
Natural Killer Cell Line NK-92-Mediated Damage of Medically Important Fungi


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. NK-92 Cell Cultivation
2.2. Assessment of the Antifungal Activity of NK-92 Cells
2.3. Preparation of the Fungi
2.4. Assessment of the Degranulation of NK-92 Cells
2.5. Assessment of the Concentration of Soluble Molecules in the Supernatant
2.6. Statistical Analyses
3. Results
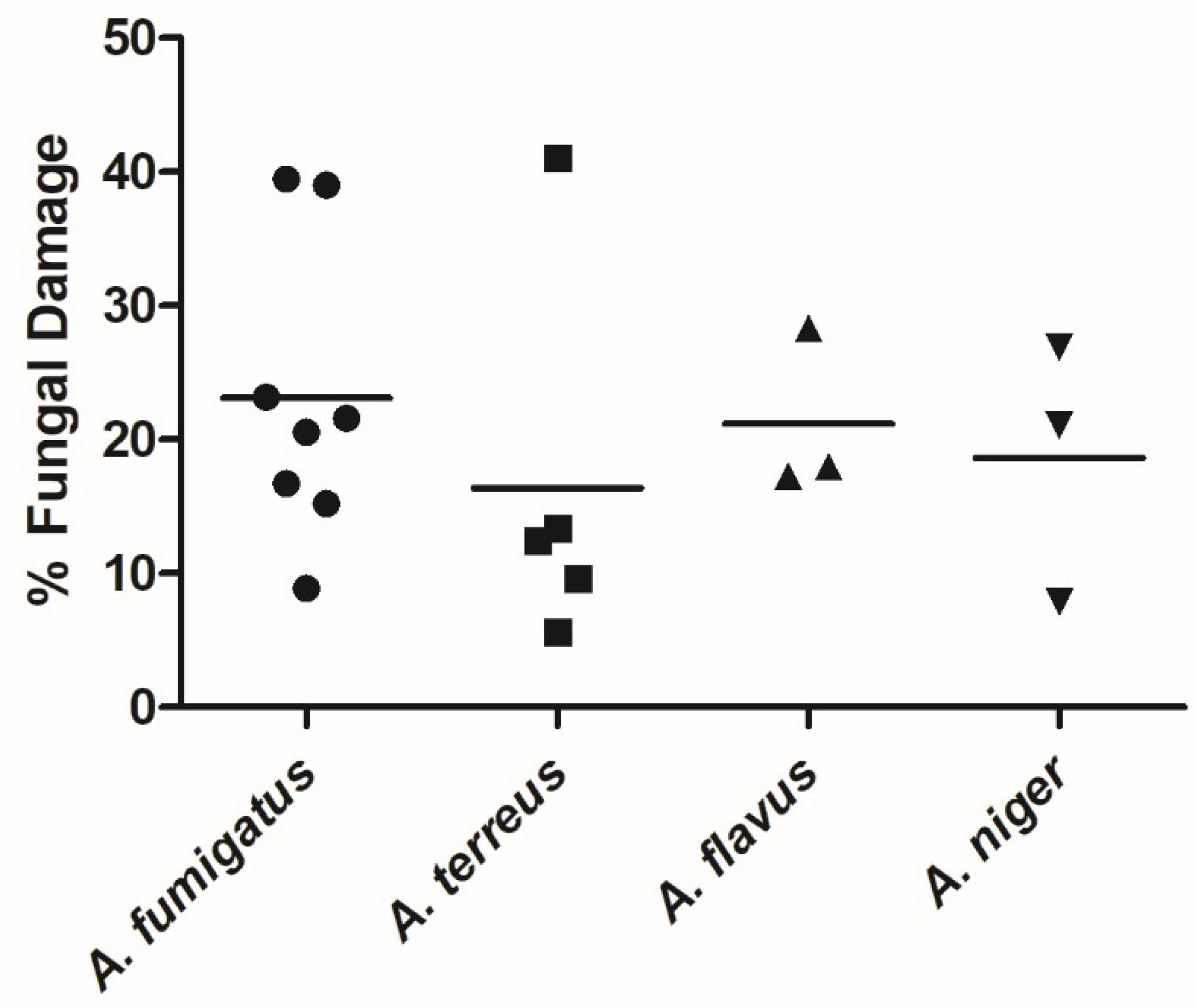
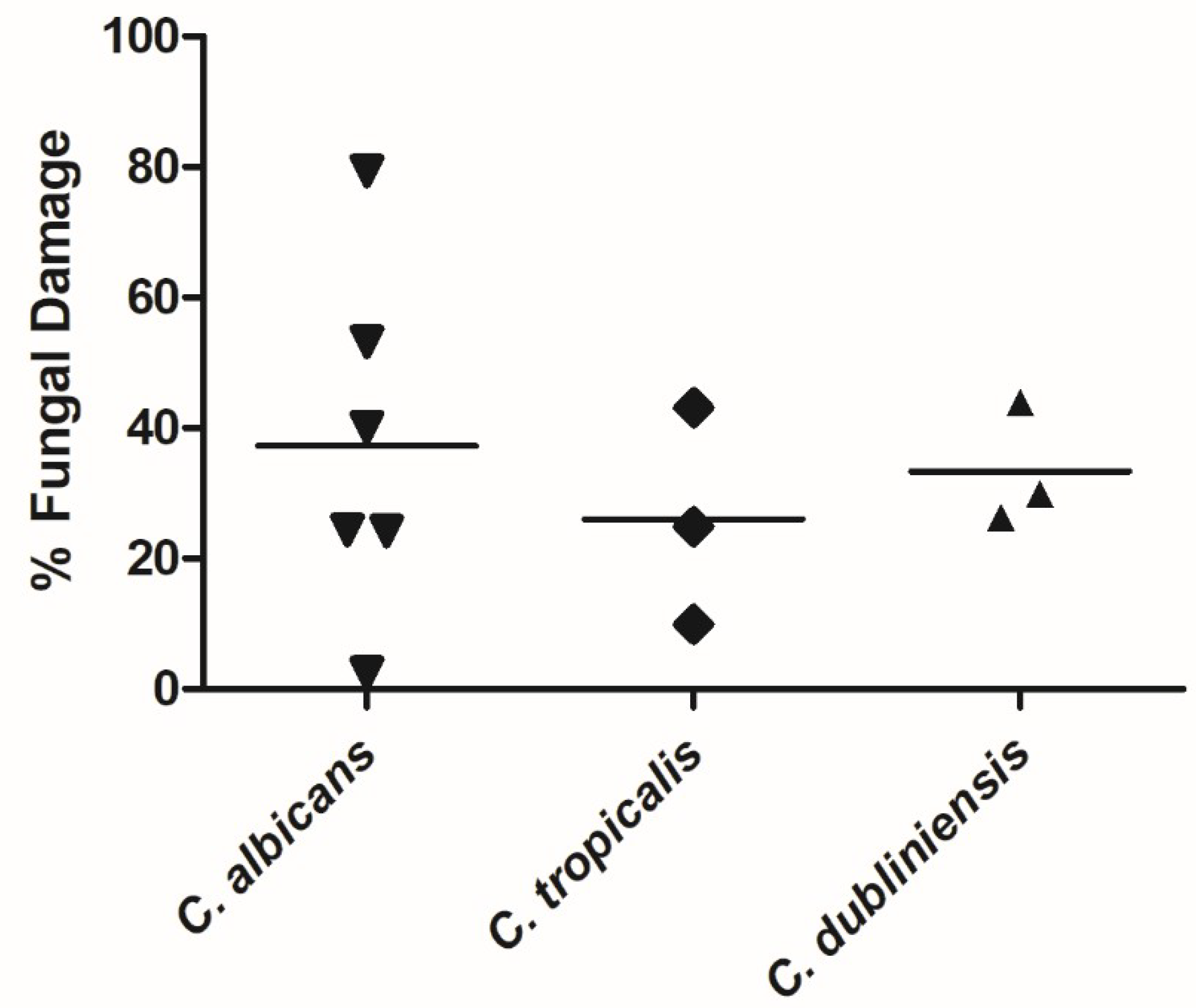
