptate, hyaline, with straight or gently curved setula at each end (5.5–)7–13.5 μm long, inserted terminally at the apex, subterminally at the base, conidia accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Specimen examined: MALAYSIA, Sarawak, from leaf spots of Acacia mangium, May 2014, M.J. Wingfield (ex-type strain of Codinaea acaciae CBS 139907).
Habitat and geographical distribution: Saprobe on woody roots, leaf litter, leaf spots and decaying petioles of
Acacia mangium,
Camelia sinensis,
Cedrela odorata,
Cyperus radians, and other unidentified hosts in Central and South America and Asia: Brazil, Cuba, India, and Malaysia [
3,
25,
36,
53,
134,
135]. Rambelli et al. [
35] reported
C. assamica on fallen leaves of 10 hosts from Africa from Ivory Coast, namely
Anthonotha fragrans,
Calpocalyx brevibracteatus,
Chrysophyllum taiense,
Dialium aubrevillei,
Hypselodelphys violacea,
Lophira alata,
Manniophyton fulvum,
Memecylon lateriflorum,
Pentaclethra macrophylla, and
Piptadeniastrum africanum.
According to GlobalFungi, the identical sequences were found in two soil samples from the forest in Asia on Malaysian peninsula and Kalimantan [
117], and in two root samples from forest in South America in French Guiana [
136]. The sites have a tropical climate (MAT avg. 24 °C, MAP avg. 2612 mm).
Notes: Hughes and Kendrick [
2] gave a detailed description of
C. assamica by examining the holotype material of
Menisporella assamica from India [
3]. Unfortunately, no culture derived from the type or other material of
C. assamica is available. Our study of the ex-type strain of
C. acaciae CBS 139907 revealed it is remarkably similar to
C. assamica. Crous et al. [
53] isolated
C. acaciae from leaf spots of
Acacia mangium in Malaysia and based the description on observations in culture. However, the setae were absent and the conidiophores were somewhat smaller, 15–30 × 3–3.5 μm. Based on our experience, some
Codinaea species do not form setae readily on agar media and conidiophores may not be fully developed, which can complicate a correct identification. However, on SNA with pine needles, the fungus formed setae arranged in typical bundles with conidiophores growing from stromatic basal cells. Although conidia have one setula at each end, we have rarely observed conidia in culture with two setulae inserted at one end (two conidia in total), a character observed for example in
C. phasma (this study). Based on a detailed comparison of our observations of
C. acaciae with
C. assamica on natural material [
2,
3] and in culture [
25], we concluded that they are conspecific. Therefore,
C. acaciae is reduced to synonymy with
C. assamica.
Codinaea assamica resembles
C. paniculata [
46],
C. siamensis [
47], and
C. terminalis [
30].
Codinaea paniculata mostly occurs in the Holarctic zone and differs in shorter setae, monophialidic conidiogenous cells and conidia with shorter setulae.
Codinaea siamensis possesses broader conidia, and
C. terminalis differs in monophialidic conidiogenous cells and larger conidia.
Dictyochaeta plovercovensis [
122], whose culture and DNA data are not available, also resembles
C. assamica but differs in shorter conidiophores and narrower conidia with shorter setulae.
Codinaea dwaya (Subram. & Bhat) Réblová & Hern.-Restr.,
comb. nov. MycoBank MB 842192. (
Figure 13 and
Figure 14).
Basionym. Bahusutrabeeja dwaya Subram. & Bhat, Can. J. Bot. 55: 2204. 1977.
Culture characteristics: On CMD: colonies 75–76 mm diam, circular, flat, margin entire, mucoid, smooth, funiculose at the inoculation block, isabelline, reverse of the same color. On MLA: colonies 59–61 mm diam, circular, raised to slightly convex, margin weakly fimbriate, lanose, floccose, mucoid at the margin, white-grey with an olivaceous brown outer zone, reverse dark olivaceous brown. On OA: colonies 75–78 mm diam, circular, flat to slightly raised, margin fimbriate, lanose becoming mucoid, whitish-beige, pale brown becoming cinnamon-brown centrally when mucoid, reverse dark grey. On PCA: colonies 82–83 mm diam, circular, flat, margin fimbriate, mucoid centrally, cobwebby toward the periphery, funiculose at the inoculation block, pale olivaceous-isabelline centrally, white grey at the margin, reverse whitish. Sporulation was absent on all media.
Colonies on CMA with U. dioica stems effuse, hairy, mycelium composed of branched, septate, hyaline to brown hyphae 1–2.5 μm diam. Anamorph. Setae absent. Conidiophores (33–)74–254 μm long, tapering toward the base 3–5.5(–8.6) μm wide, gradually widening upwards, 5–10(–11.5) μm wide at the broadest point, macronematous, solitary, unbranched, straight or flexuous, smooth, medium brown becoming dark brown at maturity, darkest in the middle, paler toward the apex and base. Conidiogenous cells 18–66 × 6.5–11.5 μm, tapering to 3–6.5(–7.5) μm below the collarette, integrated, terminal, monophialidic, extending percurrently, subcylindrical with a swollen venter, fulvous, gradually paler toward the apex; collarette 4.5–10 μm wide, 1.5–3 μm deep, pale brown, flared, shallow, indistinct. Conidia of two morphologically distinct types: macroconidia variable in shape and size, the first conidium is broadly oblong or pyriform 15.5–22 × 10.5–14.5 μm (mean ± SD = 18.7 ± 2.1 × 13 ± 2.8 μm), subsequent conidia globose 15–16 μm diam (mean ± SD = 15.5 ± 0.5 μm), thick-walled, aseptate, hyaline, with 6–10 straight or gently curved setulae distributed over the surface, 6–10.5 μm long; microconidia 4.5–6.5 × 2–2.5 μm (mean ± SD = 5.1 ± 0.8 × 2.4 ± 0.2 μm), formed from the same conidiogenous loci, ellipsoidal to oblong, aseptate, hyaline, with a miniature setula at one end 1–1.5 μm long; macroconidia and microconidia accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Specimen examined: INDIA, Karnataka, Coorg district, near Abby Falls, on dead twigs of Coffea arabica, 10 May 1976, D.J. Bhat (ex-type culture CBS 261.77 = JCM 6357 = IMI 213921).
Habitat and geographical distribution: Saprobe on decaying bark and wood of
Coffea arabica and other unknown hosts, known from Asia from China and India [
120,
135,
137,
138]. The GlobalFungi database did not contain similar sequences (≥98% sequence identity) of this species.
Notes: In the phylogenetic analysis, the ex-type strain of
B. dwaya, the generic type, clustered in the
Codinaea clade among species with falcate to ellipsoidal conidia with two or more setulae, setae and variability in conidiophores. Based on this result,
Bahusutrabeeja is reduced to synonymy with
Codinaea and a new combination is proposed. The unique characters of
C. dwaya are globose or pyriform conidia with multiple appendages. Our observations of the ex-type strain of
C. dwaya are consistent with those of Subramanian and Bhat [
120]. In addition, we observed microconidia with a rudimentary appendage, which formed alongside the macroconidia from the same conidiogenous locus (
Figure 14K–O).
Two other
Bahusutrabeeja species,
B. bunyensis [
139] and
B. globosa [
126], closely resemble
C. dwaya in having globose to subglobose, multi-setulate conidia.
Bahusutrabeeja bunyensis differs in having smaller conidia with only 2–3 setulae, while
B. globosa has larger conidia compared to
C. dwaya. The conidia of
B. angularis [
140] have an angular outline and should be compared with
Nawawia [
141]. Other species currently referred to
Bahusutrabeeja differ in conidial shape and lack setulae, and their taxonomic treatment needs to be resolved using phylogenetic arguments.
Codinaea ellipsoidea (Z.L. Luo, K.D. Hyde & H.Y. Su) Réblová & Hern.-Restr., comb. nov. MycoBank MB 842193.
Basionym. Dictyochaeta ellipsoidea Z.L. Luo, K.D. Hyde & H.Y. Su, Fungal Divers. 99: 593. 2019.
For description, illustration, and comparison with morphologically similar species see Luo et al. [
29].
Habitat and geographical distribution: A saprobe on submerged wood of unidentified hosts, known only from Asia, China [
29]. The GlobalFungi database did not contain similar sequences (≥98%) of this species.
Notes: The species is known from several collections from China. It is characterized by the absence of setae and having dark pigmented, setiform conidiophores terminating into a monophialide with setulate ellipsoidal conidia.
≡ Dictyochaeta fertilis (S. Hughes & W.B. Kendr.) Hol.-Jech., Folia geobot. phytotax. 19: 426. 1984.
Culture characteristics: On CMD: colonies 75–77 mm, circular, flat, margin fimbriate, lanose, somewhat funiculose centrally, white-beige, reverse olivaceous beige. On MLA: colonies 66–68 mm diam, circular, convex, margin entire, lanose, floccose, with funiculose projections at the centre, becoming finely furrowed, grey-brown with a dark olivaceous brown outer zone, isabelline at the colony margin, reverse dark grey-brown. On OA: colonies >100 mm diam (colony reached the edge of 10 mm Petri dish), circular, flat, margin entire, lanose, olivaceous brown, camel brown toward the periphery with a dark olivaceous grey outer zone, reverse dark grey. On PCA: >100 mm diam (colony reached the edge of 10 mm Petri dish), flat, lanose, cobwebby toward the margin, beige, olivaceous beige at the margin, reverse dark olivaceous grey. Sporulation was abundant on all media after prolonged incubation (>6 weeks).
Colonies on PCA effuse, hairy, mycelium composed of branched, septate, subhyaline to pale brown hyphae 1.5–3 μm diam. Anamorph. Setae 152–340 μm long, 4.5–6.6 μm wide near the base, arise solitary or in group of 2–4 from dark brown stromatic cells, erect, straight or flexuous, septate, smooth, unbranched, rarely branched, dark brown, paler, and thinner-walled toward the apex, apex pale brown to subhyaline, with a terminal phialidic opening. Conidiophores 66–146 μm long, 4–5(–5.5) μm wide near the base, macronematous, single or arise in fascicles of 3–7 from dark brown hyphal cells around the bases of the setae, erect, straight or flexuous, septate, smooth, unbranched, occasionally branched in the upper part, pale to medium brown, thick-walled, gradually paler and thinner-walled toward the apex. Conidiogenous cells (15–)30–47 × 4.5–5.5(–6) μm tapering to 2–2.5 μm below the collarette, integrated, terminal, monophialidic, rarely polyphialidic with one lateral opening, extending percurrently (with up to five percurrent proliferations) or sympodially, subcylindrical, slightly swollen bellow the collarette, pale brown, subhyaline at the apex; collarettes 5.5–7.5 μm wide, 2.5–3 μm deep, funnel-shaped, subhyaline to pale brown. Conidia (10–)11–14.5 × 3.5–4.5 μm (mean ± SD = 12.8 ± 1.1 × 3.9 ± 0.4 μm), falcate to ellipsoidal-fusiform, slightly truncate at the base, with straight or gently curved setula at each end, 3.5–6.5(–9) μm long, conidia accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Specimens examined: FRANCE, Guadeloupe region, on root of Musa sp., 1966, J. Brun (culture CBS 242.66 = MUCL 15427). NEW ZEALAND, Auckland province, Waitakere Range, Waiatarua, on basis of dead leaves of Rhopalostylidis sapidae, 8 May 1963, S.J. Hughes 706c (isotype of Codinaea fertilis DAOM 93548c). NEW ZEALAND, on root of Betula sp., 1979, L. Mattson LEV 13960 (culture IMI 233824).
Habitat and geographical distribution: Based on literature records,
C. fertilis commonly occurs on decaying leaves, petioles, palm fronds, bark, wood, woody fruits and roots of
Alchornea cordifolia,
Calathea stromata,
Carya ovata,
Cedrela odorata,
Ctenanthe oppenheimiana,
Euterpe oleracea,
Fraxinus excelsior,
Freycinetia sp.,
Maranta bicolor,
Nothofagus menziesii,
Pandanus tectorius,
Pandanus sp.,
Quercus ilex,
Q. robur,
Quercus sp.,
Rhopalostylis sapida,
Stromanthe sanguinea and other unknown hosts in terrestrial and freshwater habitats. The species was reported from Africa, Australasia, Europe, South and North America, and Southeast Asia: Brazil, Brunei, Canada, Czech Republic, China, Ivory Coast, Malaysia, New Zealand, The Netherlands, Philippines, Slovak Republic, and USA [
2,
25,
26,
30,
35,
36,
37,
44,
121,
142,
143,
144,
145,
146,
147,
148]. Strains examined in this study were isolated from roots of
Betula sp. from New Zealand and
Musa sp. from France (Guadeloupe).
According to GlobalFungi, of all the fungi studied, this species is the most abundant worldwide in different climatic regions. The identical sequences were found in 655 samples, originating from 46 studies, collected on all continents including Antarctica. It was found in Africa (Ethiopia, Madagascar, South Africa, St. Helena island, Zambia, Zimbabwe), Asia (China, Malaysia, Papua New Guinea), East and South Australia, Europe (Northern Spain, Estonia, Germany, Austria, the Netherland, Slovenia), Hawaii, Central America and Central America (Mexico, Costa Rica, Puerto Rico, Trinidad and Tobago, Panama), North America (Oregon, Kansan, South Carolina, Minnesota, Tennessee, Luisiana, Georgia, Michigan), South America (Argentina). The sample types were soil (58%), rhizosphere (20%), root (15%) and shoot (6%) collected in cropland (37%), forest (30%), grassland (23%), woodland (7%), shrubland (2%) and desert (0.2%) habitats. The locations, with very few exceptions, have a tropical or temperate climate (MAT avg. 15 °C, MAP avg. 1066 mm).
Codinaeella fertilis has also been recorded as a plant pathogen causing root rot of
Lolium perenne,
Medicago sativa,
Trifolium repens,
T. praetense,
T. subterraneum and other important perennial pasture legumes in Australasia and North America: New Zealand and USA e.g., [
54,
56,
149,
150,
151].
Notes: Hughes and Kendrick [
2] described
C. fertilis from leaf bases of
Rhopalostylidis sapidae in New Zealand with conidia 9–15.4 × 2–3 μm and setulae 5–10 μm long. The revision of the isotype (
Figure 15) revealed certain variability in the shape and width of conidia. Although conidia were mostly falcate, 13–15 × 2.5(–3) μm, some conidia were also ellipsoidal-falcate and somewhat wider, 10–13 × (2.5–)3(–3.5) μm. Our observations of
C. fertilis in culture are consistent with those of the isotype; conidia of the strain IMI 233824 were predominantly ellipsoidal-falcate, however wider (3.5–4.5 μm) when grown in culture (
Figure 16). Although
C. fertilis sporulated abundantly on all four media, conidiophores associated with setae were formed only on MLA and PCA.
In characters of conidia,
C. fertilis is comparable to other species such as
C. aristata [
1],
C. paniculata [
46], and
Dictyochaeta plovercovensis [
122].
Codinaea aristata differs by narrower conidia with two asymmetrical setulae at each end,
C. paniculata possesses conidia longer in their upper range with longer setulae and
D. plovercovensis has narrower conidia with longer setulae.
According to literature, C. fertilis is a common species with a worldwide distribution. It is possible, however, that the arrangement of setae and conidiophores in the bundles may have been misinterpreted or overlooked. In such cases, C. fertilis could be confused with Ca. lutea or Ca. parvilobata (this study). They differ in setae, which grow independently and conidiophores are interspersed among them. The suspicion that some strains might be misidentified as C. fertilis was confirmed by molecular data. Three of the analysed strains, initially identified as C. fertilis (CBS 624.77, ICMP 14613, and ICMP 15540), were confirmed as members of the genus Codinaeella and represent Ca. lutea.
Codinaea gonytrichodes Shearer & J.L. Crane, Mycologia 63: 245. 1971. (
Figure 11 and
Figure 17).
≡ Codinaeopsis gonytrichodes (Shearer & J.L. Crane) Morgan-Jones [as “gonytrichoides”], Mycotaxon 4: 167. 1976.
≡ Dictyochaeta gonytrichodes (Shearer & J.L. Crane) Kuthub. & Nawawi [as “gonytrichoides”], Mycol. Res. 95: 845. 1991.
≡ Dictyochaetopsis gonytrichodes (Shearer & J.L. Crane) Whitton, McKenzie & K.D. Hyde [as “gonytrichoides”], Fungal Divers. 4: 156. 2000.
Culture characteristics: On CMD: colonies 45–50 mm diam, circular, raised, margin entire to fimbriate, lanose, floccose, sparsely funiculose, cobwebby at the margin, whitish-brown, reverse dark brown. On MLA: colonies 75–78 mm diam, circular, raised, margin lobate, lanose, floccose, funiculose at the inoculation block, zonate, mouse grey centrally, paler toward the periphery dark olivaceous grey at the margin, aerial mycelium with numerous miniature colorless exudates, reverse dark grey to black. On OA: colonies 72–75 mm diam, circular, flat, margin lobate, cobwebby centrally, lanose toward the periphery, brown at the colony centre, beige-grey toward the periphery, reverse dark beige centrally, dark grey to black toward the margin. On PCA: colonies 75–76 mm diam, circular, flat, raised centrally, margin fimbriate, lanose, floccose, cobwebby at the margin, becoming mucoid locally, whitish-beige, olivaceous brown at the margin, reverse dark olivaceous brown. Sporulation was abundant on MLA, OA, and PCA, sparse on CMD.
Colonies on OA effuse, hairy, mycelium composed of branched, septate, subhyaline to brown hyphae that become encrusted upon aging, 2.5–4 μm diam. Anamorph. Setae absent. Conidiophores 160–355 μm long, 4–7 μm wide above the swollen base, gradually tapering toward the apex, macronematous, arise singly or in groups of 2–3 from knots of hyphal cells, erect, straight or slightly flexuous, septate, smooth, brown, nodose hyphae are formed laterally below the septa, pale to medium brown. Conidiophores unbranched, sometimes branched in the upper part; branches 28–45(–61) × 3–5 μm arise from the nodose and collar-like hyphae, apex pale brown to subhyaline, sterile or terminating into a mono- or polyphialide. Conidiogenous cells 11.5–17 × 4–6 μm tapering to 1.5–2 μm below the collarette, mono- occasionally polyphialidic with 1–3 lateral openings, extending sympodially, discrete, lateral, arise as stalked or sessile phialides on collar-like and nodose hyphae or directly on conidiophores, mostly in groups of (2–)3–6(–7), or integrated, terminal at the apex of the conidiophore or phialide-bearing branches, subcylindrical to lageniform, pale brown, subhyaline at the apex; collarette 3–4 μm wide, 1.5–2(–2.5) μm deep, funnel-shaped, subhyaline to pale brown. Conidia 12–14.5 × 2–2.5(–3) μm (mean ± SD = 13.4 ± 0.6 × 2.4 ± 0.3 μm), falcate, tapering toward both ends, slightly truncate at the base with an inconspicuous scar, aseptate, hyaline, with straight or gently curved setula at each end 7.5–11.5 μm long, inserted terminally at the apex, subterminally at the base, conidia accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Specimen examined: JAPAN, Saitama Prefecture, Ogawa-machi, Seikoji Temple, decaying plant material, 4 September 1993, W. Gams & G. Okada (culture CBS 593.93).
Habitat and geographical distribution: Saprobe on decaying leaves, seeds, herbaceous stems and fruits of
Acer sp.,
Carya sp.,
Castanopsis cuspidata,
Liriodendron tulipifera,
Ochroma pyramidale,
Rubus sp. and other unknown hosts in freshwater and terrestrial habitats. The species is found in Asia and South and North America: Brazil, Japan, Malaysia and USA [
6,
21,
35,
37,
42,
127,
152,
153]. Occasionally it occurs on submerged wood of
Ochroma pyramidale (balsa wood) in USA [
6]. Rambelli et al. [
35] recorded
C. gonytrichodes on leaf litter of
Combretum dolichopetalum,
Manniophyton fulvum,
Newtonia duparquetiana, and
Xylopia acutiflora from Africa, Ivory Coast.
According to GlobalFungi, the identical sequences were found in 10 soil samples from forests in Asia in Iran and East China [
115,
117], and North America in Ohio and Georgia [
119,
154]. The sites have a temperate or humid continental climate (MAT avg. 15 °C, MAP avg. 1096 mm).
Notes: For descriptions and illustrations on natural substrate, see Shearer and Crane [
6], Morgan-Jones [
42] and Kuthubutheen and Nawawi [
21]. The morphological characteristics of conidiophores, conidiogenous cells and conidia of the strain CBS 593.93 match the protologue in all details and are well-comparable with other known specimens of this species. Only conidia of CBS 593.93 were slightly broader in vitro, 2–2.5(–3) μm wide, compared to those in nature, 1.2–2.3 μm [
6], 1.5–2.5 μm [
42], and 1.5–2.5 μm [
21].
Codinaea gonytrichodes is unique among other
Codinaea in having nodose hyphae at the septa, from which lateral phialides and branches grow. In older cultures, the apex is modified into a polyphialide and discrete, lateral polyphialides frequently occur in the upper part of the conidiophore.
Dictyochaetopsis polysetosa [
127] closely resembles
C. gonytrichodes in conidial morphology and branched conidiophores, but branches of the former species are setose, acicular, dark brown to black, arise in verticilli from nodose hyphae in the upper part of the conidiophore and remain sterile.
Dictyochaeta pahangensis [
21] is similar to
C. gonytrichodes in having knots of hyphae at the septa along the conidiophore axis, but differs in the absence of branches, longer conidia and nodose hyphae that are bulbose and dark brown with 1–2 discrete phialides.
Codinaea lignicola (Z.L. Luo, H.Y. Su & K.D. Hyde) Réblová & Hern.-Restr., comb. nov. MycoBank MB 842194.
Basionym. Dictyochaeta lignicola Z.L. Luo, H.Y. Su & K.D. Hyde, Fungal Divers. 99: 595. 2019.
For description, illustration and comparison of
C. lignicola with similar species, see Luo et al. [
29].
Habitat and geographical distribution: Saprobe on submerged wood, known only from Asia, China [
29]. According to GlobalFungi, the identical sequences were found in 12 samples of rhizosphere soil (92%) or soil (8%) collected in forest (92%) or grassland (8%) habitats in Asia in North East China and South Korea [
155,
156]. The locations have a cold and humid climate (MAT avg. 8 °C, MAP avg. 1285 mm).
Notes: Among Codinaea species, C. lignicola resembles C. ellipsoidea with simple, dark pigmented conidiophores terminated by monophialidic conidiogenous cells and absence of setae, but differs in having falcate vs. ellipsoidal conidia.
Codinaea pandanicola (Tibpromma & K.D. Hyde) Réblová & Hern.-Restr., comb. nov. MycoBank MB 842195.
Basionym. Dictyochaeta pandanicola Tibpromma & K.D. Hyde, Fungal Divers. 93: 127. 2018.
For description, illustration of
C. pandanicola and its comparison with similar species, see Tibpromma et al. [
48].
Habitat and geographical distribution: Saprobe on decaying leaves of
Pandanus sp., known only from Southeast Asia, Thailand [
48]. According to GlobalFungi, the identical sequences were found in 12 soil and one root samples from forests of Central America and the Caribbean (10 samples, Puerto Rico, Panama, Trinidat and Tobago, French Guiana) [
119], North America (1 sample, Georgia) [
157] and Asia (1 sample, Malaysia) [
158]. The locations have tropical climate (MAT avg. 21 °C, MAP avg. 2955 mm).
Notes: Although the description of
C. pandanicola did not include setae, on the photograph accompanying the protologue ([
48], Figure 95b), a dark brown, thick-walled seta (broken) terminating into a phialide is accompanied by shorter, paler, and thinner-walled conidiophores terminating into mono- or polyphialides.
Codinaea paniculata Réblová & J. Fourn., MycoKeys 74: 14. 2020. (
Figure 18).
For description, characteristics in culture, illustrations, and comparison with similar species, see Réblová et al. [
46].
Habitat and geographical distribution: Saprobe on decaying wood and submerged leaves of
Alnus glutinosa,
Fraxinus excelsior and other unidentified hosts in Europe in France and United Kingdom [
46]. Based on comparison of the ITS sequences of the ex-type strain with environmental sequences deposited in GenBank,
C. paniculata was also isolated as a root endophyte from a beach grass
Elymus mollis in North America in the USA, Oregon (ITS: KU838460, KU839605) [
159] and from soil samples from an ancient woodland in the United Kingdom (ITS: KM374380) [
160].
According to GlobalFungi, the identical sequences were found in 111 samples from soil (64%) or roots (36%) in grassland (43%), forest (38%), cropland (10%), or woodland (8%) habitats. Ninety-two samples originate from Europe (Belgium, Estonia, Germany, Monte Negro, Northern Spain, South Slovenia, Sweden, Wales), five samples from Africa (St. Helena island), one from Asia (Malaysia), five samples from South America (North Argentina), and eight samples were from Southeast Australia and Tasmania. The samples originate from 13 studies and the sites have temperate climate (MAT avg. 10 °C, MAP avg. 881 mm).
Specimens examined: FRANCE, Ariège, Pyrénées Mts., Rimont, La Maille brook, alt. 550 m, 28 May 2018 (incubated in moist chamber for 1 week), on submerged decaying wood, J. Fournier M.R. 3950 (holotype PRA-16319, ex-type culture CBS 145098). FRANCE, Ariège, Pyrénées Mts., Rimont, Le Baup stream, ca. 1.5 km from the village along D18 road, alt. 550 m, 12 June 2009, on submerged wood of Fraxinus excelsior, J. Fournier J.F. 09153 (PRA-16320, culture CBS 127692), Ibid., 23 May 2008, on submerged wood of Alnus glutinosa, J. Fournier & M. Delpont J.F. 08124 (PRA-16321, culture CBS 126573). UNITED KINGDOM, Liverpool, University Campus Liverpool, 1992, on submerged dead leaf in a pool, G.L. Hennebert (culture MUCL 34876).
Notes:
Codinaea paniculata represents the C1 morphotype. We observed variability among strains of
C. paniculata from France (CBS 127692, CBS 145098) and the United Kingdom (MUCL 34876), which is shown in
Figure 18. None of the three isolates produced pigments diffusing into the agar. The most prominent growth and abundant aerial mycelium, sometimes with submerged growth, was observed on MLA and OA. The growth on CMD and PCA was rather sparse.
Codinaea phasma Hern.-Restr. & Réblová,
sp. nov. MycoBank MB 842198. (
Figure 11 and
Figure 19).
Etymology: Phasma (L) ghost, phantom, referring to the mysterious appearance of the multisetulate conidia in culture.
Typification: PUERTO RICO, on decaying twig of an unidentified plant, 19 July 2018, M. Hernández-Restrepo M.H.R. 18014 (holotype CBS H-24747, culture ex-type CBS 147516).
Description on the natural substrate: Colonies effuse, hairy, black. Anamorph. Setae 160–380 μm long, 5–10 μm wide near the base, gradually tapering upwards, arise solitary or in groups of two from dark brown stromatic cells, erect, straight or flexuous, septate, smooth, dark brown, thick-walled, paler, and thinner-walled toward the apex, unbranched, apex pale brown with a terminal or one to several lateral phialidic openings. Conidiophores 35–97 μm long, 2.5–4.5 μm wide near the base, macronematous, single or arise in fascicles of 2–5 from stromatic cells around the base of the setae, erect, straight or slightly bent, septate, smooth, medium to pale brown, paler toward the apex.
