
Tr-milRNA1 Contributes to Lignocellulase Secretion under Heat Stress by Regulating the Lectin-Type Cargo Receptor Gene Trvip36 in Trichoderma guizhouence NJAU 4742



Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Small RNA cDNA Library Construction and High-Throughput Sequencing
2.3. Tr-milRNAs Sequence Analysis and Target Prediction
2.4. Relative Expression of Tr-milRNA1 and the Corresponding Target Genes
2.5. Deletion and Overexpression of Fungal Small RNAs
2.6. Generation of the Target Gene Mutants
2.7. Growth and Enzyme Activity Assays of NJAU 4742
2.8. Confocal Imaging of EGL-GFP and CBH-GFP in the ER and Golgi Apparatus between wt and ΔTrvip36
3. Results
3.1. Identification and Quantification of milRNAs in NJAU 4742
3.2. Identification of Candidate Target Genes Related to Tr-milRNAs of NJAU 4742
3.3. Tr-milRNA1 Exhibits a Critical Role in Lignocellulose Utilization at Different Temperatures
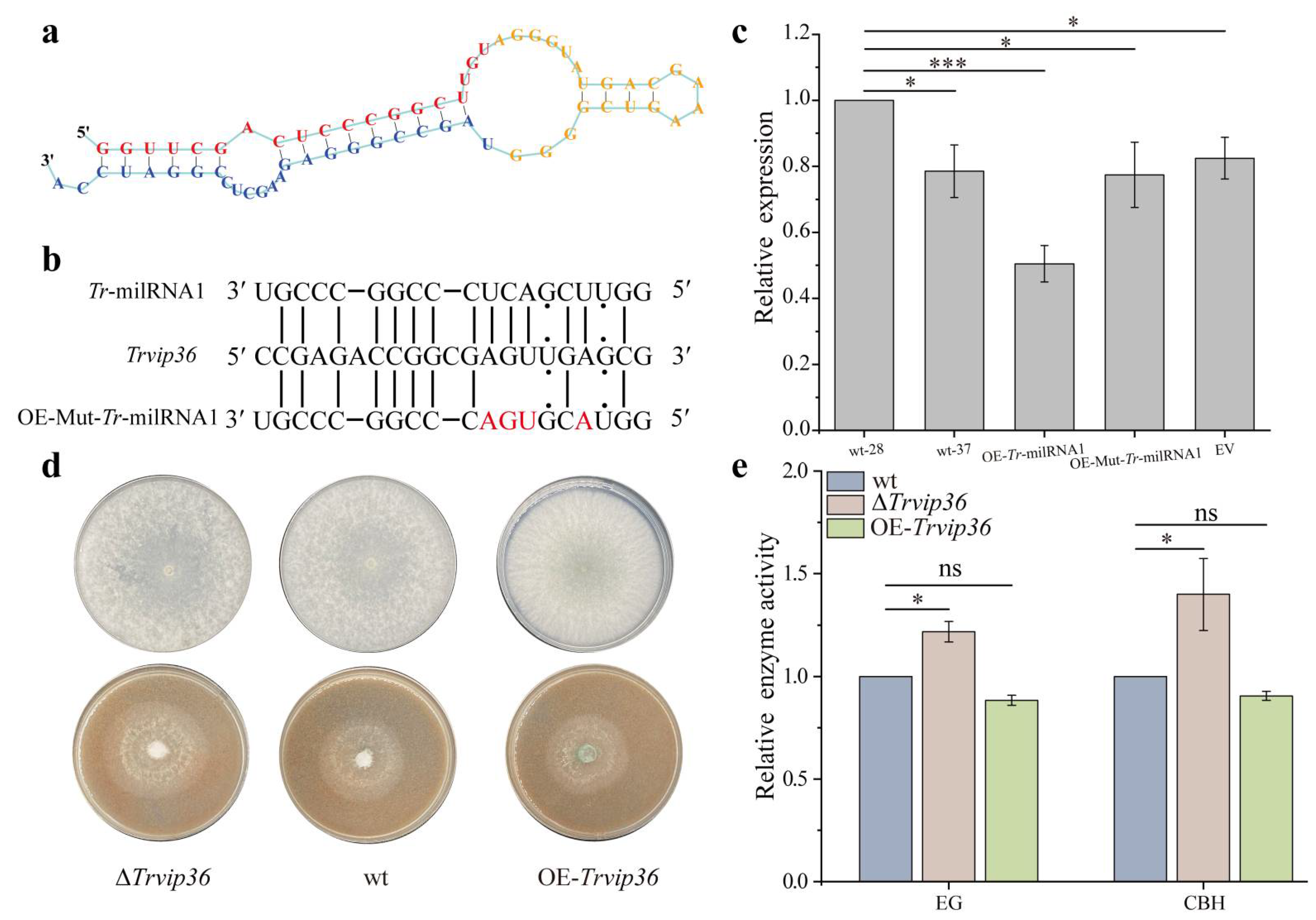
3.4. Expression of Trvip36 Could Be Regulated by Tr-milRNA1
3.5. Tr-milRNA1 Regulates Target Gene Expression in a Sequence-Specific Manner
3.6. Deletion of Trvip36 Improved the Secretion of Lignocellulases of NJAU 4742 under Heat Stress
3.7. Deletion of Trvip36 Reduces the Retention of Lignocellulases in the ER of NJAU 4742 under Heat Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chang, S.S.; Zhang, Z.; Liu, Y. RNA Interference Pathways in Fungi: Mechanisms and Functions. Annu. Rev. Microbiol. 2012, 66, 305–323. [Google Scholar] [CrossRef] [Green Version]
- Castel, S.E.; Martienssen, R.A. RNA interference in the nucleus: Roles for small RNAs in transcription, epigenetics and beyond. Nat. Rev. Genet. 2013, 14, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Holoch, D.; Moazed, D. RNA-mediated epigenetic regulation of gene expression. Nat. Rev. Genet. 2015, 16, 71–84. [Google Scholar] [CrossRef]
- Zhang, T.; Zhao, Y.L.; Zhao, J.H.; Wang, S.; Jin, Y.; Chen, Z.Q.; Fang, Y.Y.; Hua, C.L.; Ding, S.W.; Guo, H.S. Cotton plants export microRNAs to inhibit virulence gene expression in a fungal pathogen. Nat. Plants 2016, 2, 16153. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chang, S.S.; Liu, Y. RNA interference Pathways in Filamentous Fungi. Cell. Mol. Life Sci. 2010, 67, 3849–3863. [Google Scholar] [CrossRef] [Green Version]
- Romano, N.; Macino, G. Quelling: Transient inactivation of gene expression in Neurospora crassa by transformation with homologous sequences. Mol. Microbiol. 1992, 6, 3343–3353. [Google Scholar] [CrossRef]