Conidiogenous cells 16–43 × 2.5–4 μm, 5–7 μm wide at the broadest point, tapering to 1.5–2.5 μm below the collarette, integrated, terminal, mono- or polyphialidic, occasionally extending percurrently, subcylindrical, swollen bellow the collarette, pale brown; collarettes 2–3.5 μm wide, 2–3 μm deep, funnel-shaped to slightly tubular, pale brown. Conidia 13.5–18 × 3–4 μm (mean ± SD = 15.1 ± 3.8 × 1.2 ± 0.4 μm), oblong to falcate, slightly truncate at the base with an inconspicuous basal scar, aseptate, hyaline, with a straight or gently curved setula at each end 5–13 μm long, inserted terminally at the apex, subterminally at the base, conidia accumulate in whitish slimy fascicles. Teleomorph. Unknown.
Culture characteristics: On CMD: colonies 27–30 mm diam, circular, flat to slightly convex, margin fimbriate, lanose, floccose, whitish-beige with a darker olivaceous beige zone at the margin, pale ochre pigment diffusing into the agar, reverse of the same colors. On MLA: colonies 14–17 mm diam, circular, raised, margin lobate, lanose becoming mucoid, aerial mycelium restricted to the centre of the colony, deeply furrowed with nearly cerebral-like folds, whitish centrally, pink-beige toward the margin, pale apricot pigment diffusing into the agar, reverse apricot. On OA: colonies 20–22 mm diam, circular, raised, margin undulate, weakly furrowed, lanose, aerial mycelium with colorless exudates, whitish-grey, olivaceous to mouse grey toward the margin with a whitish outer zone, pale ochre pigment diffusing into the agar, reverse apricot-grey. On PCA: colonies 42–55 mm diam, circular, slightly convex centrally with flat margin, margin entire to weakly fimbriate, lanose, whitish centrally, brown to olivaceous-brown toward the periphery, darker at the margin, reverse dark brown. Sporulation was abundant on MLA, OA, and PCA, sparse on CMD.
Colonies on CMA with U. dioica stems effuse, hairy, mycelium composed of branched, septate, hyaline to pale brown hyphae 2–4.5 μm diam. Setae, conidiophores and conidiogenous cells similar to those from nature. Setae 97–236 μm long, 3.5–6 μm above the swollen base, dark olivaceous brown, paler toward the apex, apex pale brown to subhyaline, with a terminal phialidic opening. Conidiophores 44–104 μm long, 3.5–7 μm wide above the base, arise singly or in groups, interspersed among the setae, medium brown to olivaceous brown, paler toward the apex. Conidiogenous cells 24–34 × 3.5–5 μm, 4.5–7.5 μm wide at the broadest part, tapering to 1.5–4 μm below the collarette; collarette 2–6 μm wide, 1.5–8 μm deep, funnel-shaped to tubular, subhyaline. Conidia 12–18.5 × 4.5–8 μm (mean ± SD = 14.3 ± 5.6 × 1.5 ± 1.0 μm), irregularly ellipsoidal, tapering toward both ends, sometimes slightly contracted near the base, basal end truncate to obtuse with an inconspicuous basal scar, aseptate, hyaline, with 3(–5), straight or gently curved setulae 3.5–11 μm long, inserted terminally at the apex, subterminally at the base and also dorsally and ventrally, accumulate in slimy whitish fascicles.
Habitat and geographical distribution: Saprobe on decaying wood, known only from the Caribbean from Puerto Rico. According to GlobalFungi, the identical sequences were found in two soil samples from forest in Puerto Rico [
118]. The sites have tropical climate (MAT avg. 22.45 °C, MAP avg. 3013).
Notes: On natural substrate, C. phasma forms typical bundles of setae and conidiophores and oblong to falcate conidia with a single, simple setula at each end. When grown in culture, the conidia become irregularly ellipsoidal, slightly inflated in the middle and possess up to five setulae inserted also dorsally and ventrally and occasionally they can branch. It resembles C. terminalis, but the later species differs from C. phasma in longer, falcate conidia. In the present phylogeny, the two species are shown as unrelated lineages.
Codinaea siamensis (J. Yang, K.D. Hyde & J.K. Liu) Réblová & Hern.-Restr.,
comb. nov. MycoBank MB 842199. (
Figure 13 and
Figure 20).
Basionym. Dictyochaeta siamensis J. Yang, K.D. Hyde & J.K. Liu, Mycol. Prog. 15: 1159. 2016.
Culture characteristics: On CMD: colonies 35–42 mm diam, circular, flat, margin fimbriate, lanose becoming cobwebby, mucoid at the margin, beige-brown, reverse of the same color. On MLA: colonies 63–66 mm diam, circular, raised, margin entire, furrowed, lanose, funiculose, pale olivaceous grey, grey-brown at the margin, reverse dark brown. On OA: colonies 77–80 mm diam, circular, slightly raised, margin lobate, lanose, grey, olivaceous grey-brown at the margin, reverse dark grey to nearly black. On PCA: colonies 76–80 mm diam, circular, flat, margin entire, lanose to cobwebby, floccose, grey-brown, olivaceous brown at the margin, reverse dark olivaceous brown. The strain sporulated only on MLA after prolonged incubation (>6 weeks).
Colonies on MLA effuse, hairy, mycelium composed of branched, septate, hyaline to pale brown hyphae 1.5–3 μm diam. Anamorph. Setae 240–330 μm long, 6.5–9 μm wide near the base, arise singly or in groups of 2(–3), erect, straight or flexuous, septate, smooth, dark brown, thick-walled, paler and thinner-walled toward the apex, unbranched, apex pale brown to subhyaline, with a terminal or 1–2 lateral phialidic openings. Conidiophores 67–125(–153) μm long, 3.5–5 μm wide near the base, macronematous, single or arise in fascicles of 3–6 around the base of the setae, unbranched, erect, straight or flexuous, septate, smooth, medium brown, paler toward the apex. Conidiogenous cells 12.5–30 × 3.5–4.5(–5) μm tapering to 1.5–2.5 μm below the collarette, integrated, terminal, mono- or polyphialidic with 1–3 lateral apertures while internally septa can be formed, extending percurrently and sympodially, cylindrical, pale brown, subhyaline at the apex; collarettes 3.5–5 μm wide, 2–2.5(–3) μm deep, funnel-shaped, subhyaline, apical part evanescent with age. Conidia 11.5–14.5 × 2.5–4 μm (mean ± SD = 12.8 ± 0.8 × 3.1 ± 0.3 μm), suballantoid to oblong, slightly curved, asymmetrical, slightly obtuse at the base with an inconspicuous basal scar, aseptate, hyaline, with straight or gently curved setula at each end (4.5–)5–8.5(–10) μm long, inserted terminally at the apex, subterminally at the base, conidia accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Specimen examined: PAPUA NEW GUINEA, Madang Province, Finisterre Range, soil in the tropical rain forest, November 1995, A. Aptroot, (culture CBS 194.96).
Habitat and geographical distribution: This species occurs in soil, on submerged leaves of
Pandanus sp. and decaying wood of an unidentified host, and is known from Australasia and Southeast Asia: Papua New Guinea and Thailand ([
47,
48], this study).
According to GlobalFungi, the identical sequences were found in 55 samples of soil (89%) or root (11%) collected in forest (98%) and freshwater aquatic (2%) habitat in Central America and the Caribbean (30 samples, Puerto Rico, Panama), South America (eight samples, Brazil, French Guyana, Trinidad and Tobago), Asia (12 samples, South China, Papua New Guinea, Thailand, Malaysia), Australia (four samples, Tasmania), Africa (one sample, Madagascar) and North America (one sample, Minnesota), originating from 13 studies. The localities mostly have tropical or temperate climate (MAT avg. 23 °C and MAP avg. 2319 mm).
Notes:
Codinaea siamensis has been reported from two collections, which differ in shape and length of conidia and setulae. In the ex-type strain MFLUCC 15-0614, conidia were described as falcate, 15.5–21 × 2.5–4 μm with setulae 7–12 μm long [
47], while in the other isolate MFLUCC 16-0371, conidia were allantoid, cylindrical or long fusiform and shorter, 8–17 × 2–5 μm with setulae 1–10 μm long [
48]. Conidia observed in the strain CBS 194.96 on MLA correspond to the shape and size of conidia given for the non-type collection of
C. siamensis by Tibpromma et al. [
48]. The conidiophores formed in culture were longer than those from nature due to the phialides, which frequently elongated percurrently. The ITS sequences of all three strains of
C. siamensis are nearly identical and correlate with 99.57% sequence identity, which is equal to a difference of two base pairs between two sequences. The
tef1-α sequence identity between MFLUCC 16-0371 and CBS 194.96 was 99.78%; the
tef1-α sequence of the ex-type strain was not available.
Codinaea paniculata [
46] is similar to
C. siamensis, but differs in shorter setae and narrower conidia.
Codinaea siamensis is also comparable to
C. terminalis, which differs in shorter setae, monophialidic conidiogenous cells and longer conidia in their upper range.
Codinaea terminalis (C.G. Lin & K.D. Hyde) Réblová & Hern.-Restr., comb. nov. MycoBank MB 842200.
Basionym. Dictyochaeta terminalis C.G. Lin & K.D. Hyde, Mycosphere 10: 672. 2019.
For description and illustration, see Lin et al. [
30].
Habitat and geographical distribution: Saprobe on decaying leaves, known so far from Asia, China [
30]. According to GlobalFungi, the identical sequences were found in two soil samples from the cropland and forest habitats in South China [
161,
162]. The localities have a temperate climate (MAT avg. 20 °C, MAP avg. 1719 mm).
Notes:
Codinaea terminalis is characterized by setae and conidiophores growing together in bundles (C1 morphotype) and is similar to
C. assamica and
C. siamensis. However, the two latter species differ in having longer setulae and wider conidia. For detailed comparison with other species, see
Table S4.
Codinaea vermispora (Z.F. Yu & R.F. Castañeda) Réblová & Hern.-Restr., comb. nov. MycoBank MB 842201.
Basionym. Phialolunulospora vermispora Z.F. Yu & R.F. Castañeda, MycoKeys 76: 23. 2020.
For description and illustration, see Zheng et al. [
50].
Habitat and geographical distribution: The species was isolated from submerged leaves of a dicotyledonous plant and is known only from Asia, China [
50]. The GlobalFungi database does not contain similar (≥98%) sequences of this species.
Notes:
Codinaea vermispora was initially accommodated in the monotypic genus
Phialolunulospora [
50] and distinguished from
Codinaea by vermiform to sigmoid conidia bearing an eccentric basal appendage. Although the authors emphasized the single basal appendage of the conidium, the photograph ([
50],
Figure 2) shows that the tip of the conidium is strongly attenuated and the transparency and demarcation of this region from the rest of the conidium is identical to the basal appendage. Phylogenetic analysis suggests placement of this species in the strongly supported
Codinaea clade. Characteristics of conidia (shape, size and the basal appendage) cause
C. vermispora to be well distinguished among other members of the genus.
3.5.2. The Genus Codinaeella
Codinaeella Réblová & Hern.-Restr., gen. nov. MycoBank MB 842000.
Type species. Codinaeella minuta (Tubaki) Réblová & Hern.-Restr.
Etymology: Codinae- and -ella (L) diminutive but here used as a name-forming suffix, referring to fungi morphologically similar to Codinaea.
Description: Colonies on natural substrate effuse, lanose, brown to reddish-brown, composed of conidiophores and setae, mycelium semi-immersed or immersed. Anamorph. Setae present or occasionally absent, grow singly or in small groups from repent hyphae or knots of hyphal cells, erect, straight or flexuous, septate, brown, unbranched, always fertile with a terminal or several lateral phialidic openings, setae rarely absent. Conidiophores macronematous, mononematous, crowded, arise singly or in groups from repent hyphae or knots of hyphal cells, scattered among the setae if present, unbranched, occasionally branched, erect, straight or flexuous, sometimes geniculate, brown, septate, smooth. Conidiogenous cells integrated, terminal on conidiophores or short phialide-bearing branches, or discrete, lateral on conidiophores or 1–2-celled stalks, mono- and polyphialidic, extending percurrently and sympodially, pigmented, often with persistent remnants of the collarettes; collarettes flared, funnel-shaped, the apical part may become soon evanescent. Conidia falcate, cylindrical-fusiform, curved, slightly asymmetrical, tapering toward both ends, slightly truncate at the base with an inconspicuous scar, aseptate, hyaline, with straight or gently curved setula at each end inserted terminally at the apex and subterminally at the base, accumulate in slimy fascicles. Teleomorph. Unknown.
Habitat and geographical distribution: Members of Codinaeella are saprobes, found on decaying bark, wood, woody fruits, leaves, petioles or palm fronds of various plants, but also in soil and living roots, and have a worldwide distribution in Holarctic realm, but also occur in the subtropical and tropical geographic zones of Africa, Asia, Australasia, and South America.
Notes: Two morphotypes were found among
Codinaeella species (
Figure 8). Species with the predominant morphotype CA1 (e.g.,
Ca. minuta,
Ca. lutea,
Ca. parvilobata) form unbranched conidiophores that grow singly or in small groups from repent hyphae or knots of hyphal cells and are usually scattered among longer, darker and thicker-walled unbranched setae. The setae resemble conidiophores; they are always fertile and terminate into a mono- or polyphialide. Although conidiophores are simple on material from nature, in
Ca. minuta and
Ca. lambertiae conidiophores often branch in culture. The other morphotype CA2 is less widespread and it is represented by
Ca. filamentosa only. It is characterized by single, branched conidiophores with a sterile setiform extension and lower fertile part, and they are accompanied by shorter, unbranched conidiophores. Conidiogenous cells are monophialidic, discrete, lateral and also integrated, terminal on stalks or short phialide-bearing branches and shorter conidiophores. The teleomorph of
Codinaeella is unknown.
Eight species were accepted in
Codinaeella and all were verified with DNA sequences. Several other species closely resemble
Codinaeella and although their systematic placement is unknown and molecular data are unavailable, their inclusion in
Codinaeella should be considered based on future phylogenetic arguments. Species such as
Dictyochaeta gamundiae [
163],
D. pakhalensis [
164], and
D. taiwanensis [
165] match the CA1 morphotype.
Codinaea sinensis [
166],
Dictyochaetopsis (
Di.)
elegantissima,
Di. intermedia [
32] and
Di. menisporoides [
26] correspond to the CA2 morphotype with branched conidiophores and discrete phialides.
Codinaeella is separated from
Codinaea by molecular and morphological characters. The two genera differ in the arrangement of setae (if present) and conidiophores on the natural substrate and partly also in the production of pigments in vitro. The morphology of
Codinaea is more complex and consists of four morphotypes, but none of them is comparable with those of
Codinaeella.
Codinaeella often forms bright yellow, ochre, orange to dark burgundy pigments dispersed in agar, whereas in
Codinaea the pigments are rarely formed. A synopsis table with diagnostic features of accepted species of
Codinaeella is provided in the
Supplementary File: Table S5.
Codinaeella filamentosa (Onofri) Réblová & Hern.-Restr.,
comb. nov. MycoBank MB 842202. (
Figure 21 and
Figure 22).
Basionym. Codinaea filamentosa Onofri, Mycotaxon 14: 120. 1982.
≡ Dictyochaetopsis filamentosa (Onofri) Aramb. & Cabello, Mycotaxon 38: 12. 1990.
Description on the natural substrate: Colonies effuse, grey-brown, composed of conidiophores, mycelium immersed. Anamorph. Setae comparable to the upper layer of branched setiform conidiophores. Conidiophores macronematous, grow singly or in groups of 2–3, erect, straight or flexuous, septate, brown, smooth, form two distinct layers. Conidiophores of the upper layer 252–275 μm long, 3.5–4 μm wide near the base, branched, the upper part of the conidiophore is sterile with a setiform extension, apex obtuse and subhyaline, lower part of the conidiophore is fertile, lateral branches and grow just below the septa, they are either sterile and similar to the main stalk, tapering, up to 166 μm long, or shorter phialide-bearing branches 29.5–51 × 3.5–4 μm. Conidiophores of the lower layer 28–85 μm long, 3–4 μm wide, pale brown, paler toward the apex, unbranched with a terminal phialidic opening, occasionally with discrete phialides. Conidiogenous cells 13–35 × 2.5–3.5 μm tapering to 1.5–2 μm below the collarette, integrated, terminal or discrete, lateral, monophialidic, rarely polyphialidic with one lateral opening, elongating percurrently and sympodially, cylindrical, pale brown, subhyaline toward the apex; collarettes ca. 3 μm wide, 1.5–2 μm deep, funnel-shaped, hyaline. Conidia (13.5–)14.5–17.5(–18) × 2–2.5 (mean ± SD = 16 ± 1.6 × 2.5 ± 0.2 μm), falcate, tapering toward both ends, aseptate, hyaline, with a straight or gently curved setula at each end, 5–8 μm long, inserted terminally at the apex, subterminally at the base, conidia accumulate in slimy whitish fascicles.
Culture characteristics: On CMD: colonies 35–38 mm diam, circular, flat, margin entire, velvety, later mucoid and smooth, whitish-brown, beige toward the margin, reverse beige with a dark brown zone. On MLA: colonies 25–27 mm diam, circular, flat, raised margin, margin entire to weakly undulate, mucoid-waxy to cobwebby, lanose at the margin, colony centre dark burgundy-brown, whitish-grey to grey at the margin with a dark ochre outer zone, dark burgundy (centre) and pale ochre pigment (margin) diffusing into agar, reverse dark burgundy to black. On OA: colonies 50–53 mm diam, circular, flat, slightly raised, margin entire, mucoid, velvety to cobwebby centrally and at the margin, zonate, irregularly dark brown at the centre with beige zones of sparse growth, reverse brown. On PCA: colonies 40–42 mm diam, circular, flat, margin entire, cobwebby, slightly funiculose becoming mucoid, zonate, beige-brown, isabelline at the margin, reverse of the same colors. Sporulation was sparse on MLA, OA, PCA, absent on CMD.
Colonies on MLA effuse, mycelium composed of branched, septate, pale brown hyphae 2–2.5 μm diam. Conidiophores 160–493 μm long, 2.5–3.5 μm wide near the base, tapering upwards, sometimes with a sterile, dark brown, setiform extension up to 377 μm long, apex narrowly rounded, erect, straight or flexuous, septate, smooth, medium brown, simple or branched; phialide-bearing branches arise in the lower part of the conidiophore, secondary and tertiary branches formed frequently, parallel, extending upwards percurrently, slightly curved downwards at the apical part. Conidiogenous cells 18–42 × 3–3.5 μm tapering to 1.5–2 μm below the collarette, monophialidic, pale brown to subhyaline; collarettes 3–4 μm wide, 1.5–2.5 μm deep, subhyaline, funnel-shaped, apical part often evanescent. Conidia similar to those from nature, 13.5–17 × 2–3 μm (mean ± SD = 15 ± 0.9 × 2.4 ± 0.3 μm), falcate, setulae 5.5–8.5 μm long, accumulate in slimy whitish fascicles.
Specimen examined: USA, Louisiana, Baton Rouge, Burden Plantation, on buried leaves of Quercus sp., 11 February 2002, K.A. Seifert K.A.S. 1504 (culture CBS 147265).
Habitat and geographical distribution: A saprobe on leaf litter of
Anthonotha fragrans,
Calpocalyx brevibracteatus,
Chrysophyllum taiense,
Quercus sp. and other unknown hosts, known from Africa and North America: Ivory Coast and the USA, Louisiana ([
32,
35], this study).
According to GlobalFungi, this species was found in six soil samples from forest habitat in Eastern North America in New York, Massachusetts [
119,
167]. The sites have temperate or humid continental climate (MAT avg. 10 °C, MAP avg. 1238 mm). In addition,
Ca. filamentosa is identical with
Ca. minuta in the ITS1 barcode. Thus, only the ITS2 region was used to study its biogeography.
Notes: On the natural substrate, Ca. filamentosa forms two kinds of conidiophores, of which the longer are branched and have a sterile setiform extension; the branches are also sterile and similar to the main stalk. These longer conidiophores can be considered equivalent to the setae (although, apically always fertile) observed in other species of Codinaeella, such as Ca. lutea, Ca. mimusopis, Ca. minuta and Ca. parvilobata. Codinaeella filamentosa has a distinctive characteristic of colony on MLA. The flat, central part of the colony is dark burgundy-brown, mucoid becoming cobwebby in older cultures. Aerial mycelium is usually confined to the periphery where it is abundant, lanose and forms a raised, grey margin. During the analysis of the secondary metabolites, we identified naphthoquinones bioactive compounds produced from the central pigmented part of the colony (A. Čmoková et al., in preparation).
Four other species, such as
C. sinensis,
Dictyochaetopsis elegantissima,
Di. Intermedia, and
Di. menisporoides, resemble
Ca. filamentosa in having discrete, lateral phialides, falcate, aseptate, hyaline conidia with setulae and sometimes branched conidiophores. Sometimes longer conidiophores are accompanied by shorter simple conidiophores or stalked phialides.
Dictyochaetopsis elegantissima differs from
Ca. filamentosa in conidia with bifid setulae at each end [
32].
Dictyochaetopsis intermedia differs from
Ca. filamentosa in having conidiophores with a terminal phialidic opening and wider conidia.
Codinaea sinensis has unbranched conidiophores and shorter conidia [
166] compared to
Ca. filamentosa.
Dictyochaetopsis menisporoides differs from
Ca. filamentosa in polyphialidic conidiogenous cells, unbranched conidiophores with mostly sessile phialides, sometimes formed on branches [
26]. The systematic placement of these species is unknown; the living cultures or molecular data are not available.
Codinaeella lambertiae (Crous) Réblová & Hern.-Restr.,
comb. nov. MycoBank MB 842203. (
Figure 22 and
Figure 23).
Basionym. Codinaea lambertiae Crous, Persoonia 39: 399. 2017.
Culture characteristics: On CMD: colonies 62–64 mm diam, circular, flat, margin entire, mucoid to cobwebby, brown centrally, beige with irregular brown spots, reverse of the same color. On MLA: colonies 57–58 mm diam, circular, convex, margin entire, sparsely lanose, mucoid at the margin, zonate, furrowed, white-ochre with irregular dark brown to orange-brown intermediate zones, with an isabelline outer zone of submerge growth, reverse amber with dark brown rings. On OA: colonies 60–62 mm diam, circular, raised, margin entire, velvety to cobwebby, floccose, locally mucoid with a sparse growth, pink-brown, whitish-brown toward the margin sometimes with orange-brown outer zone, light ochre-pink pigment diffusing into the agar, reverse ochre-yellow to yellow-orange. On PCA: colonies 59–60 mm diam, circular, flat, margin entire, cobwebby to mucoid, zonate, beige, locally olivaceous beige at the margin, reverse beige. Sporulation was moderate on MLA, OA, and PCA, absent on CMD.
Colonies on OA effuse, mycelium composed of branched, septate, hyaline to pale brown hyphae 1.5–2.5 μm diam. Anamorph. Setae absent. Conidiophores 128–362 μm long, 2.5–3 μm wide near the base, usually tapering toward the base, macronematous, single, simple or branched, erect, septate, straight or flexuous, geniculate toward the apex due to numerous lateral phialidic openings, smooth, pale brown. Conidiogenous cells 19–34 × 3–3.5 μm tapering to 1.5–2(–2.5) μm below the collarette, integrated, terminal, mono- and polyphialidic with 1–6 lateral openings while internally septa can be formed, extending percurrently and sympodially, cylindrical, pale brown; collarettes 4–5.5 × 2–3(–4) μm, funnel-shaped, pale brown to subhyaline. Conidia 11.5–14.5 × 2.5–3 μm, falcate, tapering toward both ends, slightly truncate at the basal end with an inconspicuous scar, aseptate, hyaline, with a straight or gently curved setula at each end, 4–6.5 μm long, inserted terminally at the apex, subterminally at the base, accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Specimen examined: AUSTRALIA, New South Wales, Fitzroy Falls, Morton National Park, on leaves of Lambertia formosa, 26 November 2016, P.W. Crous (culture ex-type CBS 143419).
Habitat and geographical distribution: Saprobe on fallen leaves of
Lambertia formosa, known from Australia only [
10]. According to GlobalFungi, this species was found mostly in Southeast Australia and Tasmania in 14 samples, mostly in soil in forest or woodland environments [
117,
168,
169]. A single soil sample originates from Asia (East China) from a cropland biome. The sites have temperate or humid continental climate (MAT avg. °C, MAP avg. 1091 mm).
Notes: When grown on OA, the conidiophores of
Ca. lambertiae (CBS 143419) branch; the branches are fertile and curved upwards. Similar branching pattern was observed in
Ca. filamentosa and
Ca. minuta (this study). Phialides extend percurrently and later also sympodially and the upper part of the conidiophore acquires a slightly geniculate appearance. In the protologue of
Ca. lambertiae [
10], the conidiophores were described simple, conidiogenous cells monophialidic, rarely with a lateral phialidic aperture and the conidia were given longer ((13–)14–15(–18) × (2.5)–3 μm. Because of these differences we present a description of
Ca. lambertiae on OA.
Codinaeella lambertiae is similar to Ca. parvilobata but differs in having slightly longer and wider conidia with shorter setulae. While Ca. lambertiae mainly occurs in Australasia, Ca. parvilobata has a wide range of distribution.
Codinaeella lutea Réblová & Hern.-Restr
., sp. nov. MycoBank MB 842204. (
Figure 24 and
Figure 25).
Typification: CZECH REPUBLIC, South Bohemian region, Novohradské hory Mts., Horní Stropnice, Bedřichovský potok Natural Monument, on decaying cupule of Quercus sp., 5 October 2018, M. Réblová M.R. 3982 (holotype PRA-20986, culture ex-type CBS 146618).
Etymology: Luteus (L) yellow, referring to the yellow pigment diffusing into agar, and staining conidial masses and aerial mycelium in vitro.
Description on the natural substrate: Colonies effuse, grey-brown, composed of setae and conidiophores, mycelium semi-immersed. Anamorph. Setae 125–250 μm long, (3.5–)4.5–5.5 μm wide near the swollen base, grow singly or in groups, erect, straight or flexuous, septate, smooth, dark brown and thick-walled, paler and thinner-walled toward the apex, unbranched, always fertile, apex pale brown to subhyaline with a single terminal or 1–5 lateral phialidic openings. Conidiophores 39–102 μm long, 3.5–4.5 μm wide near the base, macronematous, crowded, arise singly or in groups of 2–3, scattered among the setae, unbranched, erect, straight or flexuous, slightly geniculate in the apical part, septate, smooth, pale to medium brown, paler toward the apex. Conidiogenous cells 19.5–29(–32) × 3.5–4.5 μm tapering to 1.5–2 μm below the collarette, integrated, terminal, mono- or polyphialidic with 2–8 lateral openings while internally septa can be formed, extending sympodially, cylindrical, slightly swollen below the collarette, pale brown, subhyaline at the apex, usually bearing persistent remnants of the collarettes; collarettes 3–3.5 μm wide, 2–2.5 μm deep, funnel-shaped, subhyaline to pale brown, the apical part soon evanescent. Conidia 12.5–17.5 × 2–2.5(–3) μm (mean ± SD = 15.6 ± 1.4 × 2.6 ± 0.2 μm), falcate, asymmetrical, tapering toward both ends, slightly truncate at the base with an inconspicuous basal scar, aseptate, hyaline, with straight or gently curved setula at each end (4.5–)5–8.5(–10) μm long, inserted terminally at the apex, subterminally at the base, conidia accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Culture characteristics: On CMD: colonies 35–40 mm diam, circular, flat, margin entire to fimbriate, sparsely lanose, somewhat funiculose, mucoid toward the periphery, whitish-beige centrally, pale cinnamon when mucoid, beige to isabelline toward the margin, pale yellow-gold pigment diffusing into agar, reverse cinnamon. On MLA: colonies 15–18 mm diam, circular, raised, margin entire to lobate, velvety-lanose becoming mucoid-waxy, glossy, furrowed, pink-beige, apricot to cinnamon with remnants of yellow to white-yellow aerial mycelium at the inoculation block, sometimes with a dark grey to dark burgundy outer zone when mucoid and beige-brown lanose margin, diffusing apricot pigment into the agar, more intense color at the margin, reverse ochre, apricot to cinnamon. On OA: colonies 60–65 mm, circular, raised, margin entire, velvety-lanose, floccose, occasionally partially mucoid, furrowed, sometimes ridges with small cracks, white to beige-brown centrally, dark brown in the mucoid part, with a yellow ring or irregular yellow patches of aerial hyphae with numerous yellow exudates, sometimes yellow to pale ochre pigment diffusing locally into agar, reverse orange-brown. On PCA: colonies 69–73 mm diam, circular, raised, margin entire to fimbriate, lanose, floccose, white-beige, sometimes mucoid and cinnamon centrally, with irregular yellow tufts of aerial mycelium, beige toward the margin, pale ochre pigment diffusing into agar, reverse olivaceous grey-brown to cinnamon. Sporulation was sparse on PCA, absent on CMD, MLA, and OA.
Colonies on CMA with U. dioica stems effuse, mycelium composed of branched, septate, hyaline to pale brown hyphae 1–2 μm diam. Anamorph. Setae absent, conidiophores, conidiogenous cells, and conidia similar to those from nature. Conidiophores 36–83 μm long, 2–4 μm wide above the base, usually tapering toward the swollen base, unbranched, erect, septate, smooth, pale brown to subhyaline, sometimes reduced to single conidiogenous cells. Conidiogenous cells 16–32 × 3.5–4.5 μm tapering to 1.5–2.5 μm below the collarette, integrated, terminal, mono-, occasionally polyphialidic with 1–2 lateral openings, subcylindrical, extending percurrently and sympodially, pale brown to subhyaline; collarettes 3.5–5 μm wide, 1.5–2.5 μm deep, funnel-shaped. Conidia 14.5–17 × 2–3 μm (mean ± SD = 15.8 ± 0.8 × 2.6 ± 0.3 μm), falcate, hyaline, aseptate, setulae 4–7.5 μm long, accumulate in slimy whitish or yellow fascicles. Teleomorph. Unknown.
Other specimens examined: NEW ZEALAND, Canterbury region, Christchurch, Christchurch Botanical Gardens, on a dead leaf of Quercus ilex, 2 September 2002, J.A. Cooper JAC8343 (PDD 80130, culture ICMP 14613). NEW ZEALAND, Tasman region, Murchison, Dough Boy Track, on dead cupule of Quercus sp., 15 May 2004, J.A. Cooper JAC9057 (PDD 80612, culture ICMP 15540). THE NETHERLANDS, Utrecht, Fort Rijnauwen, ingang (1b), on cupule of Quercus robur, 22 October 1977, W. Gams (CBS H-746, culture CBS 624.77).
Habitat and geographical distribution: Codinaeella lutea is a saprobe on decaying leaves and woody fruits of Quercus ilex, Q. robur, Quercus sp. And other unknown host. It is known from Australasia, Europe, and Southeast Asia: China, Czech Republic, New Zealand and The Netherlands.
According to GlobalFungi, the identical sequences were found in 53 samples of roots (72%), soils (26%), or water (2%) originating mostly from forest (81%) and grassland (13%) habitats in Europe (49 samples, Belgium Denmark, Wales) [
170,
171,
172], Asia (3 samples, Iran) [
115]. It was also found in the single water sample from North America (North Carolina) [
173]. The sites have the temperate climate (MAT avg. 11 °C, MAP avg. 85 mm).
Notes: The four strains of
Ca. lutea examined in this study produce yellow pigment, which diffuses into the agar and also stains aerial mycelium in vitro. This character is conspicuous in the ex-type strain on OA (
Figure 24K,L). In other isolates the yellow-colored hyphae formed sparsely, either on MLA or PCA (
Figure 25). In addition, yellow conidial masses formed when grown on
Urtica stems on CMA (
Figure 24M). The four isolates of
Ca. lutea from Asia, Europe and New Zealand have identical ITS sequences, although their
tef1-α sequences show certain variability corresponding to 99.35–99.78% sequence identity; the
tef1-α sequence of the Asian isolate is unavailable.
Codinaeella lutea resembles
Ca. minuta, but differs in longer setae, narrower conidia, bright yellow color of the conidial masses in vitro and especially colony characteristics in vitro including the color of the pigment released into the agar. The color ranges from yellow to pale ochre, yellow-gold to apricot in
Ca. lutea, but it is usually ochre, deep orange to bright carrot orange in
Ca. minuta. On MLA, the growth of
Ca. lutea is considerably slower (15–18 mm) than that of
Ca. minuta (42–45 mm).
Codinaeella parvilobata also bears a close resemblance to
Ca. lutea, but differs in shorter setae, shorter conidia and colony characteristics. For a detailed comparison of these two species, see notes to
Ca. parvilobata.
Dictyochaeta pakhalensis [
164] resembles
Ca. lutea, but differs in shorter and narrower conidia. In the present phylogeny,
Ca. lutea,
Ca. minuta and
Ca. parvilobata form distinct lineages. Molecular data and culture of
D. pakhalensis are unavailable.
Codinaeella mimusopis (Crous & M.J. Wingf.) Réblová & Hern.-Restr.,
comb. nov. MycoBank MB 842205. (
Figure 22 and
Figure 26).
Basionym. Dictyochaeta mimusopis Crous & M.J. Wingf., Fungal Syst. Evol. 1: 185. 2018.
For description and additional illustrations, see Crous et al. [
82].
Culture characteristics: On CMD: colonies 25–28 mm diam, circular, flat, margin entire, lanose on the inoculation block, mucoid, zonate, whitish becoming pale olivaceous brown, isabelline toward the periphery, reverse of similar colors. On MLA: colonies 38–39 mm diam, circular, flat, raised margin, margin entire, velvety-lanose, partly mucoid, whitish, pale olivaceous brown at the margin, reverse zonate, pale brown, cinnamon at the margin. On OA: colonies 61–63 mm diam, circular, flat, margin entire, lanose becoming mucoid, aerial mycelium restricted to the centre and margin of the colony, brown, olivaceous grey to white-grey centrally and at the margin, reverse olivaceous brown. PCA: colonies 57–58 mm diam, circular, flat, margin entire, lanose at the inoculation block, mucoid and smooth toward the periphery, zonate, whitish centrally, cinnamon, beige toward the margin, reverse brown. Sporulation was moderate on MLA and OA, absent on CMD and PCA.
Specimen examined: SOUTH AFRICA, Eastern Cape Province, Haga Haga, on leaves of Mimusopis caffra, December 2010, M.J. Wingfield (culture ex-type CBS 143435).
Habitat and geographical distribution: On leaves of
Mimusopis caffra known so far from Africa, South Africa [
82]. The GlobalFungi database does not contain similar (≥97%) sequences of this species.
Notes: Our observations based on the ex-type strain match the protologue in all details, except for the conidiophores. Two kinds of conidiophores were observed on sterile stems of
Urtica on CMA. Longer, setiform conidiophores up to 245 μm long were accompanied by shorter conidiophores 38–130 μm vs. conidiophores 40–150 μm [
82]. However, the longer conidiophores were not different from the shorter ones. They have almost the same color, width near the base and wall thickness. They could be considered equivalent to the setae that form regularly in other species of
Codinaeella in nature. The phialides are terminal, elongating percurrently and sympodially to form 1–3 lateral openings. Pigments diffusing from the colony into agar were not produced on any of the used media. Conidia formed on
Urtica stems on CMA accumulated in yellow slimy fascicles (
Figure 26A,B), while those formed on pine needles on SNA and other media were white (not shown).
Codinaeella mimusopis resembles
Ca. lambertiae [
10], but differs in shorter conidiophores and longer conidia [
82].
Codinaeella minuta (Tubaki) Réblová & Hern.-Restr.,
comb. nov. MycoBank MB 842207 (
Figure 27 and
Figure 28).
Basionym. Menispora minuta Tubaki, J. Hattori bot. Lab. 20: 166. 1958.
Description on the natural substrate: Colonies effuse, grey-brown to olivaceous grey, composed of setae and conidiophores, mycelium semi-immersed or immersed. Anamorph. Setae 123–184 μm long, 3–4.5 μm wide near the base, crowded, grow singly or in groups of two, erect, straight or flexuous, septate, smooth, dark brown, paler and thinner-walled toward the apex, unbranched, apex pale brown to subhyaline with a terminal or 1–3 lateral phialidic openings. Conidiophores 32–108 μm long, 3–4 μm wide near the base, macronematous, crowded, arise singly or in groups of 2–3, scattered among the setae, sometimes aggregate in loose columns, unbranched, erect, straight or flexuous, septate, smooth, pale brown, subhyaline toward the apex, sometimes reduced to single conidiogenous cells. Conidiogenous cells 20.5–35 × 2.5–3.5 μm tapering to 1.5–2 μm below the collarette, integrated, terminal, mono- and polyphialidic with 1–5 lateral openings while internally septa can be formed, extending percurrently and sympodially, cylindrical, pale brown, subhyaline toward the apex, usually bearing persistent remnants of the collarettes; collarettes 3–4 μm wide, ca. 2 μm deep, funnel-shaped, subhyaline, the apical part soon evanescent. Conidia 13–18 × 2.5–3.5 μm (mean ± SD = 16.2 ± 1.1 × 2.8 ± 0.3 μm), falcate, tapering toward both ends, slightly truncate at the base with an inconspicuous scar, aseptate, hyaline, with a straight or gently curved setula at each end, 5.5–8.5(–10) μm long, inserted terminally at the apex, subterminally at the base, conidia accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Culture characteristics: On CMD: colonies 61–65 mm diam, circular, flat, margin entire, lanose, floccose, cobwebby toward the margin, becoming mucoid, varying in color due to the absence of aerial mycelium, beige brown to pale banana yellow, occasionally orange with a yellow-gold margin, pale ochre to orange pigment diffusing into agar, reverse beige, ochre-beige to orange. On MLA: colonies 42–45 mm diam, circular, flat, margin entire to fimbriate, lanose, floccose becoming mucoid, grey-brown to dark brown when aerial mycelium is present, deep carrot orange to dull orange brown with orange-gold margin when mucoid, pale orange pigment diffusing into agar, reverse orange to dark amber. On OA: colonies 58–63 mm diam, circular, raised, margin entire, sparsely lanose becoming mucoid, beige-grey to dark grey toward the margin when aerial mycelium is present, orange with tawny margin when mucoid, sometimes orange to ochre pigment diffusing into agar, reverse dark grey-brown or amber. On PCA: colonies 70–73 mm diam, circular, flat, margin entire to fimbriate, sparsely lanose becoming mucoid, grey-brown, tawny or orange with olivaceous brown margin on mucoid spots, pale orange pigment diffusing into agar, reverse dark olivaceous brown to amber. Sporulation was abundant on all media.
Description on CMA with U. dioica stems. Colonies effuse, mycelium composed of branched, septate, hyaline to subhyaline hyphae 1.5–3 μm diam. Anamorph. Setae absent, conidiophores, conidiogenous cells, and conidia similar to those from nature. Conidiophores 46–297(–450) μm long, 3–3.5 μm wide near the base, branched or unbranched, erect, septate, smooth, pale to medium brown. Conidiogenous cells 13.5–32.5 × 3.5–4.5 μm tapering to 1.5–2 μm below the collarette, integrated, terminal, extending percurrently with 1–10 proliferations, and sympodially with 1–4 lateral openings, cylindrical, pale brown to subhyaline, with persistent remnants of the collarettes; collarettes 3.5–5 μm wide, 1.5–2.5 μm deep, funnel-shaped, dark brown becoming hyaline upon aging. Conidia 11.5–17.5(–18) × 2–3 μm (mean ± SD = 14.9 ± 1.1 × 2.5 ± 0.4 μm), falcate, setulae 4.5–7 μm long, accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Specimens examined: CZECH REPUBLIC, Central Bohemian region, Úvaly, Škvorecká obora-Králičina Nature park, on decaying acorn of Quercus sp., 21 April 2018, M. Réblová M.R. 3944 (PRA-20991, culture CBS 146619); Ibid., M.R. 3946 (PRA-20992, culture CBS 146620); Ibid., M.R. 3948 (PRA-20993, culture CBS 146621); Ibid., South Bohemian region, Novohradské hory Mts., Horní Stropnice, Bedřichovský potok Natural Monument, on decaying acorn of Quercus sp., 5 October 2018, M. Réblová M.R. 3983 (PRA-20990). FRANCE, Ariège, Pyrenées Mts., Rimont, La Maille brook, alt. 550 m, on decaying acorn of Quercus sp., 28 May 2018 (incubated for 2 weeks in a damp chamber), J. Fournier M.R. 3951 (PRA-20994, culture CBS 145099); Ibid., M.R. 3952 (PRA-20995, culture CBS 145100). ITALY, on branch of Quercus suber, F. Marras (culture CBS 115959). JAPAN, Chiba Prefecture, Kiyozumi, on dead leaves of Lithocarpus edulis, Aug. 1954, K. Tubaki (authentic strain of Menispora minuta MUCL 9903 = MUCL 15759 = CBS 280.59). THE NETHERLANDS, Gelderland province, Rheden, National Park Veluwezoom, on decaying acorn of Quercus sp., 6 September 1969, V. Holubová-Jechová (CBS 966.69); Ibid., Utrecht province, Utrecht, Utrecht Science park near Westerdijk Fungal Biodiversity Institute, on a fallen leaf of Quercus sp., August 2018, M. Hernández-Restrepo M.H.R. 18063 (culture CBS 147518). USA, Louisiana, New Orleans, Audobon Park, on leaf litter of Quercus virginiana, 19 April 1974, R. Bandoni & K.A. Pirozynski 6120 B/K.A.S. 7210 (culture DAOM 148141). USA, Texas, Houston, E. & A. Taylor Park, on a dead leaf of Quercus virginiana, date unknown, G. Bills (culture TTI-0830). USA, West Virginia, Fayette Co., Fayette Station, hardwood leaf litter, date unknown, collector unknown (culture ATCC 20960 = MF5247 = Mycosearch 1673).
Habitat and geographical distribution:
Codinaeella minuta is a saprobe on fallen leaves and woody fruits of
Fagus sylvatica,
Lithocarpus edulis,
Quercus petraea,
Q. robur,
Q. virginiana,
Quercus sp., and other unknown hosts. It occasionally occurs on bark and wood of
Q. suber. It is known from Asia, Europe, and North America: the Czech Republic, France, Japan, Italy, The Netherlands, Slovak Republic and USA ([
26,
174], this study).
According to GlobalFungi, the identical sequences were found in 20 samples of soil (80%), litter (15%) and root (5%) from forest (95%) or grassland (5%) habitats in Europe (60% of all samples, Denmark, France, Hungary, Poland, Sweden), Asia (15%, Iran), and in western part of North America (25%, Florida, Michigan). The samples originate from eight studies and the locations have temperate or humid continental climate (MAT avg. 13 °C, MAP avg. 864 mm). Codinaeella minuta is identical with Ca. filamentosa in ITS1 barcode. Thus, only ITS2 region was used to study C. minuta biogeography.
Notes: This species was described as
Menispora minuta from dead leaves of
Lithocarpus edulis, an evergreen tree native to Japan [
174]. Although Tubaki [
174] characterized this species in culture only, we update its description based on observations from nature based on the newly acquired material. Among members of
Codinaeella,
Ca. minuta is well distinguished by the production of deep orange pigment on nutrient media, especially on MLA and OA (
Figure 28,
Supplementary Files: Figures S3 and S4).
Codinaeella minuta is similar to
Ca. lutea, but it differs in shorter setae, slightly wider conidia and colony characteristics in vitro. For a detailed comparison, see notes to
Ca. lutea.
Interestingly, majority of the strains of
Ca. minuta included in this study were originally identified as
Codinaea simplex [
2]. It is possible that the concept of the latter species has been misinterpreted in the literature. Our suspicions were confirmed by the revision of the isotype of
C. simplex (DAOM 96020g), which revealed that
Ca. minuta and
C. simplex are two different species.
Codinaea simplex forms a single layer of conidiophores up to 115 μm long (usually 35–68 µm); conidiophores are several-septate but often they are reduced to conidiogenous cells with only 1–2 supporting cells. On the other hand, the conidiophores of
Ca. minuta vary in height and form two layers, setiform conidiophores 123–184 μm long referred to as setae and shorter ones up to 108 μm long. At maturity, setae and conidiophores of
Ca. minuta may cluster in loose columns, which are partly shown on
Figure 27B; they adhere to each other and are difficult to separate. Holubová-Jechová [
26] reported a collection of
C. simplex from the Czech Republic with longer conidiophores 30–155(–230) μm adhering in loose columns similar to synnemata. It is likely that this collection represents in fact
Ca. minuta, which is fairly common in the Czech Republic. Conidiophores of different heights were also observed in culture, in strains growing on stems of
U. dioica on CMA. Solitary, dichotomously branched conidiophores occur frequently in vitro when growing on agar medium. Our observations agree with those of Tubaki [
174]. Using morphological and geographical data from GlobalFungi, the systematic placement of
C. simplex was revaluated and the species was transferred to
Tainosphaeria in this study.
We observed undescribed variability in colony characteristics in strains of
Ca. minuta originating from different geographic regions (
Figures S3 and S4). The presence of orange pigment diffusing into the agar is widespread among strains, and it is most pronounced on MLA and OA. The color of the pigment differs in intensity and ranges from bright carrot orange or deep orange to yellow-orange, pale orange-brown to dark ochre. We observed that colony color is correlated with the development of aerial mycelium. Both isolates from The Netherlands, CBS 966.69 and CBS 147518, abundantly produced deep orange pigment on all media. In these isolates, aerial mycelium was scarce, restricted to the periphery or central zone of the colonies, soon became mucoid and the surface of the colonies turned deep orange (
Figure S3). In other strains the aerial mycelium was more or less abundant, lanose, locally mucoid, and only then the mucoid, bright orange-colored part of the colony with the diffusing pigment was visible. The development of orange pigment and lack of aerial mycelium differed even within a single strain and were found to be correlated with the volume of the nutrient medium and number of colonies on one plate; the pigment production was pronounced when the colonies grew singly on 10 cm Petri dish (
Figure 27F) vs. in a triple on the 6 cm Petri dish (
Figure 28). Although pigment was sparse or absent on all media in the North American strain DAOM 148141 from the frontal view (
Figure 28D), it formed just below the colony, causing the orange coloration of the reverse side (
Figure 27G,H).
In the phylogeny based on ITS-28S-tef1-α sequences, Ca. minuta is represented by 12 strains from Asia, Europe, and North America. While 11 of these strains were grouped with the ex-type strain CBS 298.61 in a monophyletic, well-supported clade, the strain ATCC 20960 clustered at the base of Ca. minuta clade and the relationship was not statistically supported. All 12 strains of Ca. minuta have identical ITS1 and ITS2 sequences, except for the isolate ATCC 20960, which differs in one base pair in the ITS1. More significant sequence divergence was observed in the tef1-α gene. However, the strain ATCC 20960 is well comparable with other strains of Ca. minuta in morphology of conidiophores, conidiogenous cells and conidia in culture, colony characteristics and production of orange pigment. In material collected in different countries the size of conidia of Ca. minuta in vitro was comparable: 13–15.5(–16) × 2–3 μm (Czech Republic), 13–16 × 2–3 μm (France), 10–16 × 2–3 μm (Japan, Tubaki 1958), 11.5–14 × 2–3 μm (USA, DAOM 148141) and 13–16.5(–17) × 2–3 μm (USA, TTI-0830). However, their conidia from nature are slightly larger, 13–18 × 2.5–3.5 μm (Czech Republic, France). In culture, conidia of the strain ATCC 20960 were slightly longer at their upper limit, 13.5–17.5(–18) × 2–3 μm (mean ± SD = 15.4 ± 1.1 × 2.4 ± 0.3 μm) with setulae 4.5–7.5 μm long. Wild type of ATCC 20960 is unknown; the herbarium specimen from which the strain was isolated is not available for study.
Despite the variability of ATCC 20960 in the
tef1-α gene, its morphology and geographic distribution based on ITS analysis of environmental sequences compares well with
Ca. minuta. Therefore, we prefer to keep this strain as a representative of this species. The strain ATCC 20960 is of a particular interest, because it has been studied in detail in terms of bioactive compounds and their potential in disease treatment. It produces β-1,3-
d-glucan synthesis inhibitor with antifungal and anti-
Pneumocystis activity [
175,
176].
Codinaeella parvilobata Réblová & Hern.-Restr.,
sp. nov. MycoBank MB 842208. (
Figure 29 and
Figure 30).
Etymology: Parvus (L) small, lobus (L) lobe, referring to small lobes at the base of conidiophores in vitro.
Typification: CZECH REPUBLIC, South Bohemian region, Novohradské hory Mts., Dobrá voda, Hojná voda National nature monument, on decaying cupule of Fagus sylvatica, 29 September 2017, M. Réblová M.R. 3900 (PRA-20987 holotype, culture ex-type CBS 144536).
Description on the natural substrate: Colonies effuse, brown-grey, composed of setae and conidiophores, mycelium semi-immersed. Anamorph. Setae 140–205 μm long, 4.5–5.5 μm wide near the swollen base, crowded, arise singly or in groups, erect, straight, flexuous or bent, septate, smooth, dark brown and thick-walled, paler and thinner-walled toward the apex, simple, apex pale brown with a terminal or 1–6 lateral phialidic openings, sometimes apex remains sterile, obtuse and hyaline. Conidiophores 40–117 μm long, 3–4.5(–5) μm wide near the base, macronematous, crowded, arise singly or in groups, scattered among the setae, unbranched, erect, straight, usually flexuous or bent, geniculate to undulate toward the apex, septate, smooth, mid to pale brown, paler toward the tip. Conidiogenous cells 9–34.5 μm × 3.5–4.5 μm tapering to 1–2 μm below the collarette, integrated, terminal, monophialidic, or polyphialidic with 1–7 lateral openings while internally septa can be formed, extending percurrently and sympodially, cylindrical, pale brown, subhyaline at the apex, usually with persistent remnants of the collarettes; collarette 2–2.5 μm wide, 1.5(–2) μm deep, funnel-shaped, hyaline, the apical part soon evanescent. Conidia 10.5–13.5 μm × 2–2.5(–3) μm (mean ± SD = 12.4 ± 0.8 μm × 2.0 ± 0.2 μm), falcate, tapering toward both ends, slightly truncate at the base with an inconspicuous scar, aseptate, hyaline, with a straight or gently curved setula at each end, 6.5–9 μm long, inserted terminally at the apex, subterminally at the base, conidia accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Culture characteristics: On CMD: colonies 40–43 mm diam, circular, flat, convex centrally, margin fimbriate, velvety-lanose, mucoid toward the periphery, whitish, beige to beige-grey, ochre-beige to brown when mucoid, isabelline toward the margin, reverse isabelline. On MLA: colonies 32–36 mm diam, circular, flat, convex centrally, margin fimbriate, lanose becoming mucoid, wrinkled, whitish grey, buff yellow, cinnamon to olivaceous brown when mucoid, pale yellow to pale ochre pigment diffusing into agar, sometimes the pigment is missing, reverse dark ivory to ochre, dark brown margin. On OA: colonies 78–80 mm diam, circular, flat, convex centrally, margin entire, velvety-lanose becoming mucoid, zonate, smoke grey, cinnamon to dark olivaceous brown when mucoid, aerial hyphae with colorless exudates, reverse amber to dark brown. On PCA: colonies 46–50 mm diam, circular, flat, convex centrally, margin entire to fimbriate, velvety-lanose, somewhat funiculose at the centre, mucoid to cobwebby toward the periphery, pale olivaceous beige, reverse beige to dark grey. Sporulation was moderate on MLA and PCA, absent on CMD and OA.
Colonies on MLA effuse, mycelium composed of branched, septate, subhyaline to pale brown hyphae 1.5–3.5 μm diam, or monilioid hyphae 4–5.5 μm diam. Anamorph. Setae absent, conidiophores, conidiogenous cells and conidia similar to those from nature. Conidiophores 38–84 μm long, 1.5–3 μm wide near the base, sometimes tapering toward the base, arise singly from vegetative or monilioid hyphae, usually attached by small lobes at the base, unbranched, erect, septate, smooth, pale brown. Conidiogenous cells 20–31 × 2.5–3.5 μm tapering to 1.5–2 μm, monophialidic, pale brown; collarette 2–2.5 μm wide, 1.5–2 μm deep, funnel-shaped, pale brown. Conidia 10.5–14 × 2–2.5 μm (mean ± SD = 12.6 ± 0.8 × 2.3 ± 0.2 μm), falcate, hyaline, aseptate, setulae 4–8 μm long, accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Other specimens examined: CZECH REPUBLIC, South Bohemian region, Novohradské hory Mts., Dobrá voda, Hojná voda National nature monument, on decaying seed of Fagus sylvatica, 4 October 2012, M. Réblová M.R. 3757 (PRA-20988, culture CBS 144658 = MUCL 56490). CZECH REPUBLIC, Central Bohemian region, Úvaly, Škvorecká obora-Králičina Nature park, on decaying acorn of Quercus sp., 21 April 2018, M. Réblová M.R. 3947 (PRA-20989, culture CBS 144792). BELGIUM, Wallonie, Brabant wallon, Louvain-la-Neuve, Bois de Lauzelle, on decaying leaf in forest stream, date unknown, date unknown, G.L. Hennebert (culture MUCL 28054).
Habitat and geographical distribution: A saprobe on decaying seeds and wooden fruits often buried in soil and on leaves of Fagus sylvatica, Quercus sp. and an unknown host in terrestrial and freshwater biotopes, known from Europe from Belgium and the Czech Republic.
According to GlobalFungi, the identical sequence was found in 227 samples of soil (65%), litter (13%), root (11%), and deadwood (9%) collected mostly in forest (90%), grassland (4%) or anthropogenic (4%) habitats. Most of the samples were from Europe (177 samples, Czech Republic, Belgium, Denmark, France, Germany, Wales), 25 samples were from North America (Massachusetts, New Jersey) and 25 samples were from Asia (Eastern China, Japan and South Korea). The samples originated from 25 studies. The sites have temperate climate (MAT avg. 8.4 °C, MAP avg. 851 mm).