- Verdel, A.; Moazed, D. RNAi-directed assembly of heterochromatin in Fission yeast. FEBS Lett. 2005, 579, 5872–5878. [Google Scholar] [CrossRef]
- Carthew, R.W.; Sontheimer, E.J. Origins and mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [Green Version]
- Mallory, A.C.; Vaucheret, H. Functions of microRNAs and related small RNAs in plants. Nat. Genet. 2006, 38, S31–S36. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.C.; Li, L.; Gu, W.; Xue, Z.; Yi, L. Diverse Pathways Generate MicroRNA-like RNAs and Dicer-Independent Small Interfering RNAs in Fungi. Mol. Cell 2010, 38, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Wang, Z.; Zhang, J.; Meng, H.; Huang, B. Genome-wide identification and profiling of microRNA-like RNAs from Metarhizium anisopliae during development. Fungal Biol. 2012, 116, 1156–1162. [Google Scholar] [CrossRef]
- Weiberg, A.; Wang, M.; Lin, F.M.; Zhao, H.; Zhang, Z.; Kaloshian, I.; Huang, H.D.; Jin, H. Fungal Small RNAs Suppress Plant Immunity by Hijacking Host RNA Interference Pathways. Science 2013, 342, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.; Zhong, J.; Jiang, L.; Liu, G.; Gou, C.Y.; Wu, Q.; Wang, Y.; Luo, J.; Gou, D. Identification of microRNA-Like RNAs in the filamentous fungus Trichoderma reesei by solexa sequencing. PLoS ONE 2013, 8, e76288. [Google Scholar] [CrossRef]
- Jin, Y.; Zhao, J.H.; Zhao, P.; Zhang, T.; Wang, S.; Guo, H.S. A fungal milRNA mediates epigenetic repression of a virulence gene in Verticillium dahliae. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2019, 374, 20180309. [Google Scholar] [CrossRef] [Green Version]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Mukherjee, P.K.; Zeilinger, S.; Grigoriev, I.V.; Kubicek, C.P. Trichoderma: The genomics of opportunistic success. Nat. Rev. Microbiol. 2011, 9, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Daisuke, N.; Osamu, S.; Norihito, K.; Naoki, M.; Kazuo, Y. Stable interaction of the cargo receptor VIP36 with molecular chaperone BiP. Glycobiology 2007, 17, 913–921. [Google Scholar]
- Reiterer, V.; Nyfeler, B.; Hauri, H.P. Role of the lectin VIP36 in post-ER quality control of human alpha1-antitrypsin. Traffic 2010, 11, 1044–1055. [Google Scholar] [CrossRef]
- Hoang, H.D.; Maruyama, J.I.; Kitamoto, K.; Brakhage, A. Modulating Endoplasmic Reticulum-Golgi Cargo Receptors for Improving Secretion of Carrier-Fused Heterologous Proteins in the Filamentous Fungus Aspergillus oryzae. Appl. Environ. Microbiol. 2015, 81, 533–543. [Google Scholar] [CrossRef] [Green Version]