Notes: In culture, some conidiophores of Ca. parvilobata are attached to the pigmented, monilioid hyphae with a lobate, swollen base, a character that was not observed in other species of the genus. The pale yellow to pale ochre pigment diffusing in the agar on MLA was produced only in two strains, including the ex-type strain. The pigment production and visibility is correlated with the absence of aerial mycelium; the colonies become mucoid and their surface changes from whitish-grey to buff yellow, cinnamon to olivaceous brown.
It is challenging to distinguish Ca. parvilobata and Ca. lutea based on morphological characters, because the size of their conidiophores, conidiogenous cells, and also conidia partially overlap under natural conditions. Codinaeella lutea differs in having longer setae and longer conidia in their upper range. However, both species differ especially in colony characteristics and pigment production on MLA and OA. Codinaeella parvilobata can also be compared with Ca. minuta, but the latter species differs in longer conidia and production of ochre to deep orange pigment.
Codinaeella pini (Crous & M.J. Wingf.) Réblová & Hern.-Restr.,
comb. nov. MycoBank MB 842210 (
Figure 22 and
Figure 31).
Basionym. Codinaea pini Crous & M.J. Wingf., Persoonia 33: 251. 2014.
For description and additional illustrations, see Crous et al. [
8].
Culture characteristics: On CMD: colonies 60–61 mm diam, circular, raised, margin entire, sparsely lanose, floccose becoming mucoid, beige to olivaceous grey centrally, isabelline-beige toward the periphery, reverse brown. On MLA: colonies 76–77 mm diam, circular, raised, margin entire, lanose, floccose, mucoid at the margin, finely furrowed, whitish, ochre-beige at the margin, pale yellow-ochre pigment diffusing into the agar, reverse brown. On OA: colonies 76–78 mm diam, circular, flat, margin entire, cobwebby to powdery centrally becoming mucoid, beige, whitish toward the margin, reverse pale brown. On PCA: colonies 78–79 mm diam, circular, flat, margin entire, sparsely lanose becoming mucoid, pale brown centrally, isabelline toward the periphery, reverse whitish. Sporulation was abundant on PCA, moderate on CMD, MLA, and OA.
Specimen examined: UGANDA, on dead needles of Pinus patula, January 2014, M.J. Wingfield (culture ex-type CBS 138866).
Habitat and geographical distribution: Saprobe on dead needles of
Pinus patula, known so far from Africa, Uganda [
8]. According to GlobalFungi, the identical sequences were found in 58 soil and five shoot samples (
Pinus taeda needles) in the forests of North America (North Carolina, Louisiana, Ontario, British Columbia) originating from studies of Wilhelm et al. [
177] and Oono et al. [
178]. The sites have temperate, hemiboreal, or humid continental climate (MAT avg. 17 °C, MAP avg. 1144 mm).
Notes: Our observations of the ex-type strain match the protologue. However, conidiophores on MLA formed two distinct layers compared to a single layer of conidiophores 30–100 μm long reported by Crous et al. [
8]. On MLA, the upper layer consisted of unbranched, flexuous or bent, setiform conidiophores 130–232 × 2.5–3.5 μm terminating into a mono- or polyphialide with 1–2 lateral apertures, while the lower layer comprised of shorter conidiophores 56–98 × 2.5–3.5 μm with a terminal phialidic aperture. The strain sporulated on all nutrient media included in this study and also on stems of
Urtica on CMA and pine needles on SNA. Conidia accumulated in slimy whitish fascicles.
Codinaeella yunnanensis (Z.L. Luo, K.D. Hyde & H.Y. Su) Réblová & Hern.-Restr., comb. nov. MycoBank MB 842211.
Basionym. Codinaea yunnanensis Z.L. Luo, K.D. Hyde & H.Y. Su, Fungal Divers. 99: 590. 2019.
For description and illustrations, see Luo et al. [
29].
Habitat and geographical distribution: Saprobe on submerged decaying wood, known only from Asia, China [
29]. According to GlobalFungi, the identical sequences were found in three soil samples from forests in Asia (Central China Hubei province and Iran) [
115,
179]. The localities have temperate climate (MAT avg. 16 °C, MAP avg. 1094 mm).
Notes:
Codinaeella yunnanensis resembles
Ca. lutea and
Ca. mimusopis in conidial characters, however, the size of their conidia significantly overlaps. For morphological comparison, see
Table S5. In the phylogenetic analysis, all three species formed separate lineages;
Ca. yunnanensis nested in a clade with
Ca. lutea and
Ca. pini.
3.5.3. The Genus Nimesporella
Nimesporella Réblová & Hern.-Restr., gen. nov. MycoBank MB 842002.
Type species. Nimesporella capillacea Réblová & Hern.-Restr.
Etymology: An anagram of Menisporella, a synonym of Codinaea.
Description: Colonies effuse, brown, composed of conidiophores, mycelium immersed. Anamorph. Setae absent. Conidiophores macronematous, mononematous, single or in groups, erect, straight or flexuous to geniculate in the upper part, unbranched, septate, smooth, brown, paler toward the apex. Conidiogenous cells integrated, terminal, polyphialidic, extending sympodially, paler than the conidiophore, often with persistent remnants of the collarettes; collarettes flared, soon evanescent. Conidia ellipsoidal, rounded at the apical end, papillate at the basal end, aseptate, hyaline, with a straight or gently curved setula at each end, basal setulae positioned ventrally, conidia accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Habitat and geographical distribution: Nimesporella occurs on decaying plant material in freshwater and terrestrial habitats, and it is known from Africa.
Notes:
Nimesporella capillacea is the only species of the genus available to represent this uncommon morphotype in the present phylogeny. It forms a basal lineage to the
Codinaea clade but their relationship is not statistically well supported.
Nimesporella differs from
Codinaea mainly by the characters of conidiogenous cells with multiple phialidic apertures, early evanescent collarettes and papillate, broadly ellipsoidal conidia with a simple setula at each end, inserted terminally at the apex and somewhat ventrally at the base. When grown in culture, the phialides extend sympodially for a short distance, producing densely aggregated phialidic openings encircled by tubular, apically flared collarettes. Similar polyphialides were also recorded in
Multiguttulispora [
49].
Species such as
Dictyochaeta aliformis,
D. daphnioides and
D. tumidospora known only from Southeast Asia from Malaysia [
22] bear a striking resemblance to
N. capillacea in all characteristics. On material from nature [
22], conidiogenous cells frequently elongate sympodially, while internally septa may be formed. As a result, 1–2 intercalary phialidic openings are formed per each cell in the upper part of the conidiophore. After careful comparison of their original descriptions and illustrations with
N. capillacea we suggest that they are likely congeneric. Their transfer to
Nimesporella should be supported by DNA data, which are unavailable for study at the moment. The strain of
N. capillacea was originally deposited under the name
D. daphnioides.
Nimesporella capillacea Réblová & Hern.-Restr.,
sp. nov. MycoBank MB 842212. (
Figure 32).
Etymology: Capillus (L) hair or thread, -acea (L) indicating resemblance, meaning hair-like or thread-like, referring to conidial setulae.
Typification: IVORY COAST, Tai National Park, leaf litter, 11 March 1992, S. Pagano (holotype IMI 358908 microscopic slides, ex-type culture IMI 358908).
Culture characteristics: On CMD: colonies 41–42 mm diam, circular, flat, margin fimbriate, sparsely lanose becoming cobwebby, white, beige toward the periphery, reverse grey-beige, isabelline at the margin. On MLA: colonies 38–39 mm diam, circular, convex, margin entire, lanose, floccose, furrowed, cobwebby at the margin, whitish centrally, beige-grey to ochre-beige with an olivaceous beige outer zone, reverse dark amber. On OA: colonies 65–68 mm diam, circular, flat, margin entire to weakly fimbriate, velvety-lanose, floccose to funiculose centrally, mucoid at the margin, beige with an olivaceous outer zone, reverse olivaceous beige. On PCA: colonies 51–52 mm diam, flat, circular, flat, similar to colonies on OA, sparsely lanose, floccose to funiculose centrally, whitish-beige becoming olivaceous beige, with an isabelline outer zone of submerged growth, reverse zonate, olivaceous beige. Sporulation was abundant on MLA, OA, PCA, sparse on CMD.
Colonies on MLA effuse, mycelium composed of branched, septate, hyaline hyphae 1.5–2.5 μm diam. Anamorph. Setae absent. Conidiophores 28–130 μm long, 3–4.5 μm wide near the base, macronematous, occasionally reduced to conidiogenous cells, grow solitarily from vegetative hyphae or arise in groups of 2–3 from dark knots of hyphal cells, erect, straight or slightly bent, septate, smooth, brown, paler toward the tip. Conidiogenous cells 22–35(–45) × 3–3.5 μm, slightly tapering upwards, 2–3.5 μm wide below the fertile apical part, integrated, terminal, extending sympodially over a short distance, polyphialidic with 5–22 lateral openings, fertile part 5.5–17(–23.5) × 2–3.5 μm, occasionally dichotomously branched, with persistent remnants of the collarettes; collarettes 1.5–2 μm wide, 1.5–2 μm deep, hyaline, minute, tubular, apically flared, evanescent, the apical part sometimes attached to the bottom of the conidium. Conidia 8–11.5 μm × 4.5–5 μm (mean ± SD = 9.5 ± 0.9 × 4.5 ± 0.2 μm), ellipsoidal, broadly rounded at the apical end, papillate or beak-like at the basal end, aseptate, hyaline, with a straight or gently curved setula at each end, 2–3.5 μm long, positioned terminally at the apex and subterminally at the base, conidia accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Habitat and geographical distribution: Saprobe on leaf litter in freshwater habitat, known in Ivory Coast.
Notes:
Nimesporella capillacea closely resembles
N. aliformis,
N. daphnioides and
N. tumidospora, all known only from Malaysia [
22], but which differ in having longer conidiophores and larger conidia.
3.5.4. The Genus Stilbochaeta
Stilbochaeta Réblová & Hern.-Restr., gen. nov. MycoBank MB 842001.
Etymology: Stilbos (Greek) glistening, referring to slimy droplets of conidia that are glistening when fresh; chaeta (Greek) hair, bristle.
Type species. Stilbochaeta malaysiana (Kuthub.) Réblová & Hern.-Restr.
Description on the natural substrate: Colonies effuse, hairy, brown, composed of setae and conidiophores, occasionally ascomata, mycelium immersed or semi-immersed. Anamorph. Setae grow singly or in groups from stromatic cells or knots of hyphal cells, erect, straight or flexuous, septate, brown, paler toward the apex, unbranched, apex sterile, bluntly rounded, sometimes with a terminal or several lateral phialidic openings. Conidiophores macronematous, mononematous, single or grow in fascicles from stromatic cells or knots of hyphal cells around the base of the setae, erect, straight or flexuous to slightly undulating, unbranched, septate, smooth, brown, paler toward the apex. Conidiogenous cells integrated, terminal, mono- or polyphialidic, extending percurrently and sympodially, paler than the conidiophores; collarettes flared, funnel-shaped, often slightly stipitate or tubular near the base. Conidia falcate, oblong-falcate, ellipsoidal-fusiform, curved, tapering toward both ends, slightly truncate at the base, with an inconspicuous basal scar, 0–1(–3)-septate, hyaline, with a straight or gently curved setula at each end, setulae simple, bifid or trifid, inserted terminally at the apex and subterminally at the base, conidia accumulate in slimy fascicles. Teleomorph. Ascomata perithecial, non-stromatic, superficial, globose to subglobose, papillate, dark brown, setose; setae sterile, rounded at the apex. Ostiole periphysate. Ascomatal wall fragile, carbonaceous, two-layered. Paraphyses disappearing with age, septate, tapering, longer than the asci. Asci unitunicate, cylindrical-clavate, shortly-stipitate, apically rounded, ascal apex with a non-amyloid apical annulus. Ascospores fusiform, hyaline, transversely septate, without gelatinous sheath or appendages.
Habitat and geographical distribution: Members of
Stilbochaeta occur in freshwater and terrestrial habitats, they are saprobes on leaf litter, bamboo and decaying bark and wood. They are widespread throughout the world, especially in tropical and temperate regions of Africa, Asia, Australasia, the Caribbean and South America ([
2,
17,
19,
28,
29,
31,
81,
180], this study).
Notes: Stilbochaeta is introduced for a group of former Codinaea and Dictyochaeta species. It is morphologically and phylogenetically well-delimited and characterized by arrangement of setae and conidiophores in bundles growing from dark stromatic cells or knots of hyphal cells. Setae have sterile, bluntly rounded apices that are sometimes modified into mono- or polyphialides, while in Codinaea the setae are tapering toward the apices, which may be also fertile. The conidia are always septate, although aseptate and septate conidia can sometimes occur together in one strain, in contrast to morphologically similar Codinaea with exclusively aseptate conidia and setae. In culture, representatives of Stilbochaeta often form yellow, gold, cinnamon, or amber pigments that diffuse into the agar. The aerial mycelium that forms in vitro is often pigmented and the surface of the colonies is usually olivaceous cinnamon, olivaceous gold or beige-orange.
Eight species are assigned to the genus, all of which were verified using molecular data. Three other species closely resemble
Stilbochaeta but have setae with acute, rarely obtuse apices and include
Dictyochaeta tortuosa [
7] with 1–2-septate conidia, and
D. caatingae [
121] and
D. matsushimae [
27] with 2–3-septate conidia. These species have not yet been evaluated based on molecular data. A synopsis table with diagnostic features of accepted species of
Stilbochaeta is provided as a
Supplementary File: Table S6.
Stilbochaeta aquatica (W. Dong & H. Zhang) Réblová & Hern.-Restr.,
comb. nov. MycoBank MB 842213. (
Figure 33 and
Figure 34).
Basionym. Dictyochaeta aquatica W. Dong & H. Zhang, Phytotaxa 362: 193. 2018.
Culture characteristics: On CMD: colonies 28–32 mm diam, circular, flat, margin entire, velvety, somewhat floccose, mucoid at the margin, whitish with a pale apricot tinge, yellow toward the margin, with an outer zone of submerged growth, pale yellow pigment diffusing into the agar, reverse pale apricot. On MLA: colonies 49–50 mm diam, circular, flat, slightly raised centrally, velvety becoming cobwebby, mucoid at the margin, whitish with an apricot-gold tinge centrally, whitish-yellow toward the periphery, pale gold pigment diffusing into the agar, reverse gold to honey. On OA: colonies 45–48 mm diam, circular, convex centrally, flat margin, margin entire, velvety, cobwebby to mucoid at the centre, zonate, yellow centrally, white toward the margin with yellow-white outer zone, pale yellow pigment diffusing into the agar, reverse bright yellow. On PCA: colonies 52–56 mm diam, circular, flat, margin entire, mucoid, yellow-gold centrally, isabelline toward the margin, yellow pigment diffusing into the agar at the centre of the colony, reverse of the same colors. Sporulation was absent on all media.
Colonies on CMA with U. dioica stems effuse, hairy, mycelium composed of branched, septate, hyaline to subhyaline hyphae 1.5–2 μm diam. Anamorph. Setae 122–182 μm, 3.5–4.5 μm wide above the base, arise singly or in groups of two from knots of dark brown hyphal cells, erect, straight or flexuous, septate, smooth, dark brown and thick-walled, paler and thinner-walled toward the apex, unbranched, apex pale brown, with a terminal or 1–3 lateral phialidic openings. Conidiophores 45–102 μm long, 3–4 μm wide near the base, macronematous, single or arise in fascicles of 2–4 from knots of dark hyphal cells around the base of the setae, erect, straight or flexuous, septate, smooth, medium to pale brown, paler toward the tip. Conidiogenous cells 10–29 × 3–3.5 μm tapering to 1.5–2 μm below the collarette, integrated, terminal, mono-, or polyphialidic with 1–4 lateral openings, extending sympodially, occasionally percurrently, subcylindrical, pale brown, subhyaline at the apex, sometimes with persistent remnants of the collarettes; collarettes 2.5–4.5 μm wide, 2–3.5 μm deep, funnel-shaped to slightly tubular, subhyaline. Conidia 13.5–17.5 × 2.5–3 μm (mean ± SD = 16.0 ± 1.1 × 2.7 ± 0.6 μm), falcate to oblong-falcate, with an inconspicuous basal scar, 0–1-septate, hyaline, with a straight or gently curved setula at each end, 5–8.5 μm long, positioned terminally at the apex and slightly subterminally at the base, conidia accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Specimen examined: PHILIPPINES, Negros Occidental, Bario Alegria, Lupit River, submerged bamboo stems, 18 August 2001, L. Cai (CBS 114070 ex-type strain of Dictyochaeta curvispora, contamination).
Habitat and geographical distribution: Saprobe on submerged wood and bamboo culm, known from Southeast Asia from the Philippines and Thailand ([
28], this study). According to GlobalFungi, the identical sequences were found in one soil sample in forest in Asia (South China) [
162]. The location has a temperate climate (MAT avg. 22 °C, MAP avg. 1627 mm).
Notes: Cai et al. [
181] described
D. curvispora for a hyphomycete with pigmented, unbranched conidiophores terminating into a monophialide and producing aseptate, hyaline, cylindrical, crescent-shaped to falcate conidia without setulae. According to the authors, the ex-type strain CBS 114070 did not sporulate in culture. It formed greyish-yellow colonies on potato-dextrose agar, a yellow pigment staining agar around the colony and pale orange-brown reverse. These culture characteristics, in particular the presence of the yellow pigment diffusing into the agar, correspond with our observations. However, the morphology of
D. curvispora is not congruent with the placement of its ex-type strain among
Stilbochaeta species. We therefore suspected that the ex-type strain might have been contaminated during isolation. Cultivation of the strain CBS 114070 on
Urtica stems on CMA yielded sporulating colonies of a fungus with the
Stilbochaeta morphology and provided the evidence that the ex-type strain of
D. curvispora was indeed contaminated. Based on comparison with morphologically similar species and their DNA data, this isolate was identified as
S. aquatica. Based on their protologues and original illustrations,
D. curvispora and
S. aquatica are two different species. For reasons explained above, the strain CBS 114070 cannot be used to represent
D. curvispora; the systematic placement of this species remains unknown.
Stilbochaeta brevisetula (S. Hughes & W.B. Kendr.) Réblová & Hern.-Restr.,
comb. nov. MycoBank MB 842214. (
Figure 35,
Figure 36 and
Figure 37).
Basionym. Codinaea brevisetula S. Hughes & W.B. Kendr., N. Z. J. Bot. 6: 338. 1968.
≡ Dictyochaeta brevisetula (S. Hughes & W.B. Kendr.) Aramb. & Cabello, Mycotaxon 34: 681. 1989. (Nom. inval., Art. 41.4).
≡ Dictyochaeta brevisetula (S. Hughes & W.B. Kendr.) Whitton, McKenzie & K.D. Hyde, Fungal Divers. 4: 137. 2000.
Typification: NEW ZEALAND, Wellington region, Tongariro National Park, Ohakune Mountain Road (alt. 762 m), on bark of Nothofagus fusca, 7 March 1963, S.J. Hughes 439a (holotype of Codinaea brevisetula PDD 20652, isotype DAOM 96420a). NEW ZEALAND, Otago region, Queenstown-Lakes district, Mount Aspiring National Park, Makarora, Mt. Shrimpton track, on decaying wood of a trunk of Nothofagus sp., 24 March 2005, M. Réblová M.R. 3347/NZ 620 (epitype designated here MBT 10004617, PDD 119677, culture ex-epitype ICMP 22549).
Description on the natural substrate: Colonies effuse, grey-brown to olivaceous grey, composed of setae, conidiophores and ascomata. Anamorph. Setae 94–150 μm long, 3.5– μm wide above the base, single or arise in groups of 2–3 from knots of dark brown hyphal cells, erect, straight or slightly flexuous, septate, smooth, medium brown, thick-walled, darker at the base, paler and thinner walled toward the apex, unbranched, occasionally extending percurrently, apex subhyaline, bluntly rounded and slightly inflated, sterile. Conidiophores 43–100 μm long, 3.5–4.5 μm wide above the base, macronematous, arise single or in fascicles of 4–9 from knots of dark brown hyphal cells around the base of the setae, erect, straight to slightly undulating, septate, smooth, pale brown, paler toward the tip. Conidiogenous cells 16.5–32 × 3.5–4.5 μm tapering to (1.5–)2 μm below the collarette, integrated, terminal, mono- or polyphialidic with 1–3 lateral openings, extending percurrently and sympodially, cylindrical, pale brown, subhyaline at the apex; collarettes 3.5–4 μm wide, 2–2.5(–3) μm deep, funnel-shaped, subhyaline. Conidia 17–22 × 3–4 μm (mean ± SD = 19.2 ± 1.6 × 3.3 ± 0.3 μm), oblong-falcate, slightly tapering toward both ends, with an inconspicuous basal scar, 1-septate, hyaline, with a straight or gently curved setula at each end, 1–1.5 μm long, inserted terminally at the apex and subterminally at the base, conidia accumulate in slimy whitish fascicles. Teleomorph. Ascomata 250–330 μm diam, non-stromatic, superficial, globose to subglobose, papillate, dark brown, clothed with setae that are similar to those accompanying the conidiophores of the anamorph, becoming glabrous with age; setae yellow-brown, pale brown to subhyaline toward the apex, apex bluntly rounded, slightly inflated, sterile. Ostiole periphysate. Ascomatal wall fragile, carbonaceous, 24–30 μm thick, two-layered. Outer layer consisting of brown, polyhedral cells with opaque walls. Inner layer consisting of several rows of thin-walled, elongated, hyaline cells. Paraphyses 3.5–5 μm wide tapering to 1.5–3 μm, septate, longer than the asci. Asci 100–114 × 7.5–8.5 μm (mean ± SD = 108.0 ± 4.9 × 8.0 ± 0.5 µm), 83–98 μm (mean ± SD = 92.0 ± 4.4 µm) long in the sporiferous part, cylindrical-clavate, stipitate, apically rounded, ascal apex with a shallow, non-amyloid apical annulus 2.5–3 μm wide, 1–1.5 μm high. Ascospores (15–)16–24 × 3–4(–4.5) μm (mean ± SD = 18.7 ± 2.1 × 3.4 ± 0.3 µm), fusiform, hyaline, 3-septate, 2-seriate or obliquely uniseriate in the ascus.
Culture characteristics: On CMD: colonies 32–35 mm diam, circular, flat, raised centrally, margin entire, velvety-lanose, mucoid locally, cobwebby at the margin, with a prominent outer zone of submerged growth, finely furrowed, white to pale ochre-beige, isabelline at the margin, reverse pale olivaceous beige. On MLA: colonies 33–36 mm diam, circular, flat, convex centrally, margin fimbriate, lanose, olivaceous ochre to olivaceous cinnamon, with an olivaceous brown to apricot outer zone of submerged growth, pale ochre-brown pigment diffusing into agar, reverse dark olivaceous orange. On OA: colonies 32–34 mm diam, circular, convex, margin entire to undulate, lanose, somewhat floccose, aerial mycelium occasionally reduced, colonies cobwebby to mucoid, pale olivaceous apricot to olivaceous grey-brown with an orange tinge, pale ochre/apricot pigment diffusing into agar, reverse olivaceous-ochre to cinnamon, isabelline at the margin. On PCA: colonies 30–32 mm diam, circular, flat, raised centrally, margin undulate, lanose, cobwebby toward the periphery, white-beige at the centre, pale beige-apricot to pale sepia toward the periphery with an outer zone of submerged growth, pale apricot pigment diffusing into agar, reverse olivaceous apricot. Sporulation was abundant on MLA, OA, moderate on PCA, absent on CMD.
Colonies on OA effuse, hairy, mycelium composed of branched, septate, subhyaline to hyaline hyphae 2–3 μm diam. Anamorph. Setae, conidiophores, conidiogenous cells and conidia similar to those from nature. Setae 116–132 μm long, 4–5 μm wide above the base, extending percurrently, sterile. Conidiophores 49–87(–105) μm long, 3.5–4.5 μm wide above the base, arise singly or in small groups, interspersed among the setae. Conidiogenous cells 14–34 × 3.5–5 μm tapering to 2–2.5 μm below the collarette, integrated, terminal, mono- or polyphialidic with 1–2 lateral openings, extending percurrently and sympodially, cylindrical, pale brown, subhyaline at the apex; collarettes 4–4.5 μm wide, 2–2.5 μm deep, funnel-shaped, subhyaline. Conidia (15.5–)17–22 × 3–4 μm (mean ± SD = 19.2 ± 1.6 × 3.4 ± 0.3 μm), 1-septate, setulae 1–1.5 μm long, accumulate in slimy whitish fascicles. Teleomorph. Not observed.
Specimens examined: NEW ZEALAND, Otago region, Queenstown-Lakes district, Mount Aspiring National Park, Makarora, Mt. Shrimpton track, on decaying wood of Nothofagus sp., 24 March 2005, M. Réblová M.R. 3339/NZ 611 (PDD 119678, ICMP 22548); Ibid., M.R. 3338/NZ 610, M.R. 3344/NZ 616, M.R. 3349/NZ 622. NEW ZEALAND, Otago region, Queenstown-Lakes District, Makarora, Makarora Bush Walk, on decaying wood of a trunk, 30 March 2005, M. Réblová M.R. 3410/NZ 691 (PDD 119679, ICMP 22551). NEW ZEALAND, West Coast region, Buller district, Victoria Forest Park, Lyell, Lyell Historical Walk, on decaying wood of a branch, 6 April 2005, M. Réblová M.R. 3455/NZ 742 (PDD 119680, ICMP 22552). NEW ZEALAND, West Coast region, Buller district, Victoria Forest Park, Murray Creek Track, on decaying wood of Nothofagus sp., 21 February 2003, M. Réblová & K.A. Seifert M.R. 2587/NZ 49 (PDD 81887). NEW ZEALAND, West Coast region, Buller district, Victoria Forest Park, Palmer’s Hut ca. 18 km SW of Springs Junction on unpaved road, Lake Christabel track, on decaying wood of Nothofagus sp., 28 February 2003, M. Réblová M.R. 3423/NZ 162 (PDD 119681, ICMP 15125). NEW ZEALAND, West Coast region, Westland district, Otira, Kelly Shelter, Cockayane Nature Walk MR, on decaying wood, 11 April 2005, M. Réblová M.R. 3466/NZ 754.
Habitat and geographical distribution: Saprobe on decaying bark and wood of
Nothofagus fusca,
Nothofagus sp. and other unidentified hosts, known only from Australasia from New Zealand ([
2], this study). The database GlobalFungi does not contain similar sequences (≥98%) of this species.
Notes: Stilbochaeta brevisetula is the only species of the genus known to have a teleomorph, which is reported for the first time in this study. Ascomata are often covered by setae that resemble those associated with the conidiophores of the anamorph, although with age the setae disappear and ascomata become glabrous. The anamorph-teleomorph relationship was experimentally verified. Stilbochaeta brevisetula is well distinguishable from other members of the genus by the shortest conidial setulae (1–2.7 μm) and setae that are sterile, apically bluntly rounded and slightly inflated.
The teleomorph of
S. brevisetula is remarkably similar to
Chaetosphaeria (Ch.)
hebetiseta [
182], a saprobic lignicolous species with a
Chloridium-like anamorph. Their ascomata are covered with always sterile, brown, septate setae that terminate into rounded subhyaline cells and they also share 3-septate, hyaline, fusiform ascospores of comparable size. However, the ascospores of
S. brevisetula are smooth-walled, while those of
Ch. hebetiseta are verrucose, also their anamorphs clearly differ in characters of conidia. The two species are shown distantly related in the phylogenetic tree (
Figure 2).
Stilbochaeta cangshanensis (Z.L. Luo, K.D. Hyde & H.Y. Su) Réblová & Hern.-Restr., comb. nov. MycoBank MB 842215.
Basionym. Dictyochaeta cangshanensis Z.L. Luo, K.D. Hyde & H.Y. Su, Fungal Divers. 99: 592. 2019.
For description and illustrations, see Luo et al. [
29].
Habitat and geographical distribution: Saprobe on submerged decaying wood, known only from Asia, China [
29]. According to GlobalFungi, the identical sequences were found in 13 soil and one shoot samples collected in forests of Europe (five samples, South Italy), Asia (seven samples, Iran, South China), and Eastern North America (one sample, Tennessee) [
115,
117,
162,
183,
184]. The sites have temperate or Mediterranean climate (MAT avg. 17 °C, MAP avg. 842 mm).
Notes: This species closely resembles
S. aquatica and it is difficult to distinguish them based solely on morphological characters, especially when the size of their conidia significantly overlap, i.e., 14–18 × 2–3 μm with setulae 8–13 μm long in
S. aquatica [
28] vs. 15–18 × 2.5–3.5 μm in
S. cangshanensis [
29]; the length of the setulae of the latter species were not given in the protologue. In the phylogenetic analysis, the two species form distinct lineages.
Stilbochaeta malaysiana (Kuthub.) Réblová & Hern.-Restr.,
comb. nov. MycoBank MB 842216. (
Figure 34 and
Figure 38).
Basionym. Dictyochaeta malaysiana Kuthub., Trans. Br. mycol. Soc. 89: 356. 1987.
Culture characteristics: On CMD: colonies 24–25 mm diam, circular, raised, margin undulate, lanose, cobwebby at the margin, grey-beige centrally, beige toward the margin, ochre pigment diffusing into agar, reverse dark cinnamon. On MLA: colonies 62–63 mm diam, circular, raised, flat margin, margin fimbriate, lanose, zonate, finely furrowed, whitish-beige becoming olivaceous ochre-beige toward the periphery, dark cinnamon at the margin, caramel pigment diffusing into the agar, reverse dark cinnamon. On OA: colonies 61–62 mm diam, circular, raised, margin lobate, lanose, floccose, furrowed, zonate with a central cobwebby ring surrounded by small lanose areas of radially spreading mycelium, ochre-grey centrally, pale ochre-beige to olivaceous cinnamon toward the periphery, aerial hyphae with numerous pale ochre exudates, dark cinnamon pigment diffusing into the agar, reverse dark amber to brown. On PCA: colonies 32–38 mm diam, flat, circular, margin undulate, sparsely lanose to cobwebby, ochre-beige, pale cinnamon pigment diffusing into the agar, reverse cinnamon. Sporulation was moderate on CMD and PCA, absent on MLA and OA.
Colonies on CMA with U. dioica stems effuse, hairy, mycelium composed of branched, septate, hyaline to subhyaline hyphae 1.5–3 μm diam. Anamorph. Setae 230–281 μm long, 4–5.5 μm wide near the swollen base, single or arise in groups of two from knots of dark brown hyphal cells, erect, straight or flexuous, septate, smooth, dark brown and thick-walled, paler and thinner-walled toward the apex, unbranched, apex pale brown, sterile. Conidiophores 78–113 μm long, 4–5.5 μm wide near the base, macronematous, single or arise in fascicles of 2–5 from knots of dark brown hyphal cells around the base of the setae, erect, straight or bent, septate, smooth, medium to pale brown, paler toward the tip. Conidiogenous cells 17–21(–37) × 3.5–4.5(–5) μm tapering to 1.5–2 μm below the collarette, integrated, terminal, mono- or polyphialidic with 1–3 lateral openings while internally septa can be formed, extending percurrently and sympodially, subcylindrical, pale brown; collarettes 4.5–7 μm wide, 3.5–4.5 μm deep, funnel-shaped to slightly campanulate, tubular at the base, pale brown. Conidia 21–26 × 2.5–3.5 μm (mean ± SD = 23.6 ± 1.3 × 3.0 ± 0.2 μm), falcate, with an inconspicuous basal scar, 1-septate, hyaline, with a straight or gently curved setula at each end, 12–18 μm long, positioned terminally at the apex and slightly subterminally at the base, conidia accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Specimen examined: MALAYSIA, Selangor, Kepong Forest Reserve, on decaying leaves, October 1986, A.J. Kuthubutheen KUM 534 (holotype IMI 312436, ex-type strain IMI 312436).
Habitat and geographical distribution: Saprobe on decaying leaves known from Southeast Asia, Malaysia [
19]. According to GlobalFungi, the identical sequences were found in 13 soil samples collected in forests of the Caribbean (12 samples, Puerto Rico, Trinidad and Tobago [
119,
185] and South Asia (one sample, India) [
117]. The locations have tropical climate (MAT avg. 22 °C, MAP 3124 mm).
Notes: Kuthubutheen [
19] listed two specimens with different localities in the protologue, however, only one number to indicate the holotype IMI 312436. By comparing the protologue with the type material deposited in the Kew herbarium, it was found that the locality of a specimen listed second in order is the place of collection of the holotype. Kuthubutheen [
19] did not indicate an ex-type strain of his new species. Strain of
S. malaysiana, with identical designation IMI 312436 as the holotype and accompanying metadata, deposited by A.J. Kuthubutheen, was obtained from the CABI culture collection. We confirm its identity and connect the holotype with the ex-type strain in this study.
Stilbochaeta novae-guineensis closely resembles S. malaysiana, but differs from it by shorter conidia. Stilbochaeta aquatica is also comparable with S. malaysiana, but differs in smaller 0–1-septate conidia.
Stilbochaeta novae-guineensis (Matsush.) Réblová & Hern.-Restr.,
comb. nov. MycoBank MB 842217 (
Figure 39 and
Figure 40).
Basionym. Codinaea novae-guineensis Matsush., Microfungi of the Solomon Islands and Papua-New Guinea: 14. 1971.
≡ Dictyochaeta novae-guineensis (Matsush.) A.I. Romero, Boln Soc. argent. Bot. 22: 76. 1983.
Description on the natural substrate: Colonies effuse, brown, composed of setae and conidiophores, mycelium semi-immersed. Anamorph. Setae 95–162 μm long, 3–5 μm wide near the swollen base, arise singly or in groups of two from dark brown stromatic cells, erect, straight or flexuous, septate, smooth, dark brown and thick-walled, paler and thinner-walled toward the apex, unbranched, apex pale brown, sterile or with a terminal phialidic opening. Conidiophores 27.5–58 μm long, 3–5 μm wide near the base, macronematous, arise singly or in groups of 5–7 from stromatic cells around the base of the setae, erect, straight or slightly bent, septate, smooth, medium to pale brown, paler toward the tip. Conidiogenous cells 17–30 × 2.5–5 μm tapering to 1–2 μm below the collarette, integrated, terminal, mono- or polyphialidic with 1–2(–3) lateral opening, extending sympodially, cylindrical, medium to pale brown; collarettes 2.5–5 μm wide, 1.5–3.5 μm deep, funnel-shaped, pale brown, the apical part often evanescent or broken off and attached to the conidial mass. Conidia 14–19 × 2–4 μm (mean ± SD = 17.1 ± 3.1 × 1.6 ± 0.3 μm), falcate to oblong-falcate, with an inconspicuous basal scar, 0–1-septate, hyaline, with a straight or gently curved setula at each end, 6–12 μm long, conidia accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Culture characteristics: On CMD: colonies 25–25 mm diam, circular, flat, margin fimbriate, lanose to cobwebby, zonate, beige-brown to isabelline with darker concentric rings, reverse dark ivory. On MLA: colonies 32–35 mm diam, circular, convex, margin entire, lanose, floccose, finely furrowed centrally, whitish-grey to grey-brown, with a prominent outer zone of submerged growth, reverse olivaceous brown. On OA: colonies 49–68 mm diam, circular, raised, margin entire, velvety-lanose, floccose, with a central zone of sparse growth, whitish to dark grey-brown, sometimes the dark pigment in subsurface mycelium is absent, colonies becoming partially mucoid, whitish-beige, reverse pale brown to dark olivaceous grey. On PCA: colonies 49–52 mm diam, circular, flat, margin fimbriate to lobate, mucoid centrally, cobwebby toward the margin, isabelline to whitish-beige, pale ochre-brown at the margin, reverse of the same colors. Sporulation was moderate on MLA, OA and PCA, absent on CMD.
Colonies on CMA with U. dioica stems effuse, hairy, mycelium composed of branched, septate, hyaline to subhyaline hyphae 2–3 μm diam. Anamorph. Setae, conidiophores, conidiogenous cells and conidia similar to those from nature. Setae 100–202 μm long, 3–5 μm wide near the base, sterile or with a terminal or several lateral phialidic openings. Conidiophores 24–92 μm long, 3–5 μm wide near the base, arise in fascicles of 2–5 from knots of dark hyphal cells around the base of the setae, unbranched. Conidiogenous cells 10–30 × 2.5–5 μm tapering to 1.5–2 μm below the collarette, mono- or polyphialidic with 1–5 lateral openings while internally septa can be formed, pale brown to subhyaline; collarettes 2–4 μm wide, 1.5–3.5 μm deep, funnel-shaped, subhyaline. Conidia 14.5–33 × 2–3.5 μm (mean ± SD = 19.86 ± 2.97 × 3.42 ± 0.34 μm), falcate to oblong-falcate, 0–1-septate, setulae 3–14 μm long, accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Specimens examined: PUERTO RICO, on decaying wood, 19 July 2018, M. Hernández-Restrepo M.H.R. 18004 (CBS H-24746, culture CBS 147515); Ibid., on decaying twig, 19 July 2018, M. Hernández-Restrepo M.H.R. 18018 (CBS H-24748, culture CBS 147517).
Habitat and geographical distribution: Saprobe on decaying leaves, wood and bark of
Castanopsis sp. and other unidentified hosts, found in the tropical geographical areas of Africa, Australasia, the Caribbean and South America: Brazil, Cuba, Ivory Coast, Puerto Rico and Solomon Islands ([
17,
121,
134,
146,
147,
180,
186], this study).
According to GlobalFungi, the identical sequences were found in six soil samples in forests of South America (Peru) [
116] and the Caribbean (Puerto Rico) [
119]. The sites have tropical climate (MAT avg. 22 °C, MAP avg. 3059 mm).
Notes: Onofri and Zucconi [
180] emended the species diagnosis with polyphialidic conidiogenous cells, which were described as exclusively monophialidic in the protologue [
17]. In our material of
S. novae-guineensis, the conidiogenous cells were predominantly monophialidic, occasionally with 1–2(–3) lateral phialidic apertures. When grown in culture, the polyphialides with up to five lateral openings formed frequently.
Stilbochaeta malaysiana resembles
S. novae-guineensis but differs in longer conidia. It is challenging to distinguish
S. novae-guineensis from
S. septata as the size of their conidia considerably overlaps; for their comparison see notes to the latter species.
Stilbochaeta ramulosetula (Kuthub.) Réblová & Hern.-Restr.,
comb. nov. MycoBank MB 842218. (
Figure 34 and
Figure 41).
Basionym. Dictyochaeta ramulosetula Kuthub., Trans. Br. mycol. Soc. 89: 353. 1987.
Typification: MALAYSIA, Pahang, Lepar Forest Reserve, on decaying submerged leaf, July 1986, A. J. Kuthubutheen KUM 515 (holotype IMI 312058). MALAYSIA, Pahang, Lepar Forest Reserve, on decaying submerged leaf, July 1986, A. J. Kuthubutheen (epitype designated here MBT 10004619, IMI 313452 dried culture and microscopic slide, ex-epitype culture IMI 313452).
Culture characteristics: On CMD: colonies 29–31 mm diam, circular, flat, margin fimbriate, cobwebby, mucoid toward the periphery, whitish, reverse isabelline. On MLA: colonies 26–30 mm diam, circular, raised, fimbriate to weakly undulate, velvety-lanose, floccose, finely furrowed, whitish or with irregular salmon patches at the centre, ochre-beige at the margin, pale ochre pigment diffusing into agar, reverse pale ochre-beige. On OA: colonies 17–20 mm diam, circular, flat, margin entire, velvety to cobwebby, aerial mycelium on the inoculation block with numerous cinnamon exudates, whitish, pale ochre pigment diffusing into agar, reverse pale cinnamon. On PCA: colonies 27–33 mm diam, circular, flat, margin rhizoidal, cobwebby becoming mucoid, ochre-beige, reverse of similar color. Sporulation was moderate on CMD, absent on MLA, OA, and PCA.
Colonies on CMD effuse, hairy, mycelium composed of branched, septate, hyaline hyphae 1.5–3 μm diam. Anamorph. Setae 170–225 μm long, 4.5–5 μm wide near the base, single or arise in groups of two from dark hyphal stromata, erect, straight or flexuous, septate, smooth, brown and thick-walled, paler and thinner-walled toward the apex, unbranched, apex subhyaline, bluntly rounded, sterile, occasionally terminating into a monophialide. Conidiophores 46–97 μm long, 4–5 μm wide near the base, macronematous, arise in fascicles of 3–10 from dark hyphal cells, usually aggregated around the base of the setae, erect, straight or flexuous, septate, smooth, pale brown, paler toward the tip. Conidiogenous cells 12.5–19(–33) × 3.5–4.5 μm tapering to 1.5–2 μm below the collarette, integrated, terminal, mono- rarely polyphialidic with one lateral opening while internally septa can be formed, extending sympodially, rarely percurrently, cylindrical, pale brown, subhyaline at the apex; collarettes 4.5–6.5 μm wide, 1.5–3.5 μm deep, widely funnel-shaped, slightly stipitate, pale brown to subhyaline. Conidia 16–19.5 × 2.5–3.5 μm (mean ± SD = 17.9 ± 1.1 × 3.0 ± 0.4 μm), oblong-falcate, 1-septate, slightly constricted at the septum, hyaline, rounded at each end, with a bifid setula at each end, occasionally with a bifid setula at one end and a trifid setula at the other, setulae straight or gently curved, 8–15 μm long, positioned terminally, conidia accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Habitat and geographical distribution: Saprobe on submerged leaves, known from Malaysia only [
19]. According to GlobalFungi, the identical sequences were found in two soil samples from the forest biome in the Caribbean (Trinidad and Tobago) [
185]. The site has a tropical climate (MAT avg. 26 °C, MAP avg. 2511 mm).
Notes: Kuthubutheen [
19] listed two specimens in the protologue of
S. ramulosetula but only one number to indicate the holotype IMI 312058. Based on comparison with the holotype material in the Kew herbarium, the locality of the first specimen is the place of collection of the holotype. An unpublished strain of
S. ramulosetula IMI 313452, collected in the same locality as the holotype and deposited by A.J. Kuthubutheen, was obtained from the CABI culture collection. A specimen in the Kew herbarium with the identical accession number, deposited as the microscopic slide and dried culture, is selected as the epitype in this study.
Stilbochaeta ramulosetula is well distinguished from other members of the genus by bifid and trifid setulae inserted at each end of conidia. It is closely related to S. malaysiana and S. novae-guineensis; these species share 1-septate conidia and setae that are predominantly sterile but occasionally terminate into a monophialide.
Stilbochaeta septata (B. Sutton & Hodges) Réblová & Hern.-Restr.,
comb. nov. MycoBank MB 842219. (
Figure 34 and
Figure 42).
Basionym. Codinaea septata B. Sutton & Hodges, Nova Hedw. 26: 520. 1975.
≡ Dictyochaeta septata (B. Sutton & Hodges) Aramb. & Cabello, Mycotaxon 34: 682. 1989. (Nom. inval., Art. 41.4).
≡ Dictyochaeta septata (B. Sutton & Hodges) Whitton, McKenzie & K.D. Hyde, Fungal Divers. 4: 148. 2000.
Description on the natural substrate: Colonies effuse, hairy, brown composed of setae and conidiophores, mycelium immersed. Anamorph. Setae 245–260 μm long, 3–5 μm wide near the base, arise singly or in groups of 2–3 from knots of dark brown hyphal cells, erect, straight or flexuous, septate, smooth, dark brown and thick-walled, paler and thinner-walled toward the apex, unbranched, apex subhyaline to hyaline, sterile, bluntly rounded. Conidiophores macronematous, unbranched, usually reduced to sessile or stalked conidiogenous cells, 34–37 × 3–4 μm tapering to ca. 1.5 μm below the collarette, arise singly or in groups of 2–6 from knots of hyphal cells around the base of the setae, smooth, integrated, terminal, mono- occasionally polyphialidic with one lateral aperture, extending sympodially, cylindrical, pale brown to subhyaline, paler toward the tip; collarettes 2.5–3 μm wide, ca. 2 μm deep, funnel-shaped, subhyaline. Conidia 14–15.5 × 2–2.5(–3) μm (mean ± SD = 14.9 ± 0.6 × 2.3 ± 0.2 μm), falcate, tapering toward both ends, with an inconspicuous basal scar, 1-septate, hyaline, with a straight or gently curved setula at each end 5.5–9 μm long, inserted terminally at the apex, subterminally at the base, conidia accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Culture characteristics: On CMD: colonies 28–30 mm diam, circular, flat, margin entire to weakly fimbriate, velvety-lanose, mucoid at the margin, white becoming irregularly beige to brown toward the periphery due to densely aggregated sporulating conidiophores, isabelline at the margin, with a prominent outer zone of submerged growth, reverse pale ochre, isabelline toward the periphery. On MLA: colonies 33–34 mm diam, circular, raised or crateriform, margin entire, velvety-lanose, furrowed, later developing cracks in folds, white, sepia at the margin, pale ochre pigment diffusing into the agar, reverse dark cinnamon. On OA: colonies 49–50 mm diam circular, raised, margin entire, velvety-lanose, locally mucoid, smooth, white becoming ash grey due to densely aggregated sporulating conidiophores, cinnamon at the margin, sometimes pale cinnamon pigment diffusing into agar, reverse ochre-brown. On PCA: colonies 48–49 mm diam, circular, flat, margin entire, velvety becoming cobwebby, mucoid toward the periphery, white-ochre, pale olivaceous isabelline at the margin, with a zone of submerged growth, reverse ochre. Sporulation was abundant on OA and PCA, moderate on MLA, absent on CMD.
Colonies on CMD effuse, hairy, mycelium composed of branched, septate, hyaline to subhyaline hyphae 1.5–3 μm diam. Anamorph. Setae similar to those from nature, 138–192 μm long, 3.5–4.5 μm wide near the base, arise in groups from knots of dark brown cells, unbranched, apex pale brown to subhyaline, sterile, bluntly rounded or terminating into a mono- or polyphialide. Conidiophores 34–76(–92) μm long, 3–4.5 μm wide near the base, macronematous, single or arise in fascicles of 2–4 from knots of dark hyphal cells around the base of the setae, erect, straight or flexuous, septate, smooth, unbranched, brown, paler toward the tip. Conidiogenous cells 20–26.5 × 3.5–4 μm tapering to 1.5–2 μm below the collarette, integrated, terminal, monophialidic, occasionally polyphialidic with 1–2 lateral openings, extending percurrently and sympodially, subcylindrical, pale brown, subhyaline toward the tip; collarettes 3.5–4 μm wide, 1.5–2.5 μm deep, funnel-shaped, subhyaline. Conidia similar to those from nature, 13.5–16 × 2–3 μm (mean ± SD = 14.7 ± 0.8 × 2.3 ± 0.3 μm), falcate, 1-septate, setulae 6.5–10 μm long, accumulate in slimy whitish fascicles. Teleomorph. Unknown.
Specimens examined: AUSTRALIA, New South Wales, Yarramulong, Nunkeri Native Flowers, on a fallen leaf of Melaleuca viminalis, 23 August 1999, K.A. Seifert K.A.S. 1112 (culture CBS 146716). CHILE, on leaves of Eucalyptus grandis × urophylla, June 2010, M.J. Wingfield (culture ex-epitype CBS 143386).
Habitat and geographical distribution: Saprobe on leaves of
Eucalyptus grandis,
E. grandis ×
urophylla,
Eucalyptus sp. and
Melaleuca viminalis, known from Africa, Australasia and South America: Australia, Brazil, Chile, and South Africa ([
31,
81,
187], this study).
According to GlobalFungi, the identical sequences were found in 33 samples, in forest soil in Australia (29 samples) [
169,
188,
189] and rhizosphere soil from grasslands in Africa (four samples, South Africa) [
190]. The sites have temperate climate (MAT avg. 16.4 °C, MAP avg. 884 mm).
Notes: Our specimen CBS 146716 from Australia differs from the holotype described from fallen
Eucalyptus sp. leaves in Brazil [
31] in slightly longer setae and shorter conidia. The setae have sterile, subhyaline, obtusely rounded apex on material from nature, in culture the apex is modified into a mono- or polyphialide. The conidial length corresponds to the lower range given in the protologue, (14.5–)17.5–23 μm [
31]. In the present phylogeny, our strain grouped with the ex-epitype strain of
S. septata [
82]; they share nearly identical ITS and
tef1-α sequences, which differ in one base pair in each gene.
Stilbochaeta septata resembles
S. novae-guineensis in septate conidia with a simple setula at each end and sterile or fertile setae, but conidia of the latter species are 0–1-septate vs. 1–2-septate and slightly wider with longer setulae. It is difficult to distinguish
S. septata from
S. cangshanensis [
29] because the size of their setae, conidiophores and conidia overlap. The two species are shown to be closely related in the present phylogeny but are well distinguished using ITS and
tef1-α sequences. The genetic distances between the ex-type strain of
S. cangshanensis and ex-epitype strain of
S. septata are 4.0% in the ITS and 3.6% in
tef1-α genes corresponding to the sequence identities of 96.6% and 96.5%, respectively.
Stilbochaeta submersa (Z.L. Luo, K.D. Hyde & H.Y. Su) Réblová & Hern.-Restr., comb. nov. MycoBank MB 842220.
Basionym. Dictyochaeta submersa Z.L. Luo, K.D. Hyde & H.Y. Su, Fungal Divers. 99: 597. 2019.
For description and illustration, see Luo et al. [
29].
Habitat and geographical distribution: Saprobe on decaying wood, known only from Asia, China [
29]. The database GlobalFungi did not contain similar sequences (≥98%) of this species.
Notes: Although the conidia were described as aseptate, the photographs accompanying the protologue [
29] show 0–1-septate conidia, which is in accordance with the generic concept of
Stilbochaeta.
Stilbochaeta submersa resembles
S. aquatica [
28] in the conidial size and monophialidic conidiogenous cells, but the latter species differs in shorter conidiophores.
3.5.5. The Genus Tainosphaeria
Tainosphaeria F.A. Fernández & Huhndorf, Fungal Divers. 18: 44. 2005.
Notes:
Tainosphaeria is a holomorphic genus characterized by perithecial ascomata, unitunicate asci, hyaline, transversely septate ascospores, and
Codinaea-like anamorphs [
57]. Members of the genus form effuse colonies of pigmented conidiophores terminating into a mono- or polyphialides and have hyaline, aseptate conidia with setulae.
Tainosphaeria became heterogeneous after the acceptance of several anamorphic species with different morphology [
29,
30]. Réblová et al. [
49] re-evaluated the generic concept of
Tainosphaeria and showed correlation between molecular and morphological data, followed by the segregation of its species into three genera, namely
Tainosphaeria s. str.,
Phialoturbella [
49] and
Phialogeniculata [
191]. Li et al. [
192] described another two
Tainosphaeria species that do not match the concept of the genus. They are treated in this study, see
Tainosphaeriella.
Revision of three former
Codinaea species, namely
C. parva,
C. simplex, and
C. vulgaris, described by Hughes and Kendrick [
2] from decaying plant material from New Zealand revealed that they are remarkably similar to members of
Tainosphaeria. Although their cultures or DNA data are not available, based on a detailed morphological comparison of their type material, original descriptions, illustrations and data on geographical distribution, they were transferred to
Tainosphaeria and new combinations are proposed below. For additional information, see
Section 4.5.
Several other species, such as
Dictyochaeta brachysetula and
D. chinensis [
193],
D. coffeae [
8],
D. longispora [
2], and
D. unisetula [
194] resemble
Tainosphaeria in characters of conidiophores, conidiogenous cells and conidia to some extent. However, their cultures or molecular data to verify such relationships are not available.
Tainosphaeria parva (S. Hughes & W.B. Kendr.) Réblová & Hern.-Restr.,
comb. nov. MycoBank MB 842221. (
Figure 43).
Basionym. Codinaea parva S. Hughes & W.B. Kendr., N. Z. J. Bot. 6: 354. 1968.
≡ Dictyochaeta parva (S. Hughes & W.B. Kendr.) Hol.-Jech., Česká Mykol. 42: 204. 1988.
For description and illustrations, see Hughes and Kendrick [
2].
Specimen examined: NEW ZEALAND, West Coast region, Westland district, Pukekura, Lake Ianthe, on decaying bark of Weinmannia racemosa, 8 April 1963, S.J. Hughes 569d (isotype of Codinaea parva DAOM 93565d).
Habitat and geographical distribution:
Tainosphaeria parva is a saprobic species found mainly in the Southern Hemisphere. Originally, it was described from the bark of
Weinmannia racemosa in New Zealand [
2]. Additional specimens have been recorded on fallen leaves of
Nothofagus solandri also from New Zealand [
145] and
Pandanus hornei and
P. tectorius from Seychelles [
44]. Holubová-Jechová [
195] recorded this species on a dead petiole of
Calyptrogyne dulcis in Cuba.
Notes: Our observations of the isotype match the protologue of this species. Tainosphaeria parva is characterized by mono- and polyphialidic conidiogenous cells and conidia with short appendages, which are the shortest (2–4 μm long) among other species of the genus.
Tainosphaeria simplex (S. Hughes & W.B. Kendr.) Réblová & Hern.-Restr.,
comb. nov. MycoBank MB 842222. (
Figure 44).
Basionym. Codinaea simplex S. Hughes & W.B. Kendr., N. Z. J. Bot. 6: 362. 1968.
≡ Dictyochaeta simplex (S. Hughes & W.B. Kendr.) Hol.-Jech., Folia geobot. phytotax. 19: 434. 1984.
For description and illustrations, see Hughes and Kendrick [
2].
Specimen examined: NEW ZEALAND, West Coast region, Westland district, Pukekura, Lake Ianthe, on bark of Weinmannia racemosa, 8 April 1963, S.J. Hughes 560g (isotype of Codinaea simplex DAOM 96020g).
Habitat and geographical distribution:
Tainosphaeria simplex is a saprobe, originally described from bark and wood of an evergreen shrub
Weinmannia racemosa [
2]. In addition, it occurs on fallen leaves, herbaceous stems and decaying woody fruits of
Castanopsis sp.,
Cedrela odorata,
Didelotia idea,
Diospyros sanza-minika,
Dichapetalum toxicarium,
Eucalyptus sp.,
Fagus sylvatica,
Freycinetia banksia,
Memecylon donianum,
Nothofagus fusca,
Pandanus furcatus,
P. seychellarum,
P. tectorius,
Podocarpus macrophyllus,
Q. virginiana,
Quercus sp.,
Rubus sp., and
Socratea sp.