- Miao, J.; Wang, M.; Ma, L.; Li, T.; Huang, Q.; Liu, D.; Shen, Q. Effects of amino acids on the lignocellulose degradation by Aspergillus fumigatus Z5: Insights into performance, transcriptional, and proteomic profiles. Biotechnol. Biofuels 2019, 12, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Ana, K.; Sam, G.J. miRBase: Integrating microRNA annotation and deep-sequencing data. Nucleic Acids Res. 2011, 39, D152–D157. [Google Scholar]
- Mackowiak, S.D. Identification of novel and known miRNAs in deep-sequencing data with miRDeep2. Curr. Protoc. Bioinform. 2011, 36, 12.10.1–12.10.15. [Google Scholar] [CrossRef]
- Bo, X.; Wang, S. TargetFinder: A software for antisense oligonucleotide target site selection based on MAST and secondary structures of target mRNA. Bioinformatics 2005, 21, 1401–1402. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server. Nucleic Acids Res. 2011, 39, W155–W159. [Google Scholar] [CrossRef] [Green Version]
- Varkonyi-Gasic, E.; Wu, R.; Wood, M.; Walton, E.F.; Hellens, R.P. Protocol: A highly sensitive RT-PCR method for detection and quantification of microRNAs. Plant Methods 2007, 3, 12. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Miao, Y.; Rahimi, M.J.; Zhu, H.; Steindorff, A.; Schiessler, S.; Cai, F.; Pang, G.; Chenthamara, K.; Xu, Y. Guttation capsules containing hydrogen peroxide: An evolutionarily conserved NADPH oxidase gains a role in wars between related fungi. Environ. Microbiol. 2019, 21, 2644–2658. [Google Scholar] [CrossRef] [Green Version]
- Xue, D.; Lin, D.; Gong, C.; Peng, C.; Yao, S. Expression of a bifunctional cellulase with exoglucanase and endoglucanase activities to enhance the hydrolysis ability of cellulase from a marine Aspergillus niger. Process Biochem. 2017, 52, 115–122. [Google Scholar] [CrossRef]
- Miao, Y.; Kong, Y.; Li, P.; Li, G.; Liu, D.; Shen, Q.; Zhang, R. Effect of CBM1 and linker region on enzymatic properties of a novel thermostable dimeric GH10 xylanase (Xyn10A) from filamentous fungus Aspergillus fumigatus Z5. AMB Express 2018, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Li, J.; Zhao, S.; Zhang, R.; Wang, M.; Miao, Y.; Shen, Y.; Shen, Q. Secretome diversity and quantitative analysis of cellulolytic Aspergillus fumigatus Z5 in the presence of different carbon sources. Biotechnol. Biofuels 2013, 6, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Yong, F.; Yang, Y.; Yu, Q.; Ding, M. Elaiophylin, a novel autophagy inhibitor, exerts antitumor activity as a single agent in ovarian cancer cells. Autophagy 2015, 11, 1849–1863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wi, S.G.; Cho, E.J.; Lee, D.S.; Lee, S.J.; Lee, Y.J.; Bae, H.J. Lignocellulose