According to literature, it is one of the most common
Tainosphaeria species and has been recorded from many localities in the temperate, subtropical and tropical geographical zones. According to Kirk [
196],
T. simplex is the most common species of the genus in British Isles based on herbarium records in the Kew Herbarium. The species is known from Australia, Brazil, China, Cuba, Germany, Japan, Ivory Coast, Malaysia, New Zealand, Papua New Guinea, Seychelles, Solomon Islands, Taiwan, and United Kingdom [
2,
5,
17,
25,
31,
35,
36,
37,
44,
121,
134,
145,
146,
147,
148,
196,
197,
198].
A possible misinterpretation of the species concept of
T. simplex ([
2], as
Codinaea simplex) with
Codinaeella minuta is discussed in notes to the latter species.
Codinaeella minuta resembles
T. simplex, but differs in having an upper layer of setiform conidiophores. Analysis of environmental ITS sequences from GlobalFungi supports the theory that
Tainosphaeria and
Ca. minuta have distinct regions of distribution;
Tainosphaeria is primarily distributed in Australasia and Southeast Asia and in Central America, while
Ca. minuta has a much wider range of distribution in the temperate Northern Hemisphere. Given the similarity of the two species, published records on distribution of
T. simplex should be verified using DNA data.
Notes: Our observations of the isotype are in agreement with the protologue. The conidiophores were densely aggregated on the substrate surface, but slightly shorter ca. 38–68 μm long, mostly monophialidic or with 1(–2) phialidic apertures and conidia 15–20.5 × 2.5–3 μm with setulae 6–8(–9) μm long compared to the protologue.
Tainosphaeria vulgaris (S. Hughes & W.B. Kendr.) Réblová & Hern.-Restr.,
comb. nov. MycoBank MB 842223. (
Figure 45).
Basionym. Codinaea vulgaris S. Hughes & W.B. Kendr., N.Z. J. Bot. 6: 367. 1968.
≡ Dictyochaeta vulgaris (S. Hughes & W.B. Kendr.) Aramb. & Cabello, Mycotaxon 34: 682. 1989. (Nom. inval., Art. 41.4).
≡ Dictyochaeta vulgaris (S. Hughes & W.B. Kendr.) Whitton, McKenzie & K.D. Hyde, Fungal Divers. 4: 151. 2000.
Description on the natural substrate: Colonies effuse, olivaceous-brown to amber, composed of conidiophores, mycelium immersed. Anamorph. Setae absent. Conidiophores 62–120 μm long, 4–5 μm wide near the base, macronematous, crowded, arise singly or in groups, sometimes aggregate in loose columns, unbranched, erect, straight or flexuous, slightly geniculate, septate, smooth, brown, paler toward the apex. Conidiogenous cells 14.5–30 × 3–3.5 μm tapering to 1.5–2 μm below the collarette, integrated, terminal, mono- and polyphialidic with 2–15 lateral openings while internally septa can be formed, extending percurrently and sympodially, cylindrical, pale brown, subhyaline at the apex, usually bearing persistent remnants of the collarettes; collarettes 3–4 μm wide, 1.5–2 μm deep, funnel-shaped, subhyaline to pale brown, the apical part soon evanescent. Conidia (16–)18–20 × 2–2.5 μm (mean ± SD = 18.4 ± 1.0 × 2.3 ± 0.2 μm), falcate, tapering toward both ends, narrowly rounded apically, slightly truncate at the base with an inconspicuous basal scar, aseptate, hyaline, with straight or gently curved setula at each end 2.5–4.5 μm long, inserted terminally at the apex, subterminally at the base, conidia accumulate in slimy yellow-ochre fascicles. Teleomorph. Unknown.
Culture characteristics: Colonies on PCA effuse, mycelium composed of branched, septate, subhyaline to pale brown hyphae 1.5–2.5 μm diam. Setae absent, conidiophores, conidiogenous cells and conidia similar to those from nature. Conidiophores 90–120(–150) μm long, 3.5–4 μm wide above the base, unbranched, erect, straight, flexuous or undulating, mid brown, paler toward the apex. Conidiogenous cells 15–33 × 3–4 μm tapering to 1.5–2 μm below the collarette, integrated, terminal, monophialidic, cylindrical, pale brown, subhyaline at the apex; collarettes 3–4.5 μm wide, 2–2.5 μm deep, funnel-shaped, subhyaline. Conidia (16.5–)18–21 × 2.5(–3) μm (mean ± SD = 19.1 ± 1.5 × 2.6 ± 0.2 μm), falcate, hyaline, aseptate, setulae 2–3.5 μm long, aggregated in slimy ochre fascicles.
Specimens examined: NEW ZEALAND, Wellington province, Tongariro National Park, Silica Springs Track, Whakapapanui stream, on bark of Fuscospora cliffortioides, 5 March 1963, S.J. Hughes 432b (isotype of Codinaea vulgaris DAOM 97315b). NEW ZEALAND, West Coast region, Buller district, Victoria Forest Park, Lyell, Lyell Historical Walk, on decaying wood of a branch, 6 April 2005, M. Réblová M.R. 3447/NZ 731 (PDD 119682).
Habitat and geographical distribution: Saprobe on decaying bark and wood of a wide range of hosts in New Zealand, i.e.,
Fuscospora cliffortioides,
Nothofagus truncata,
Quintinia serrata,
Rubus sp.,
Weinmannia racemosa and other unknown hosts ([
2], this study), and leaves of
Pandanus seychellarum in Seychelles [
44].
Notes: The species was recollected in New Zealand and isolated into axenic culture, where it yielded sporulating conidiophores similar to those from nature. Although conidiogenous cells were polyphialidic with numerous successive lateral apertures on the nature substrate, conidiogenous cells remained monophialidic when grown in culture. Unfortunately, the strain is no longer viable. A description and illustration based on the recent material is given above; our observations agree with the protologue of T. vulgaris.
Tainosphaeria vulgaris is similar to T. parva in the geniculate appearance of conidiophores and relatively short conidial setulae up to 5 μm long. Tainosphaeria parva differs from T. vulgaris in shorter conidia. The conidia of T. vulgaris are arranged in ochre fascicles, while the conidial heads of other Tainosphaeria species were reported white.
3.5.6. The Genus Tainosphaeriella
Tainosphaeriella Réblová & Hern.-Restr., gen. nov. MycoBank MB 842003.
Type species. Tainosphaeriella aquatica (X.D. Yu, C.X. Li & H. Zhang) Réblová & Hern.-Restr.
Etymology: Tainosphaeria and -ella (L) diminutive but here used as a name-forming suffix, referring to fungi morphologically similar to Tainosphaeria.
Colonies on natural substrate effuse, pale to dark brown, mycelium partly superficial, partly immersed.
Anamorph. Setae absent. Conidiophores macronematous, mononematous, single, erect, straight or slightly flexuous, unbranched, septate, smooth, brown, paler toward the apex. Conidiogenous cells integrated, terminal, phialidic; collarettes flared, campanulate. Conidia falcate, cylindrical to fusiform or ellipsoidal-fusiform, slightly curved, transversely septate, hyaline, occasionally laurel green at maturity, with a straight or gently curved setula at each end, conidia accumulate in slimy whitish or ochre fascicles. (Adapted after Li et al. [
192]).
Teleomorph. Unknown.
Habitat and geographical distribution: Members of
Tainosphaeriella occur on submerged decaying wood and are only known from Southeast Asia, Thailand [
192].
Notes: The genus
Tainosphaeriella (
Ta.) is described for two species with 1–3-septate, setulate conidia borne on monophialidic conidiogenous cells with campanulate to almost disk-like collarettes and unbranched conidiophores. Originally, they were placed in
Tainosphaeria by Li et al. [
192], but molecular data do not support such relationship. In the present phylogenetic analysis, the two species are unrelated to
Tainosphaeria crassiparies, the type species, and other species of
Tainosphaeria. Instead, they grouped as a sister clade to
Phialogeniculata guadalcanalensis and
Phialoturbella spp.
Two
Dictyochaeta species,
D. macrospora and
D. variabilis [
23,
24] resemble
Tainosphaeriella. Both species are native to Malaysia and are found on submerged leaves or decaying palm fronds in terrestrial environments. Unfortunately, their living cultures or DNA sequences are not available for study.
Tainosphaeriella aquatica (X.D. Yu, C.X. Li & H. Zhang) Réblová & Hern.-Restr., comb. nov. MycoBank MB 842224.
Basionym. Tainosphaeria aquatica X.D. Yu, C.X. Li & H. Zhang, Phytotaxa 509: 60. 2021.
For description and illustration, see Li et al. [
192].
Habitat and geographical distribution: Saprobe on decaying submerged wood, known only from Southeast Asia, Thailand [
192].
Notes:
Tainosphaeria aquatica closely resembles
D. variabilis [
24], which differs in having longer conidia.
Tainosphaeriella thailandensis (W. Dong, C.X. Li & H. Zhang) Réblová & Hern.-Restr., comb. nov. MycoBank MB 842225.
Basionym. Tainosphaeria thailandensis [as thailandense] W. Dong, C.X. Li & H. Zhang, Phytotaxa 509: 61. 2021.
For description and illustration, see Li et al. [
192].
Habitat and geographical distribution: Saprobic on decaying submerged wood, known only from Southeast Asia, Thailand [
192].
Notes: This species has lightly pigmented conidia that turn laurel green at maturity, making it easily distinguishable from Ta. aquatica with conidia hyaline at maturity.
3.5.7. The Genus Xyladelphia
Xyladelphia Réblová, A.N. Mill. & Hern.-Restr., gen. nov. MycoBank MB 842004.
Type species. Xyladelphia longiseta (F.A. Fernández & Huhndorf) Réblová, A.N. Mill. & Hern.-Restr.