conversion for biofuel: A new pretreatment greatly improves downstream biocatalytic hydrolysis of various lignocellulosic materials. Biotechnol. Biofuels 2015, 8, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara-Kuge, S. Involvement of VIP36 in Intracellular Transport and Secretion of Glycoproteins in Polarized Madin-Darby Canine Kidney (MDCK) Cells. J. Biol. Chem. 2002, 277, 16332. [Google Scholar] [CrossRef] [PubMed]
- Shimada, O.; Hara-Kuge, S.; Yamashita, K.; Tosaka-Shimada, H.; Yanchao, L.; Einan, L.; Atsumi, S.; Ishikawa, H. Localization of VIP36 in the post-Golgi secretory pathway also of rat parotid acinar cells. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2003, 51, 1057–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neve, E.; Svensson, K.; Fuxe, J.; Pettersson, R.F. VIPL, a VIP36-like membrane protein with a putative function in the export of glycoproteins from the endoplasmic reticulum. Exp. Cell Res. 2003, 288, 70–83. [Google Scholar] [CrossRef]
- Lamoth, F.; Juvvadi, P.R.; Steinbach, W.J. Heat shock protein 90 (Hsp90) in fungal growth and pathogenesis. Curr. Fungal Infect. Rep. 2014, 8, 296–301. [Google Scholar] [CrossRef]
- Lamoth, F.; Juvvadi, P.R.; Fortwendel, J.R.; Steinbach, W.J. Heat shock protein 90 is required for conidiation and cell wall integrity in Aspergillus fumigatus. Eukaryot. Cell 2012, 11, 1324–1332. [Google Scholar] [CrossRef] [Green Version]
- Bhabhra, R.; Askew, D. Thermotolerance and virulence of Aspergillus fumigatus: Role of the fungal nucleolus. Med. Mycol. 2005, 43, S87–S93. [Google Scholar] [CrossRef]
- Kregel, K.C. Invited Review: Heat shock proteins: Modifying factors in physiological stress responses and acquired thermotolerance. J. Appl. Physiol. 2002, 192, 2177–2186. [Google Scholar] [CrossRef] [Green Version]
- Verghese, J.; Abrams, J.; Wang, Y.; Morano, K.A. Biology of the Heat Shock Response and Protein Chaperones: Budding Yeast (Saccharomyces cerevisiae) as a Model System. Microbiol. Mol. Biol. Rev. 2012, 76, 115–158. [Google Scholar] [CrossRef] [Green Version]
- Katiyar-Agarwal, S.; Jin, H. Role of Small RNAs in Host-Microbe Interactions. Annu. Rev. Phytopathol. 2010, 48, 225. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Castillo-Gonzalez, C.; Yu, B.; Zhang, X. The functions of plant small RNAs in development and in stress responses. Plant J. 2017, 90, 654. [Google Scholar] [CrossRef] [Green Version]
- Guan, Q.; Lu, X.; Zeng, H.; Zhang, Y.; Zhu, J. Heat stress induction of miR398 triggers a regulatory loop that is critical for thermotolerance in Arabidopsis. Plant J. 2013, 74, 840–851. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.G.; Shan, J.X.; Shi, M.; Gao, J.P.; Lin, H.X. The miR156-SPL 9-DFR pathway coordinates the relationship between development and abiotic stress tolerance in plants. Plant J. 2014, 80, 1108–1117. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Guo, Y.; Tian, R.; Gao, C.; Guo, F.; Voegele, R.T.; Bao, J.; Li, C.; Jia, C.; Feng, H. Adaptive regulation of virulence genes by microRNA-like RNAs in Valsa mali. New Phytol. 2020, 227, 899–913. [Google Scholar] [CrossRef]
- Feng, H.; Xu, M.; Gao, Y.; Liang, J.; Guo, F.; Guo, Y.; Huang, L. Vm-milR37 contributes to pathogenicity by regulating glutathione peroxidase gene VmGP in Valsa mali. Mol. Plant Pathol. 2021, 22, 243–254. [Google Scholar] [CrossRef]
- Chen, R.; Jiang, N.; Jiang, Q.; Sun, X.; Wang, Y.; Zhang, H.; Hu, Z. Exploring microRNA-like small RNAs in the filamentous fungus Fusarium oxysporum. PLoS ONE 2014, 9, e104956. [Google Scholar]
- Js, A.; Lw, A.; Yang, L.B.; Qq, B.; Bw, B.; Sl, A.; Chang, L.A. Identification of milRNAs and their target genes in Ganoderma lucidum by high-throughput sequencing and degradome analysis. Fungal Genet. Biol. 2020, 136, 103313. [Google Scholar]
- O’Keefe, S. Small Molecule Inhibitors Targeting Protein Biogenesis at the Endoplasmic Reticulum; The University of Manchester: Manchester, UK, 2019. [Google Scholar]
- Kamiya, Y.; Kato, K. Sugar Recognition by Intracellular Lectins That Determine the Fates of Glycoproteins. Trends Glycosci. Glycotechnol. 2006, 18, 231–244. [Google Scholar] [CrossRef] [Green Version]
- Tyedmers, J.; Mogk, A.; Bukau, B. Cellular strategies for controlling protein aggregation. Nat. Rev. Mol. Cell Biol. 2010, 11, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Buchberger, A.; Bukau, B.; Sommer, T. Protein quality control in the cytosol and the endoplasmic reticulum: Brothers in arms. Mol. Cell 2010, 40, 238–252. [Google Scholar] [CrossRef]
- Hara-Kuge, S.; Seko, A.; Shimada, O.; Tosaka-Shimada, H.; Yamashita, K. The binding of VIP36 and α-amylase in the secretory vesicles via high-mannose type glycans. Glycobiology 2004, 14, 739–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.; Chen, X.; Liu, Z.; Zhou, X.; Zhang, Y. Effect of inoculum sources on the anaerobic digestion of rice straw. Bioresour. Technol. 2014, 158, 149–155. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA_ID | Mirdeep2_Score | RNAfold_Results | TPM (T28) | TPM (T37) | Mature_Sequence |
|---|---|---|---|---|---|
| TGA1_S20_236772 | 2,295,965.4 | yes | 983,492.79 | 976,779.71 | uuuuugagauacuccgcaacgac |