Etymology: Xýlon (Greek) wood, referring to the lignicolous habitat; adélphia (Greek), siblings, referring to two morphs occurring together.
Colonies on natural substrate effuse, hairy, brown, composed of setae, conidiophores and ascomata, mycelium immersed.
Anamorph. Setae grow singly, erect, straight or flexuous, septate, brown, unbranched, always sterile, tapering to an acute apex, sometimes two most apical cells dark brown. Conidiophores macronematous, mononematous, arise singly or in groups, scattered among the setae, unbranched, erect, straight or flexuous, septate, brown, smooth. Conidiogenous cells integrated, terminal, mono- and polyphialidic, extending percurrently and sympodially; collarettes flared. Conidia of two morphologically distinct types; macroconidia falcate to fusiform, aseptate, hyaline, with straight or gently curved setula at each end; microconidia (formed only in vitro) ellipsoidal, aseptate, hyaline to pale brown, without setulae; macroconidia and microconidia accumulate in slimy whitish fascicles.
Teleomorph. Ascomata perithecial, non-stromatic, superficial, subglobose to broadly ovoid, papillate, dark brown, setose; setae sterile, acute at the apex, identical to those of the anamorph. Ostiole periphysate. Ascomatal wall, carbonaceous, two-layered. Paraphyses septate, tapering. Asci unitunicate, cylindrical-clavate, shortly-stipitate, apically rounded, ascal apex with a non-amyloid apical annulus. Ascospores broadly fusiform to ellipsoidal, hyaline, aseptate or transversely septate, without gelatinous sheath or appendages (Adapted after Fernández and Huhndorf [
57]).
Habitat and geographical distribution: Saprobes on decaying plant matter, found in the Central, North and South America.
Notes: In the present phylogeny, Xyladelphia was shown distantly related to Codinaea and other morphologically similar genera. It formed a separate lineage near the base of the tree among taxa with non-setulate conidia.
Although the arrangement of setae and conidiophores is similar to members of
Codinaeella, the setae are darker in the uppermost part and have sterile, acute apex.
Xyladelphia clustered as a sister to
Dictyochaeta brevis [
30], which lack setae and somewhat resembles
Tainosphaeria. Due to inconsistencies in morphology, the latter species is not accepted in
Xyladelphia and for now we prefer to keep it separate until more morphological data, in vitro observations, teleomorph and other representatives are available for study.
Xyladelphia longiseta (F.A. Fernández & Huhndorf) Réblová, A.N. Mill. & Hern.-Restr., comb. nov. MycoBank MB 842226.
Basionym. Chaetosphaeria longiseta F.A. Fernández & Huhndorf, Fungal Divers. 18: 28. 2005.
For description and illustration, see Fernández and Huhndorf [
57].
Habitat and geographical distribution: Saprobic on decaying wood of twigs and branches, known from the tropical zone of Central and South America from Costa Rica, Ecuador and Puerto Rico, and also from the subtropical zone of the USA, South Carolina [
57].
Notes:
Xyladelphia longiseta is the only member of the genus and a known representative of this conspicuous morphotype. It somewhat resembles
Codinaea setosa [
2], which differs in septate conidia without setulae.
Dictyochaeta australiensis [
7] can be compared with the present species, but differs in longer setae that are undulate in the upper part and shorter conidia with longer setulae.


 ,
, 



















































{kind=link}
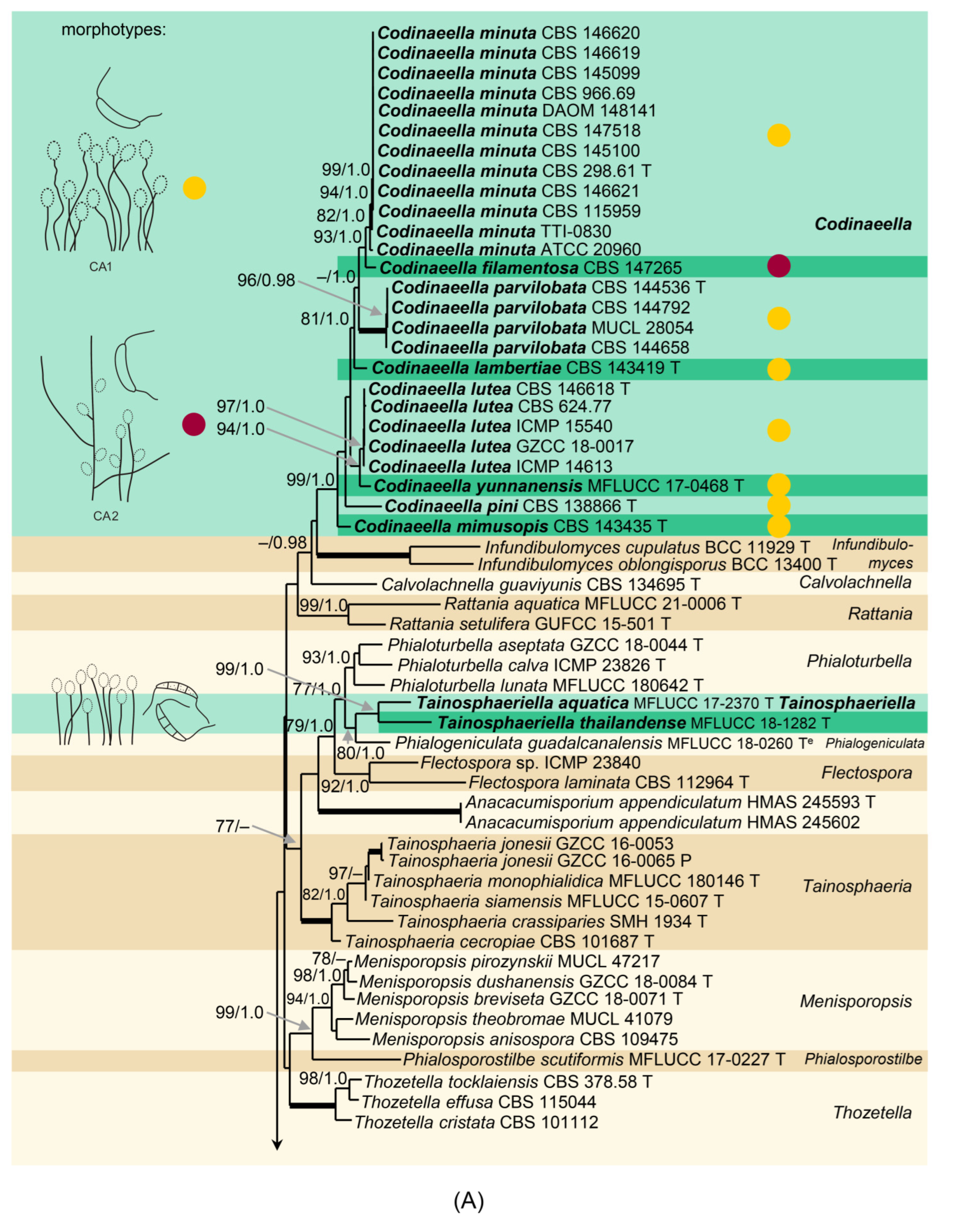{kind=link}
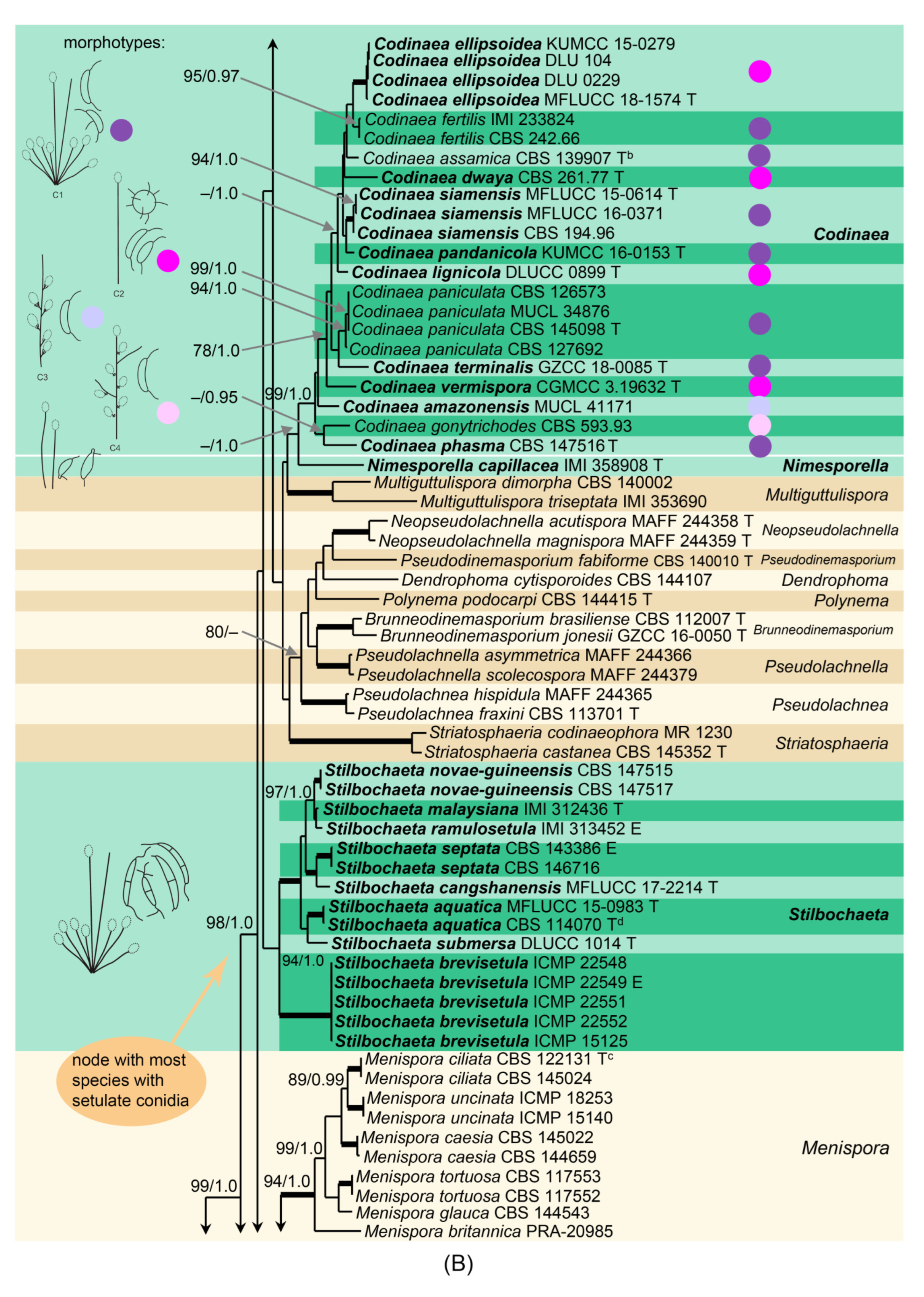{kind=link}
{kind=link}
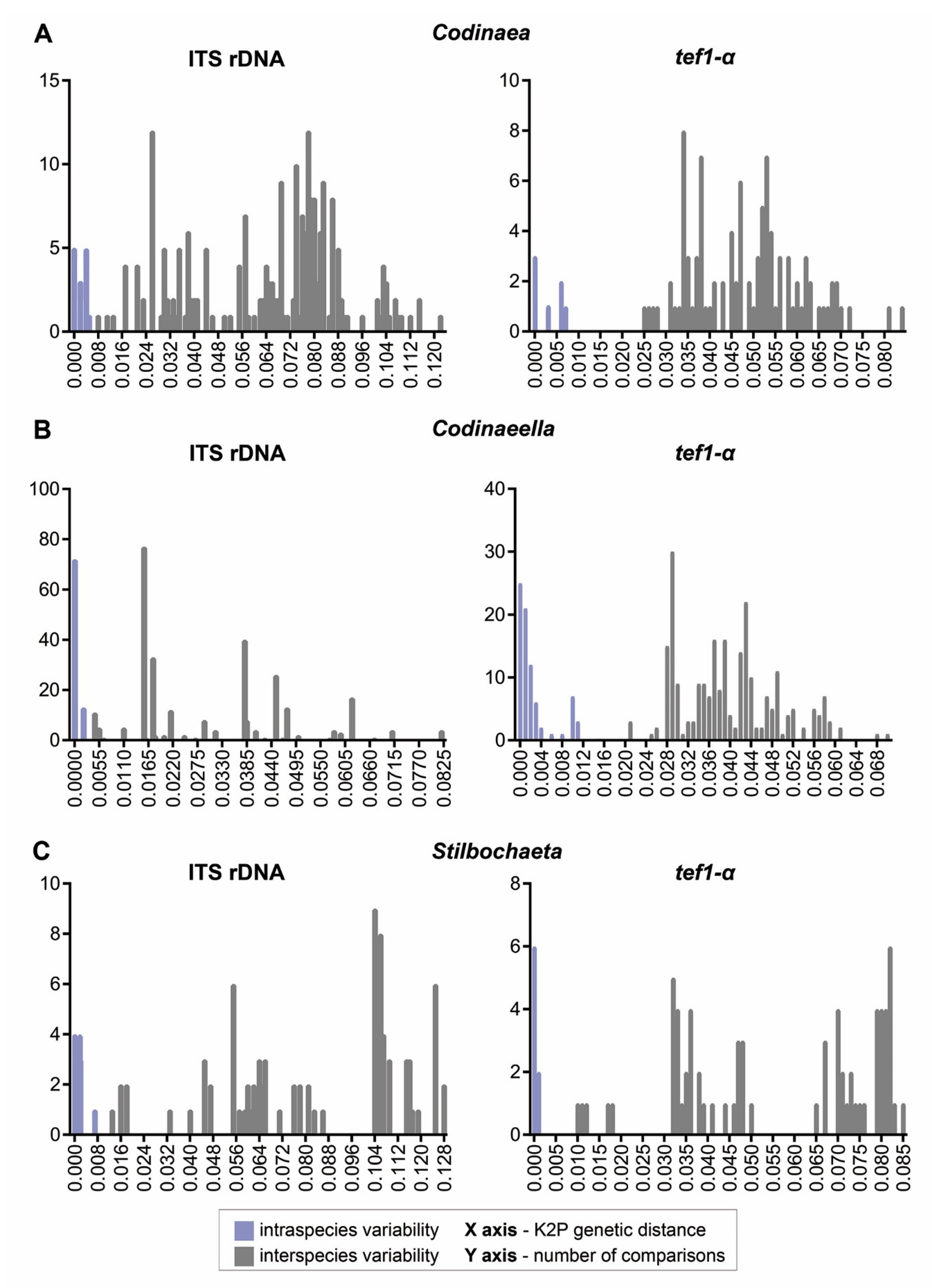{kind=link}
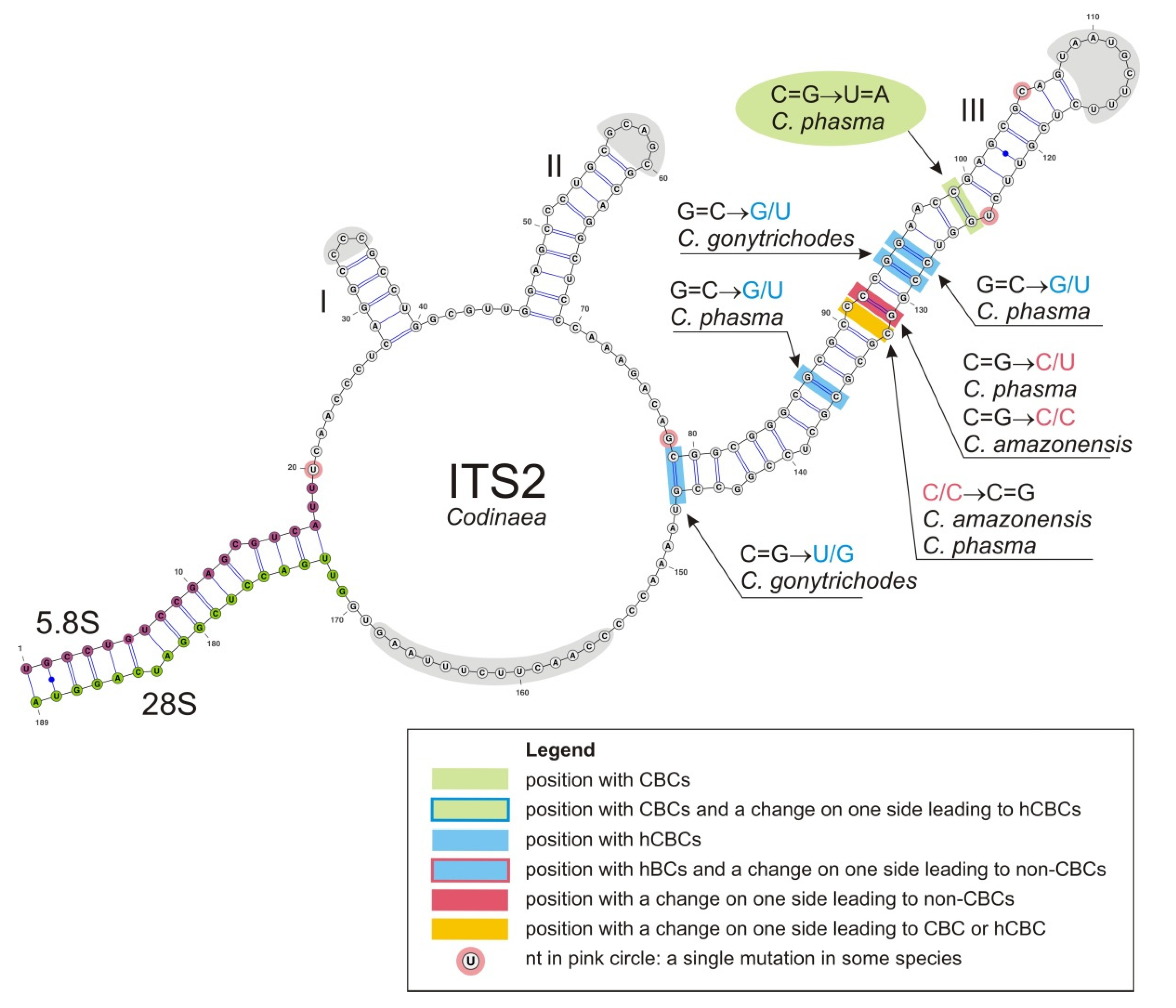{kind=link}
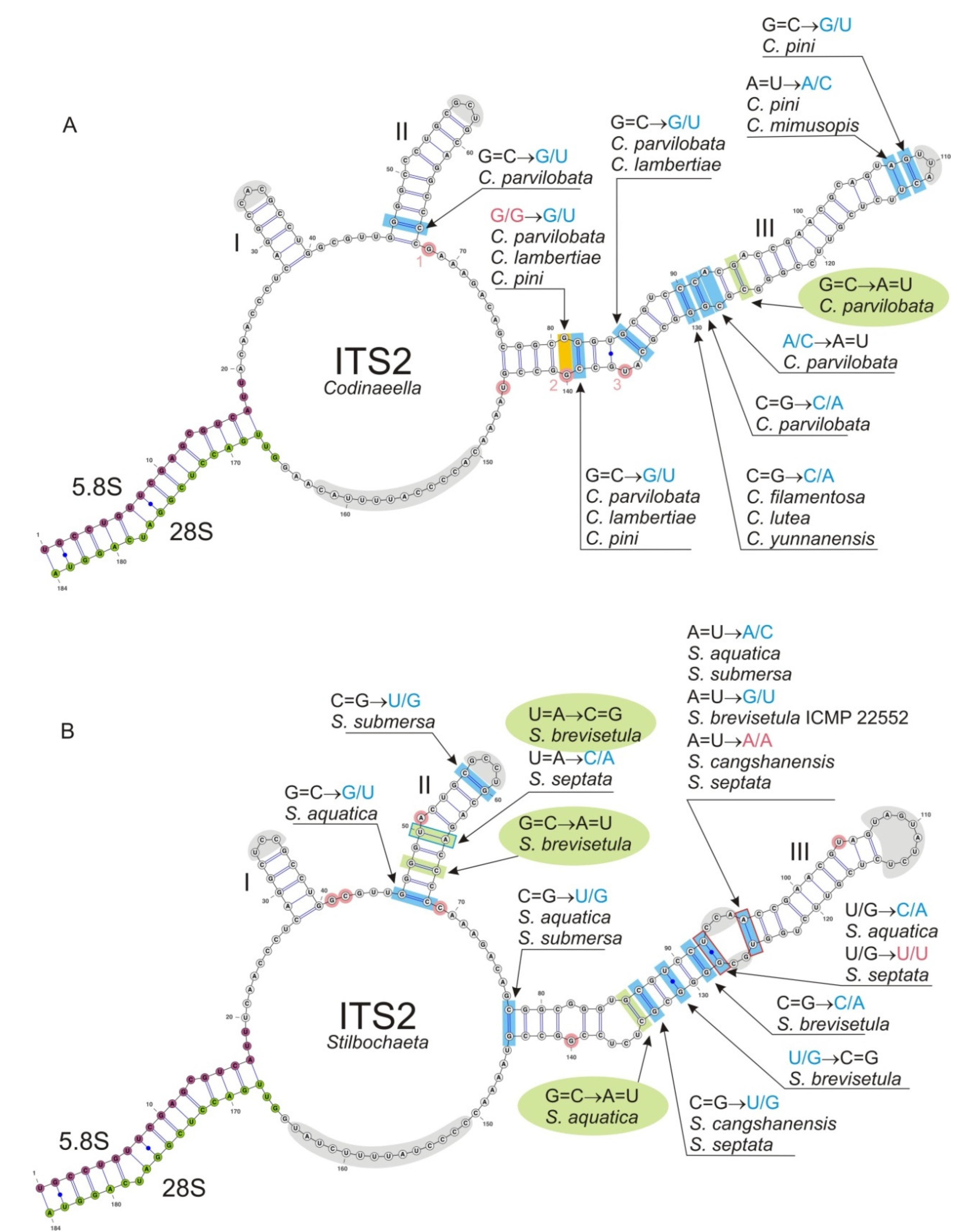{kind=link}
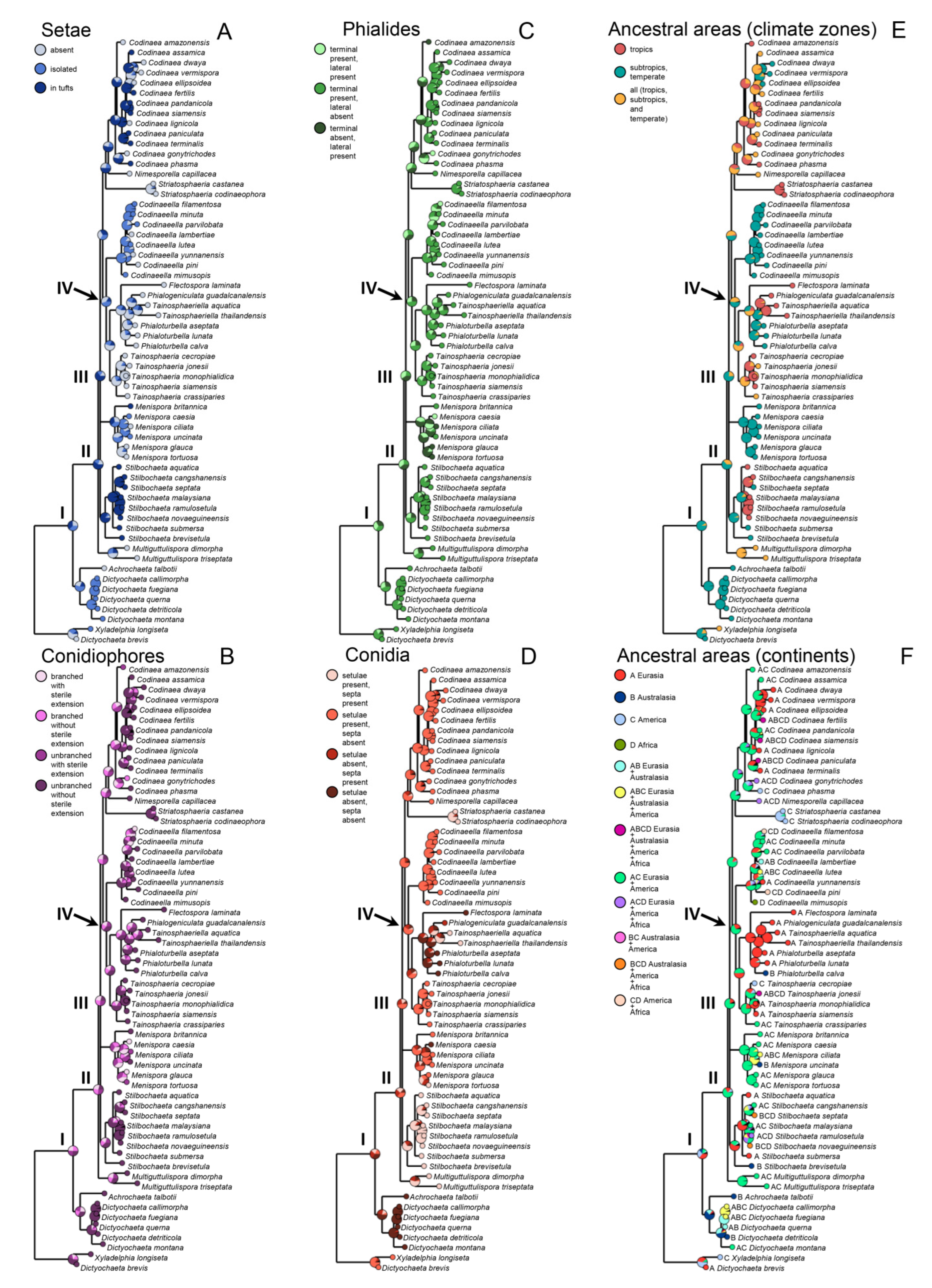{kind=link}
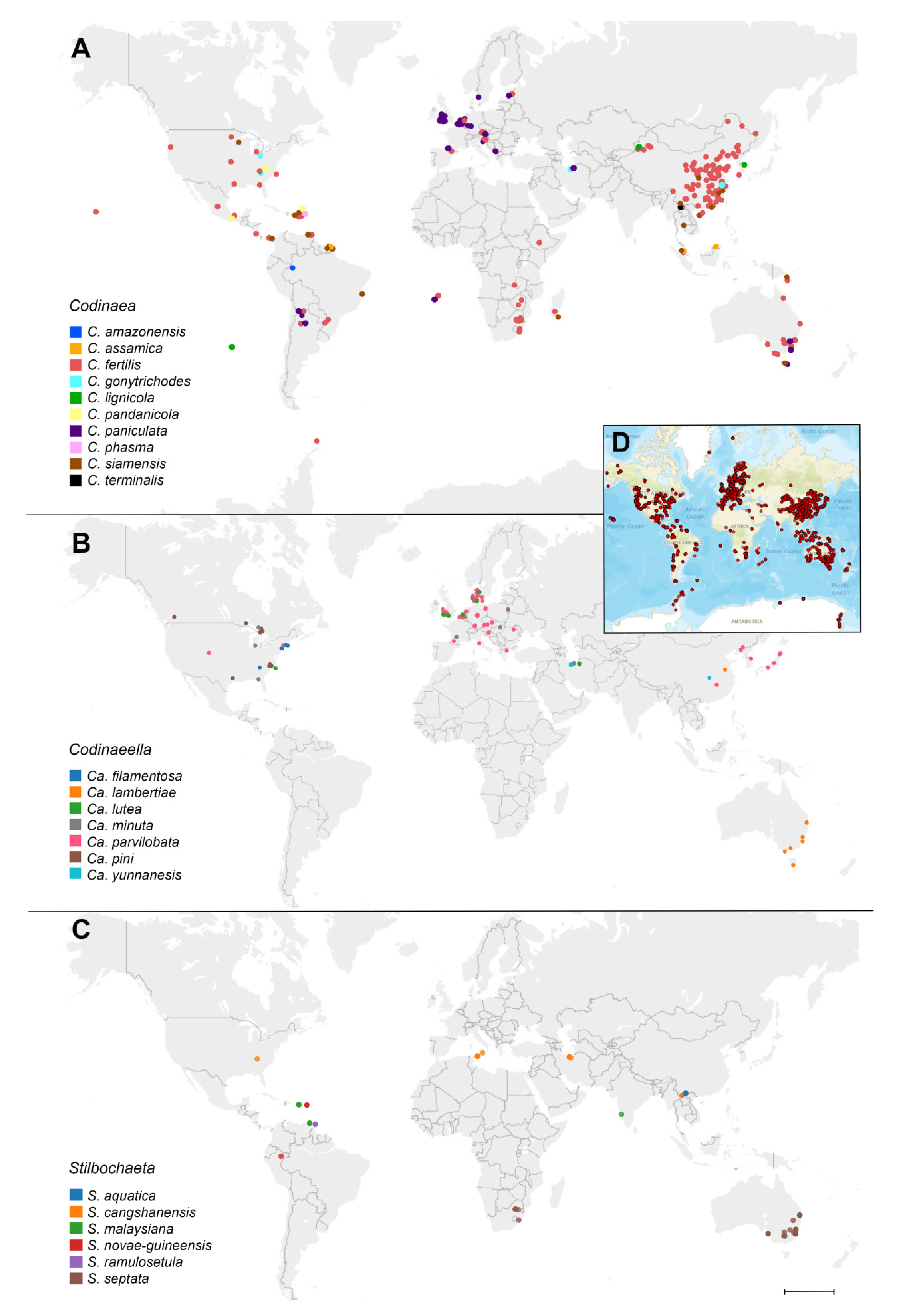{kind=link}
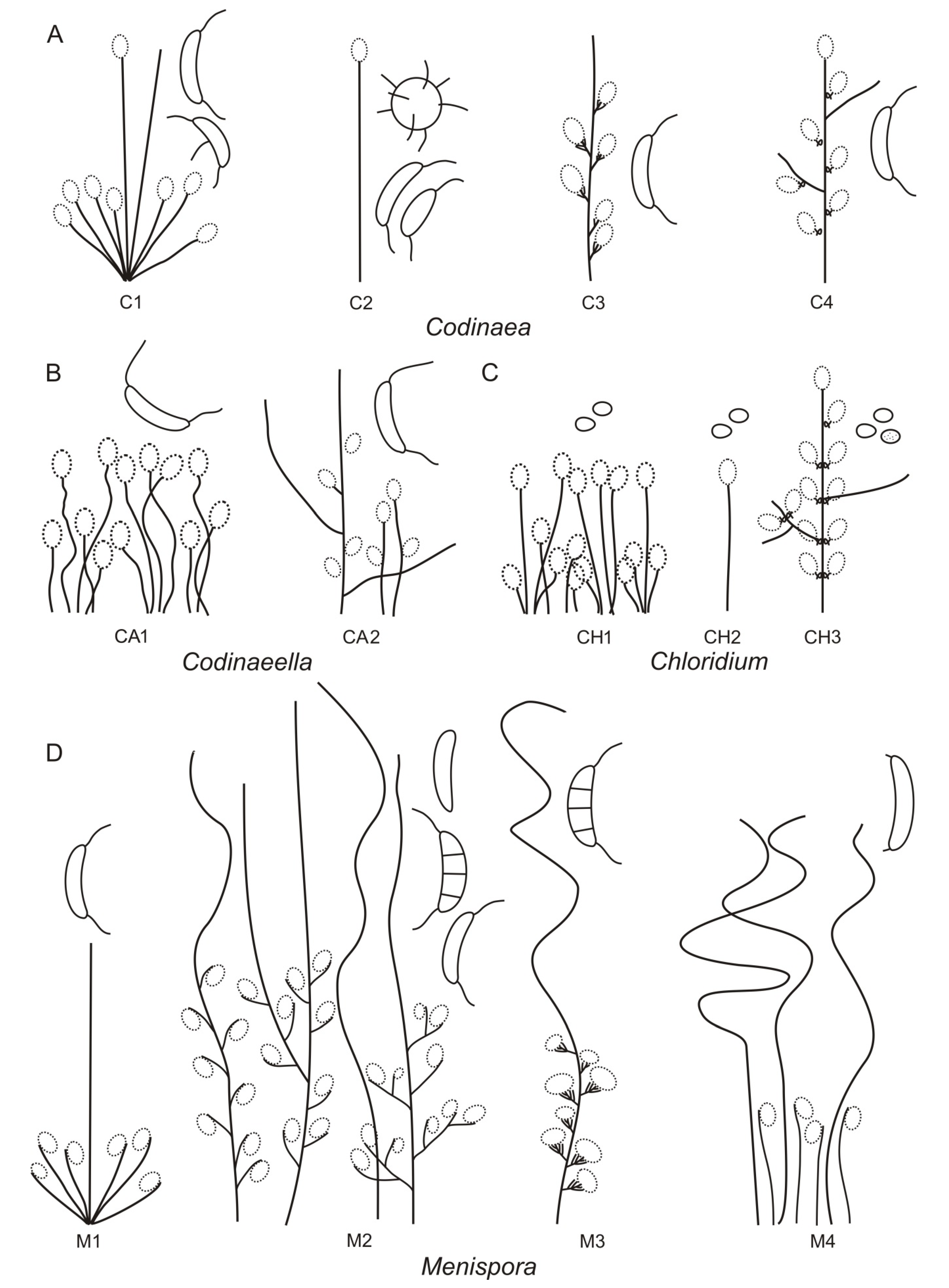{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
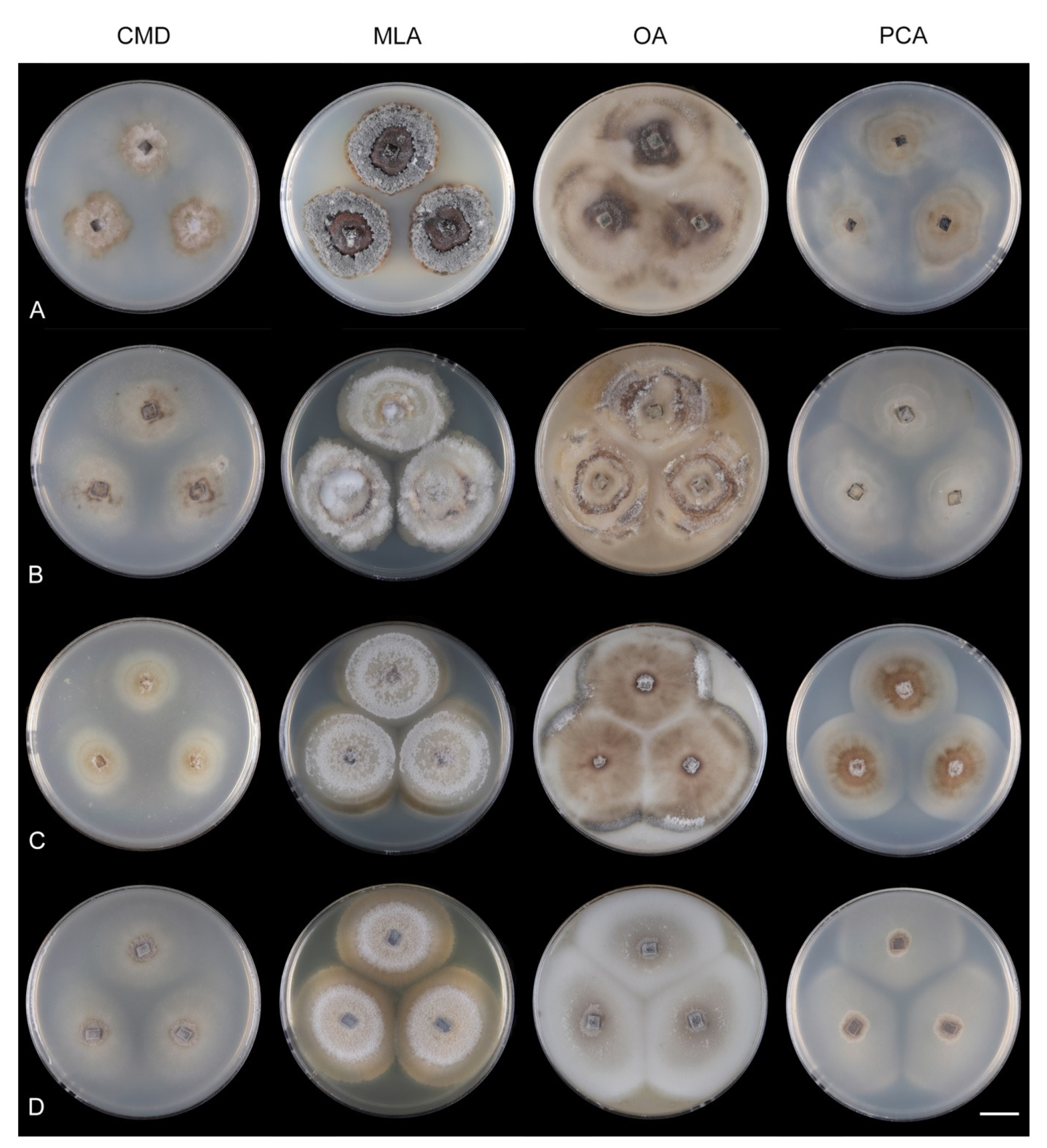{kind=link}
{kind=link}
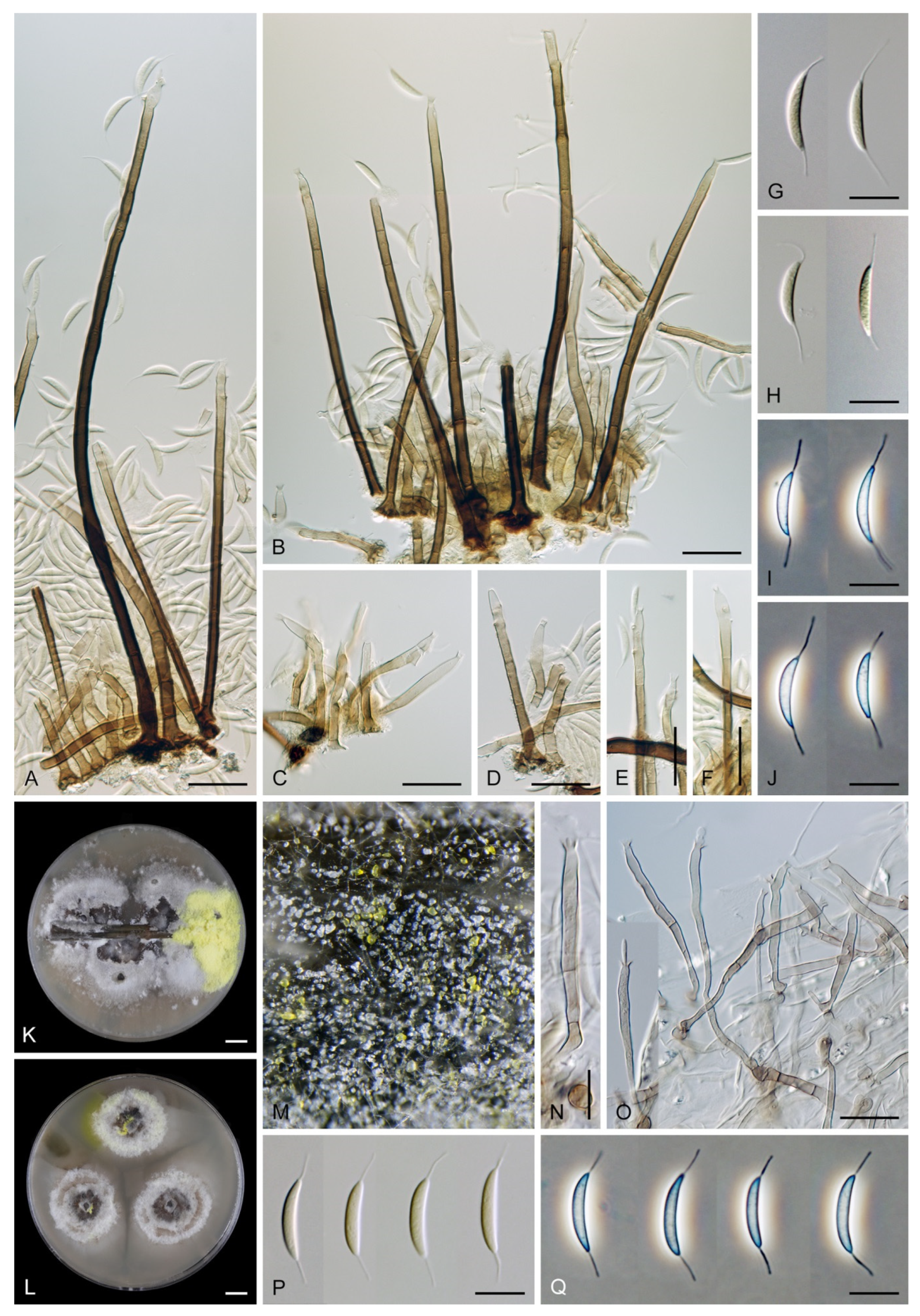{kind=link}
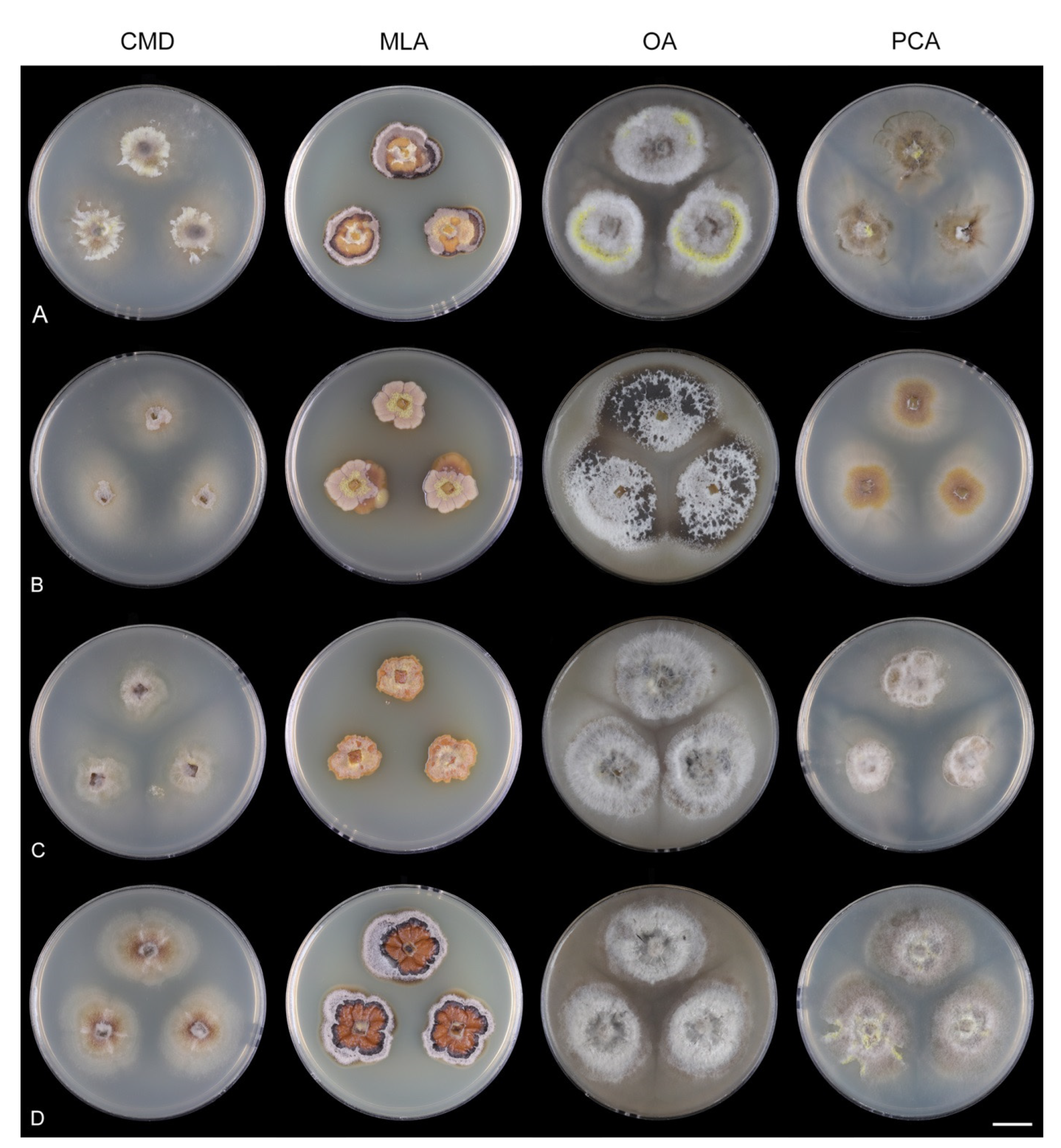{kind=link}
{kind=link}
{kind=link}
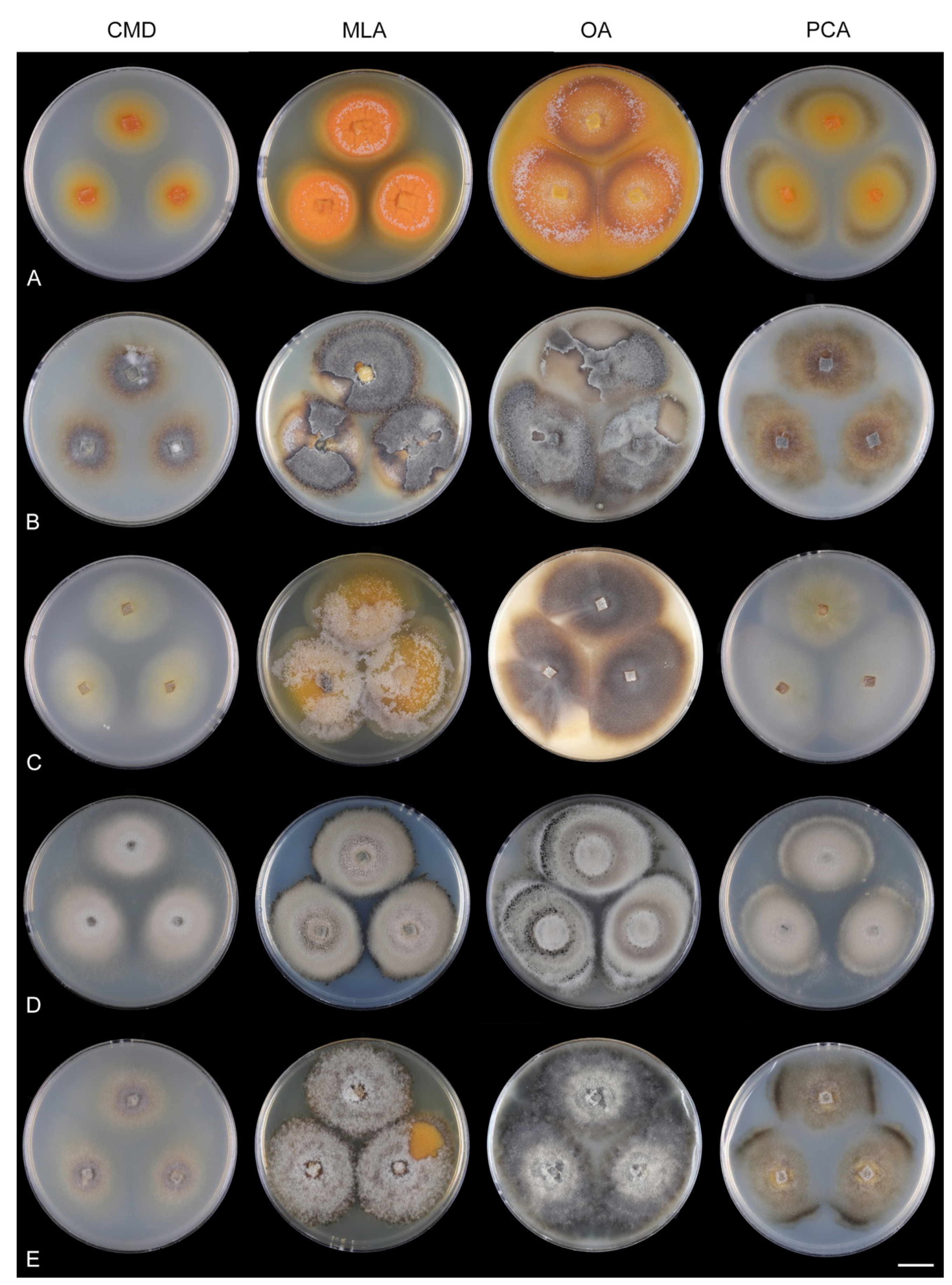{kind=link}
{kind=link}
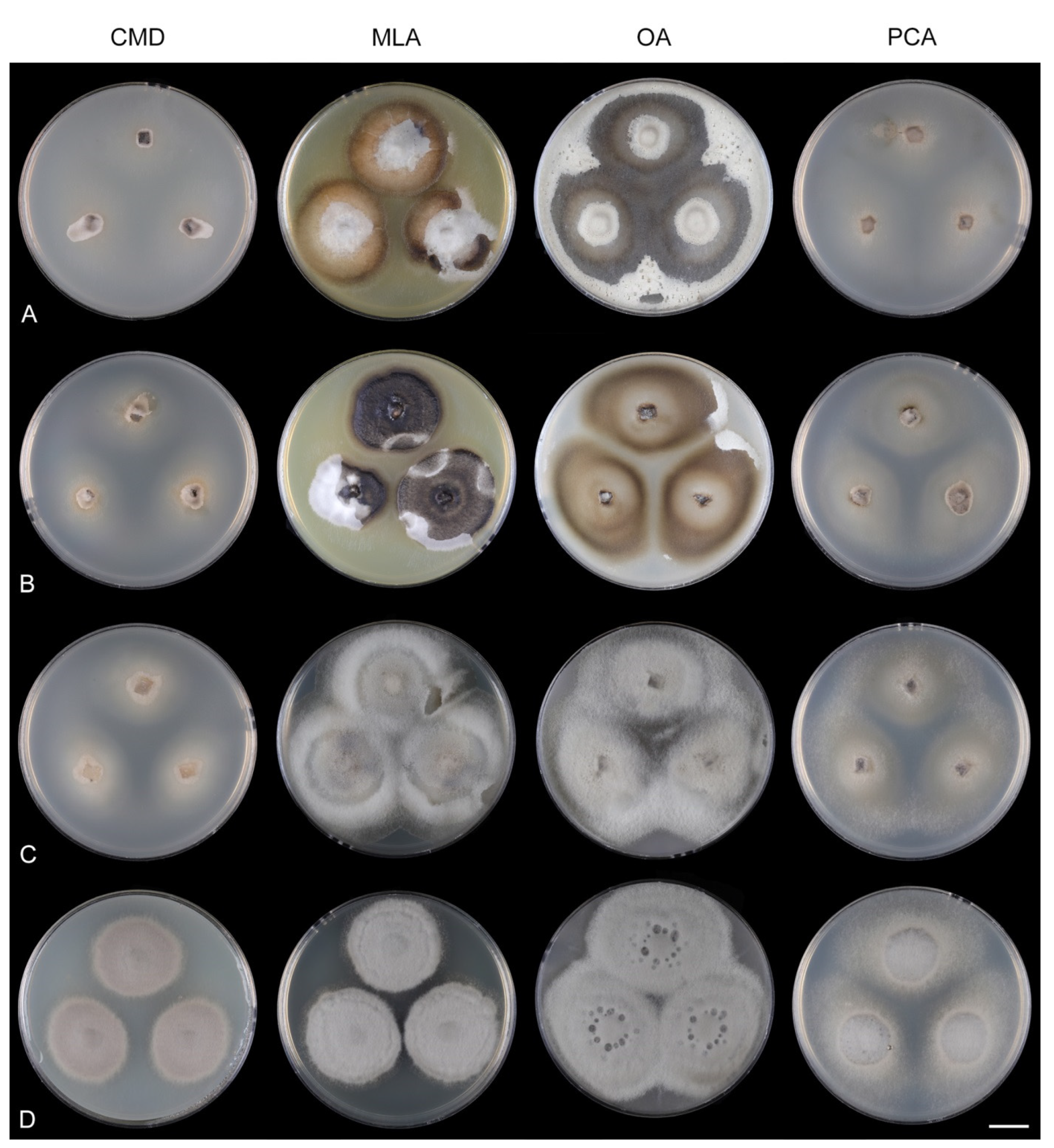{kind=link}
{kind=link}
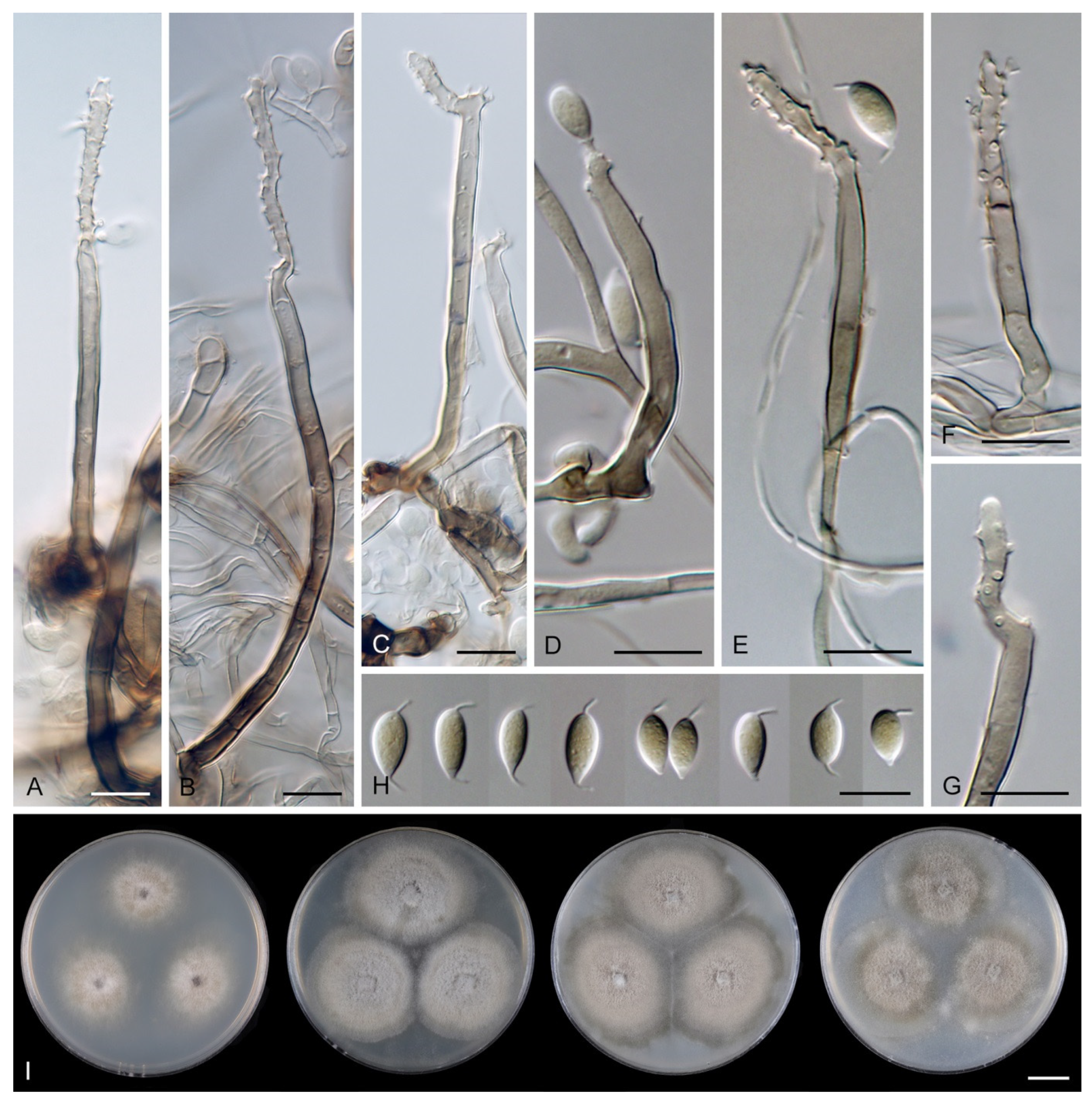{kind=link}
{kind=link}
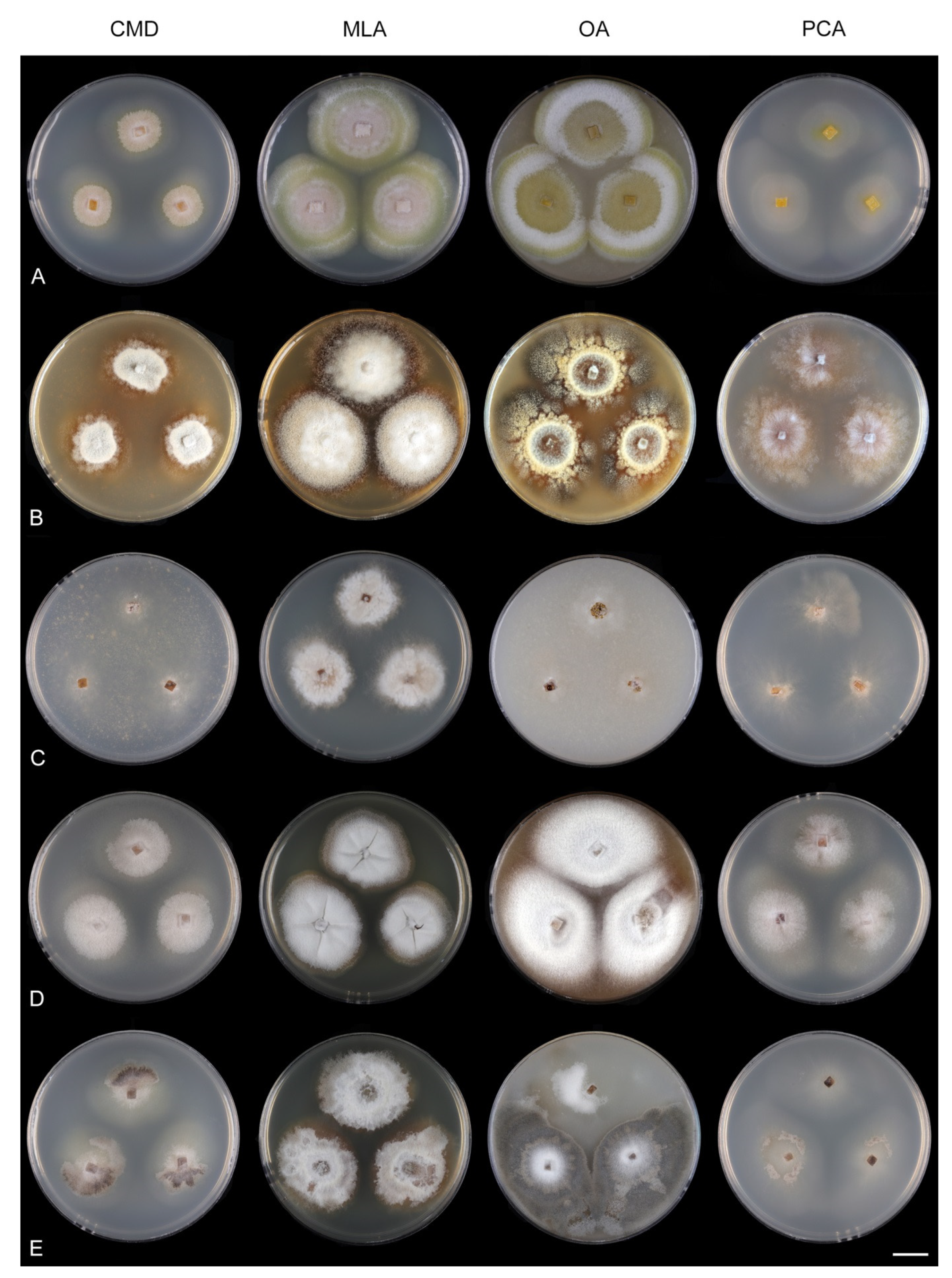{kind=link}
{kind=link}
{kind=link}
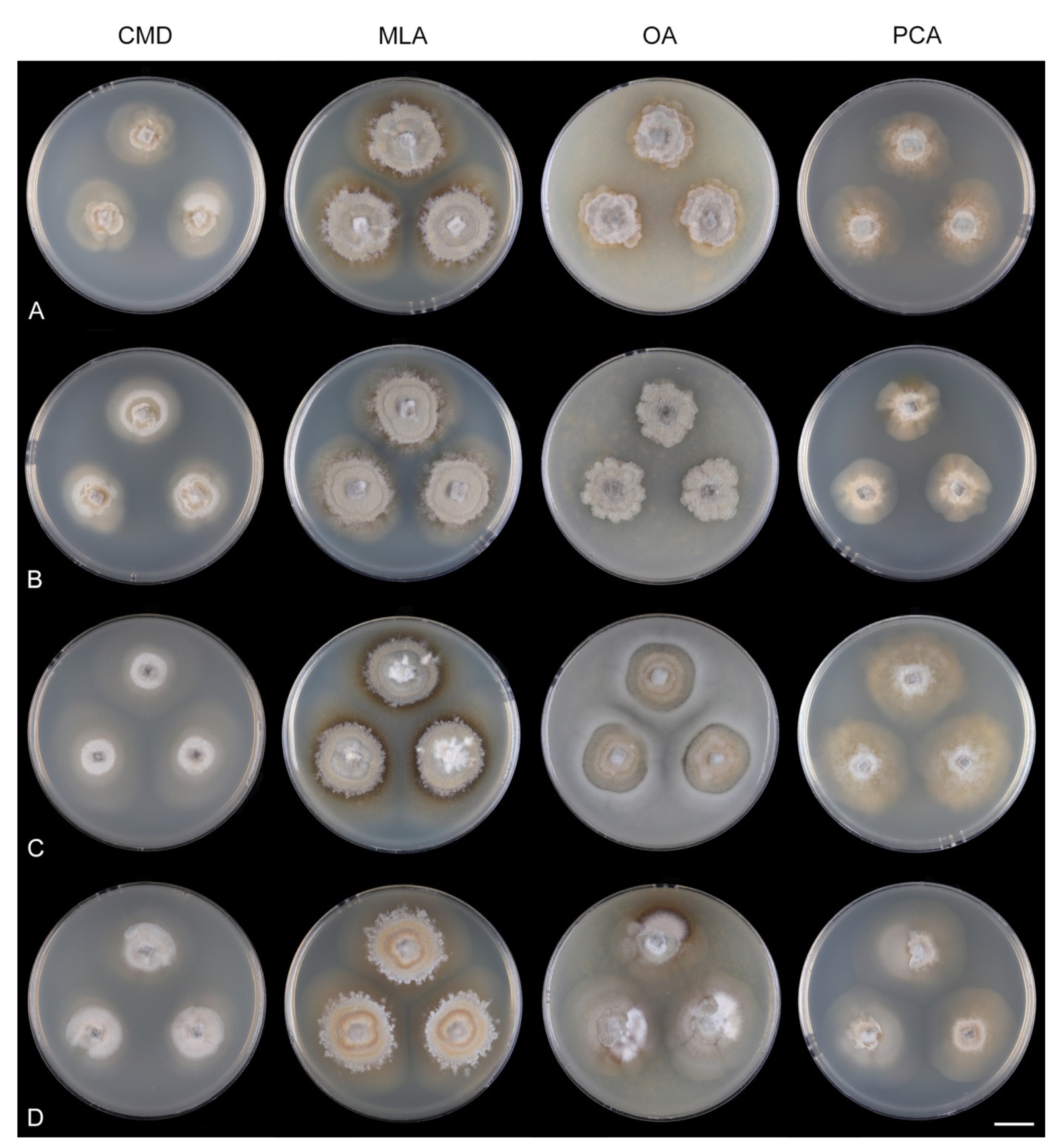{kind=link}
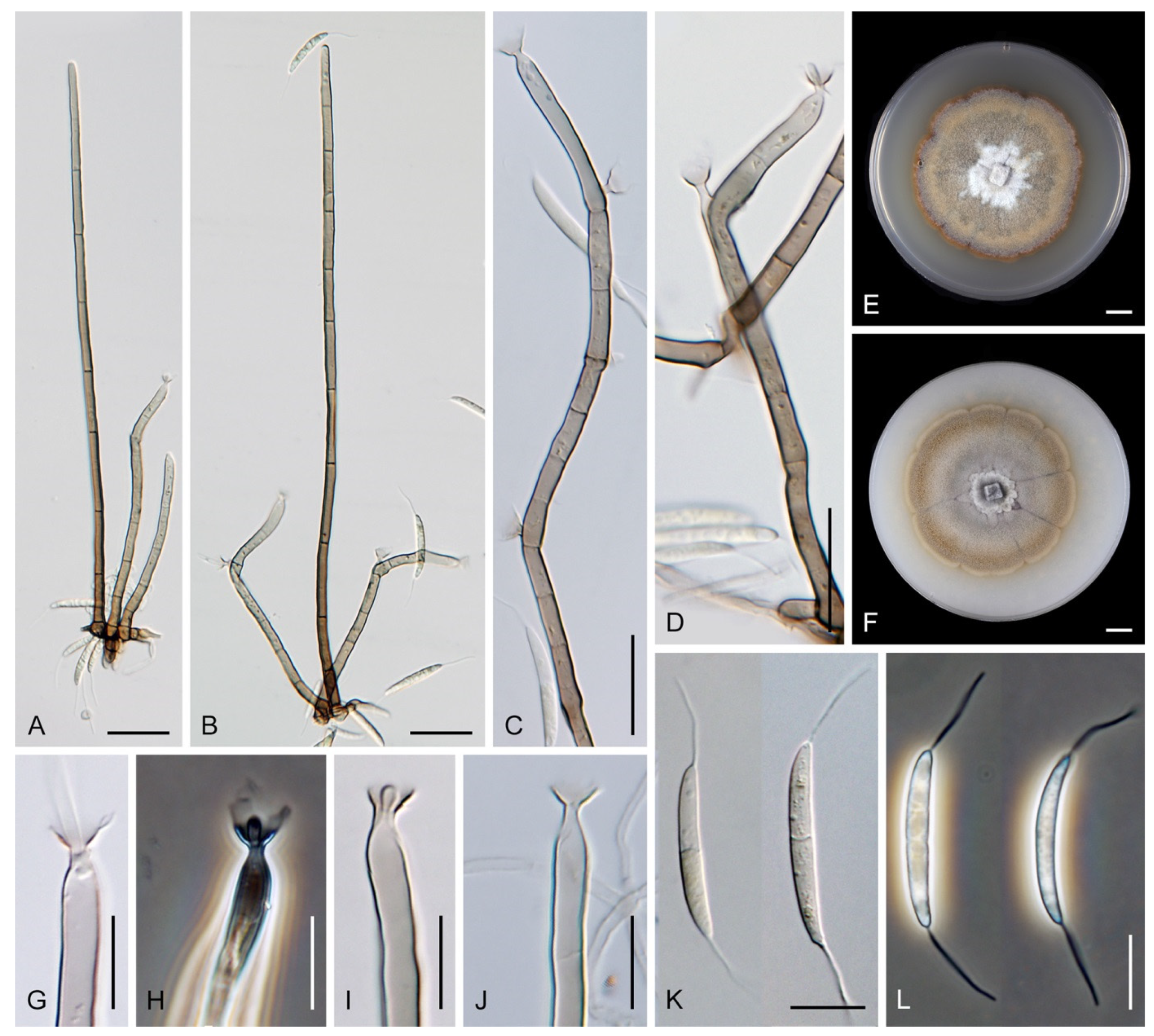{kind=link}
{kind=link}
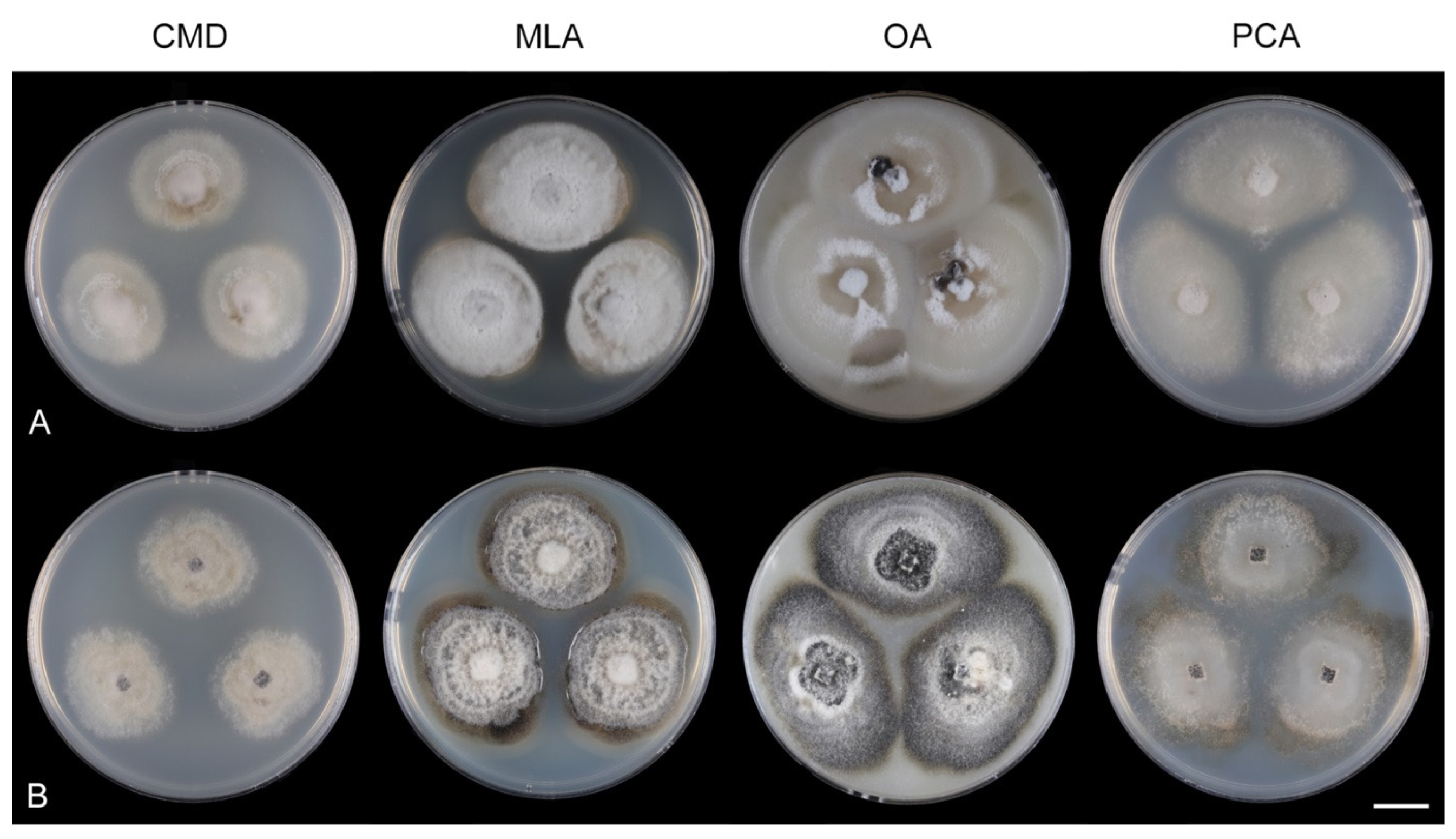{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}