| TGA1_S20_238860 | 31,534.5 | yes | 11,201.07 | 16,331.53 | uacguaggacuuuaccgugacgu |
| TGA1_S06_56271 | 115.5 | yes | 12.30 | 112.44 | uugccgaguggcagaggacuggcau |
| TGA1_S17_175348 | 105 | yes | 23.71 | 47.53 | uggaaguugaaucgagaagcccu |
| TGA1_S17_158248 | 85.8 | yes | 6.96 | 89.12 | gcaucugauuuccacccuuggguu |
| TGA1_S19_204554 | 43.5 | yes | 10.77 | 28.55 | agcuuuuggcuuuccagaacccgu |
| TGA1_S19_204542 | 38 | yes | 11.73 | 11.75 | uggcuggacggcccagagggccu |
| TGA1_S30_293034 | 14.6 | yes | 2.86 | 35.64 | cgcaggcucgauuguugucu |
| TGA1_S14_132594 | 13.2 | yes | 0.18 | 4.88 | cgccaacggcccucgcccggcuac |
| TGA1_S19_188258 | 12.5 | yes | 2.18 | 2.91 | caucugcauguugucucugg |
| TGA1_S10_93851 | 11.9 | yes | 0.56 | 18.67 | gucuagcacucuacuuuggcau |
| TGA1_S17_156622 | 9.3 | yes | 0.30 | 6.57 | uuuuuacacagauaccaguaggu |
| TGA1_S20_237911 | 7.5 | yes | 0.77 | 5.62 | ccaccaggccgcucaagacua |
| TGA1_S03_23817 | 7.1 | yes | 0.68 | 5.62 | cacugaccugcuccucgcacag |
| TGA1_S17_161581 | 7.1 | yes | 0 | 1.69 | caucuagcuuggacggcagcg |
| TGA1_S11_99749 | 6.5 | yes | 0.30 | 5.62 | ucgucacccuugaggcggaa |
| TGA1_S02_7777 | 5.4 | yes | 0.86 | 3.66 | auugcgaugccuggucagcuacuc |
| TGA1_S22_248536 | 5.4 | yes | 0.38 | 12.40 | ugaggagucugaagauggagaggaa |
| TGA1_S03_27309 | 5.3 | yes | 0.60 | 7.80 | guggacagauuagcugacccgcggg |
| TGA1_S15_136221 | 4.5 | yes | 0.36 | 0 | accaacagcggacauugcuccaa |
| TGA1_S11_101486 | 4 | yes | 0.60 | 0 | ugcuccaaaugagaaucgaagu |
| TGA1_S11_106603 | 3.7 | yes | 0.76 | 0 | ugccugguacauguacgga |
| TGA1_S07_71495 | 3.5 | yes | 1.80 | 0.85 | ugaccaagaacuucgacgucuu |
| TGA1_S15_142991 | 3.3 | yes | 0 | 2.54 | uuuggaccgugucuggaacgcua |
| TGA1_S17_178322 | 3.3 | yes | 0 | 5.62 | ucauauucucagcacuuggaau |
| TGA1_S02_16357 | 3.2 | yes | 0 | 1.70 | agguaacgucugguggcaa |
| TGA1_S05_35063 | 2.9 | yes | 0 | 4.98 | gcaagaucaaaacucaaa |
| TGA1_S17_175238 | 2.8 | yes | 4.34 | 13.62 | uggaaguugaaucgagaagccc |
| TGA1_S09_88615 | 2.7 | yes | 0 | 1.70 | cguguaccagagcgucau |
| TGA1_S08_81282 | 2.5 | yes | 0.60 | 0 | uccgauagggauguucgggcu |
| TGA1_S20_238862 | 2.5 | yes | 4634.89 | 4921.04 | uaggacuuuaccgugacgucc |
| TGA1_S22_257134 | 2.5 | yes | 1.45 | 12.20 | cgguggauugaacgggacucuuggu |
| TGA1_S05_43489 | 2.4 | yes | 28.74 | 491.72 | cucgggagaaggcggccu |
| TGA1_S07_78907 | 2.3 | yes | 0.36 | 0 | gucuucuugaucucuuau |
| TGA1_S18_185930 | 2.3 | yes | 25.59 | 160.54 | acccggcuuggagaugug |
| TGA1_S20_209761 | 2.3 | yes | 0.38 | 0 | ugaugauucggcuaguucggacag |
| TGA1_S22_248446 | 2.3 | yes | 0.94 | 10.71 | cggcgaggcuguguuucagcga |
| TGA1_S20_223477 | 2.2 | yes | 445.56 | 655.43 | uacguaggacucuaccgugacau |
| TGA1_S20_238808 | 2.2 | yes | 44.36 | 38.21 | guucgaggguugaaauga |
| TGA1_S06_55136 | 2 | yes | 0 | 1.70 | auguaugccuccugagauua |
| TGA1_S16_151294 | 2 | yes | 0 | 1.70 | acuucauccauagauaucgcaa |
| TGA1_S17_162471 | 2 | yes | 3.95 | 13.62 | uggaaguugaaucgagaagccc |
| TGA1_S19_199524 | 1.9 | yes | 0.18 | 0 | uaccucucauucacugcagug |
| TGA1_S02_9056 | 1.8 | yes | 1.77 | 26.56 | ggaaacaagguuguucugacuau |
| TGA1_S13_112804 | 1.8 | yes | 0.60 | 0 | uuaugugguacggcagagagu |
| TGA1_S04_31828 | 1.7 | yes | 0.30 | 23.35 | gguucgacucccggcuugu |
| TGA1_S14_132761 | 1.6 | yes | 7.96 | 6.95 | aacgugcaauugcuaccaa |
| TGA1_S20_229688 | 1.6 | yes | 2.62 | 5.62 | ugagggaccggauucgcca |
| TGA1_S22_253299 | 1.6 | yes | 15.54 | 38.89 | auccaaaagcucggcuuu |
| TGA1_S23_271212 | 1.5 | yes | 0 | 5.62 | agcgaggacauuuaugac |
| TGA1_S05_33550 | 1.4 | yes | 2.47 | 27.03 | accuagagaacgaugguucccauu |
| TGA1_S07_67387 | 1.3 | yes | 0.71 | 0 | aagcagauugcgaggggucauuug |
| TGA1_S22_265703 | 1.1 | yes | 0.38 | 9.86 | gggcagucuguuggacuccggu |
| TGA1_S07_76476 | 1 | yes | 1.14 | 0.85 | cucggucuguuguggauugguc |
| TGA1_S19_189543 | 0.7 | yes | 1.14 | 0 | aaggacauuuuggaggag |
| TGA1_S07_65040 | 0.1 | yes | 1.50 | 5.73 | ucgaaguugugguuguagugguagu |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, T.; Liu, J.; Wang, Q.; Liu, Y.; Li, T.; Liu, D.; Shen, Q. Tr-milRNA1 Contributes to Lignocellulase Secretion under Heat Stress by Regulating the Lectin-Type Cargo Receptor Gene Trvip36 in Trichoderma guizhouence NJAU 4742. J. Fungi 2021, 7, 997. https://doi.org/10.3390/jof7120997
Li T, Liu J, Wang Q, Liu Y, Li T, Liu D, Shen Q. Tr-milRNA1 Contributes to Lignocellulase Secretion under Heat Stress by Regulating the Lectin-Type Cargo Receptor Gene Trvip36 in Trichoderma guizhouence NJAU 4742. Journal of Fungi. 2021; 7(12):997. https://doi.org/10.3390/jof7120997
Chicago/Turabian StyleLi, Tuo, Jinding Liu, Qin Wang, Yang Liu, Ting Li, Dongyang Liu, and Qirong Shen. 2021. "Tr-milRNA1 Contributes to Lignocellulase Secretion under Heat Stress by Regulating the Lectin-Type Cargo Receptor Gene Trvip36 in Trichoderma guizhouence NJAU 4742" Journal of Fungi 7, no. 12: 997. https://doi.org/10.3390/jof7120997
APA StyleLi, T., Liu, J., Wang, Q., Liu, Y., Li, T., Liu, D., & Shen, Q. (2021). Tr-milRNA1 Contributes to Lignocellulase Secretion under Heat Stress by Regulating the Lectin-Type Cargo Receptor Gene Trvip36 in Trichoderma guizhouence NJAU 4742. Journal of Fungi, 7(12), 997. https://doi.org/10.3390/jof7120997