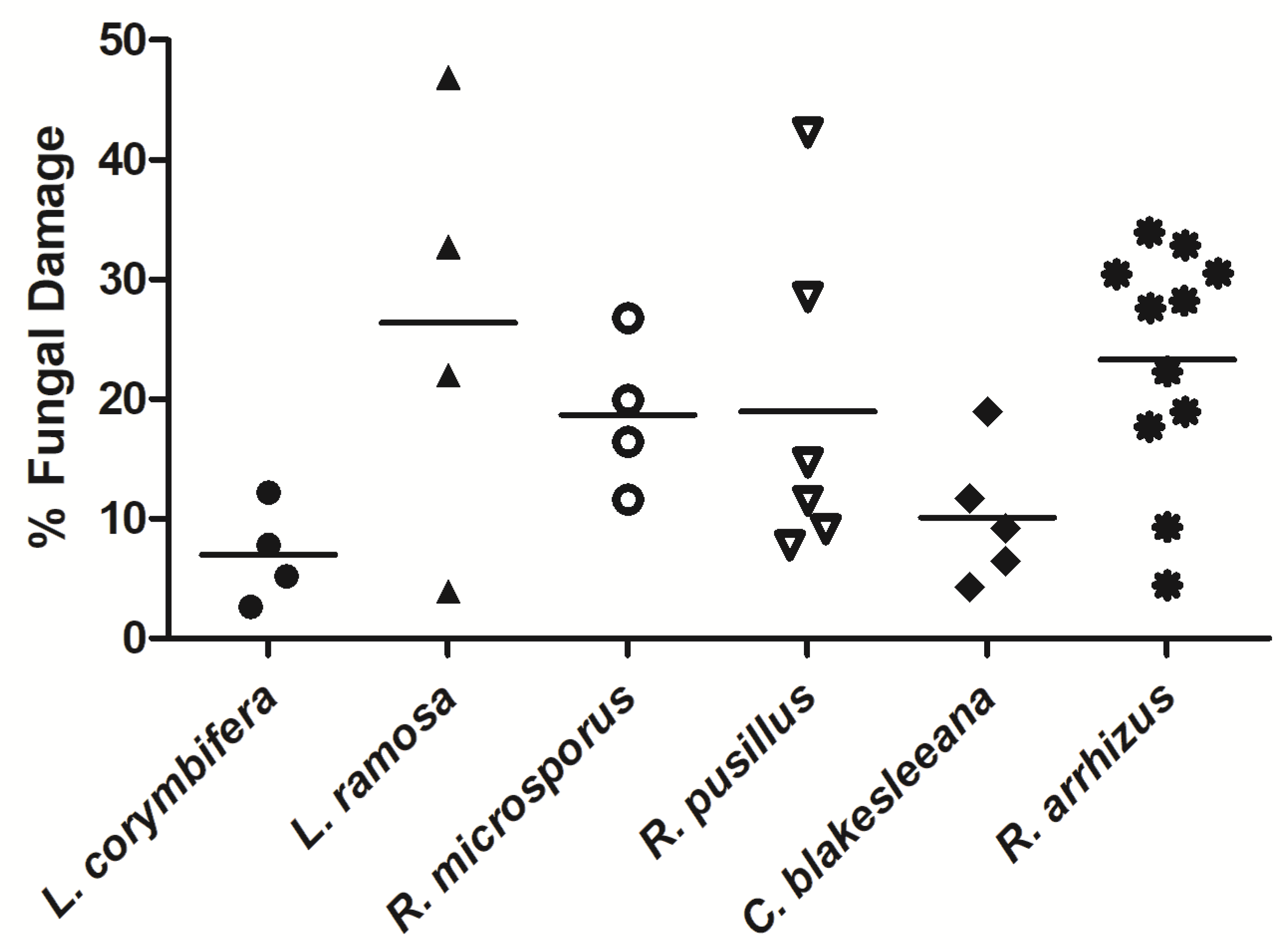
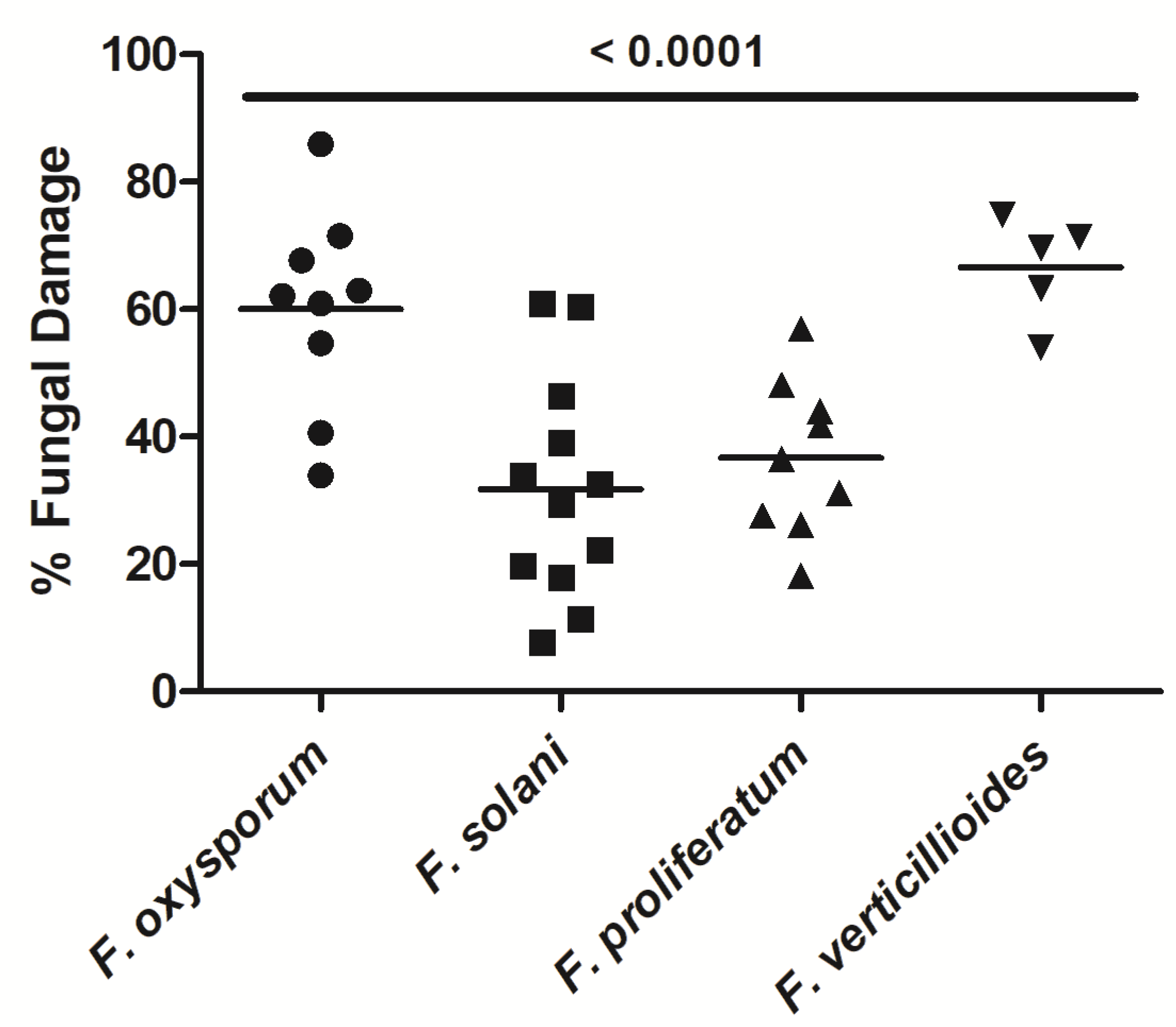
3.1. NK-92 Cells Damaged Hyphae of a Variety of Medically Important Fungi
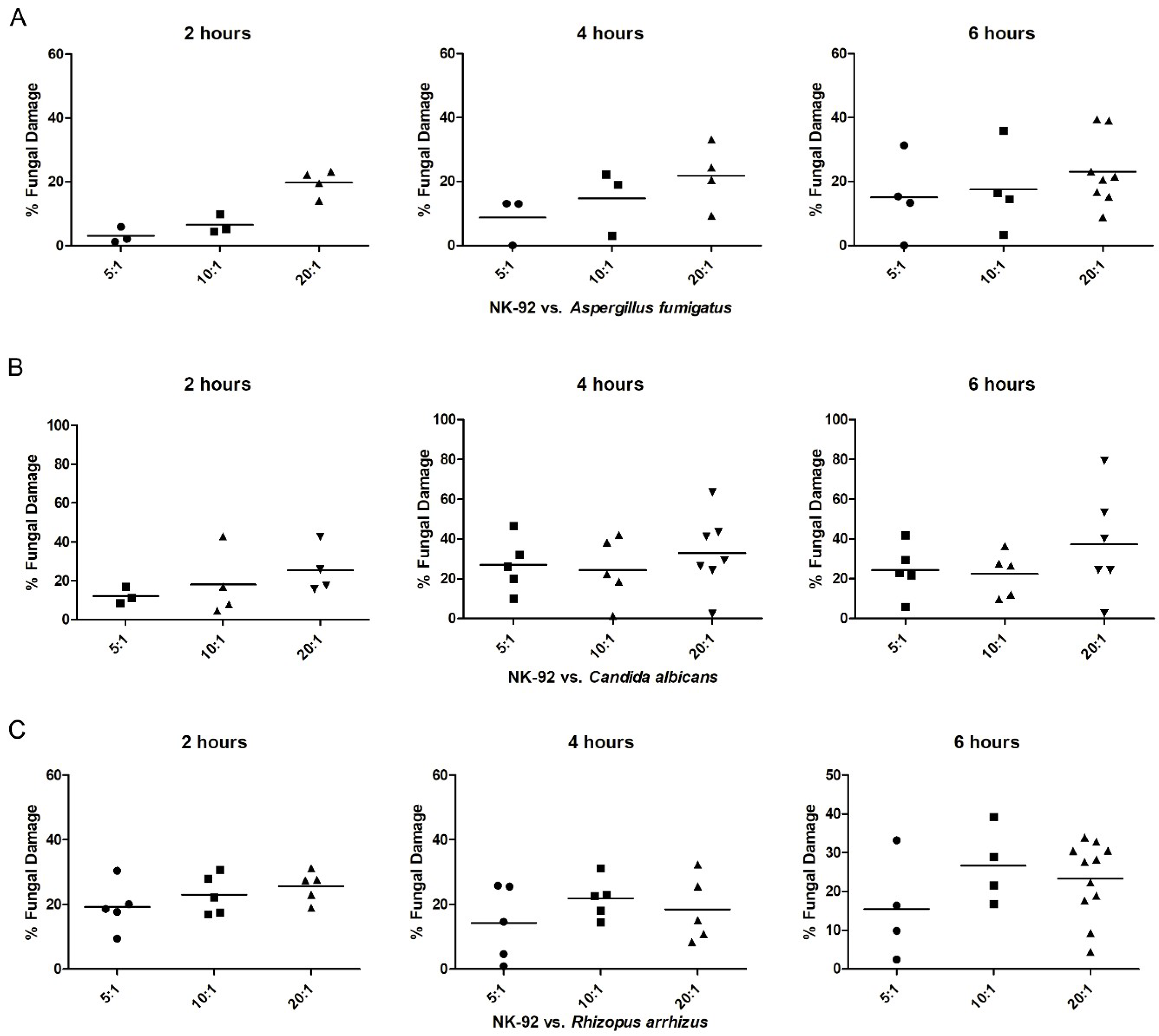
3.2. The Extent of Fungal Damage of Aspergillus fumigatus, Candida flbicans, and Rhizopus frrhizus Depended on the Co-Incubation Period and Effector/Target Ratio
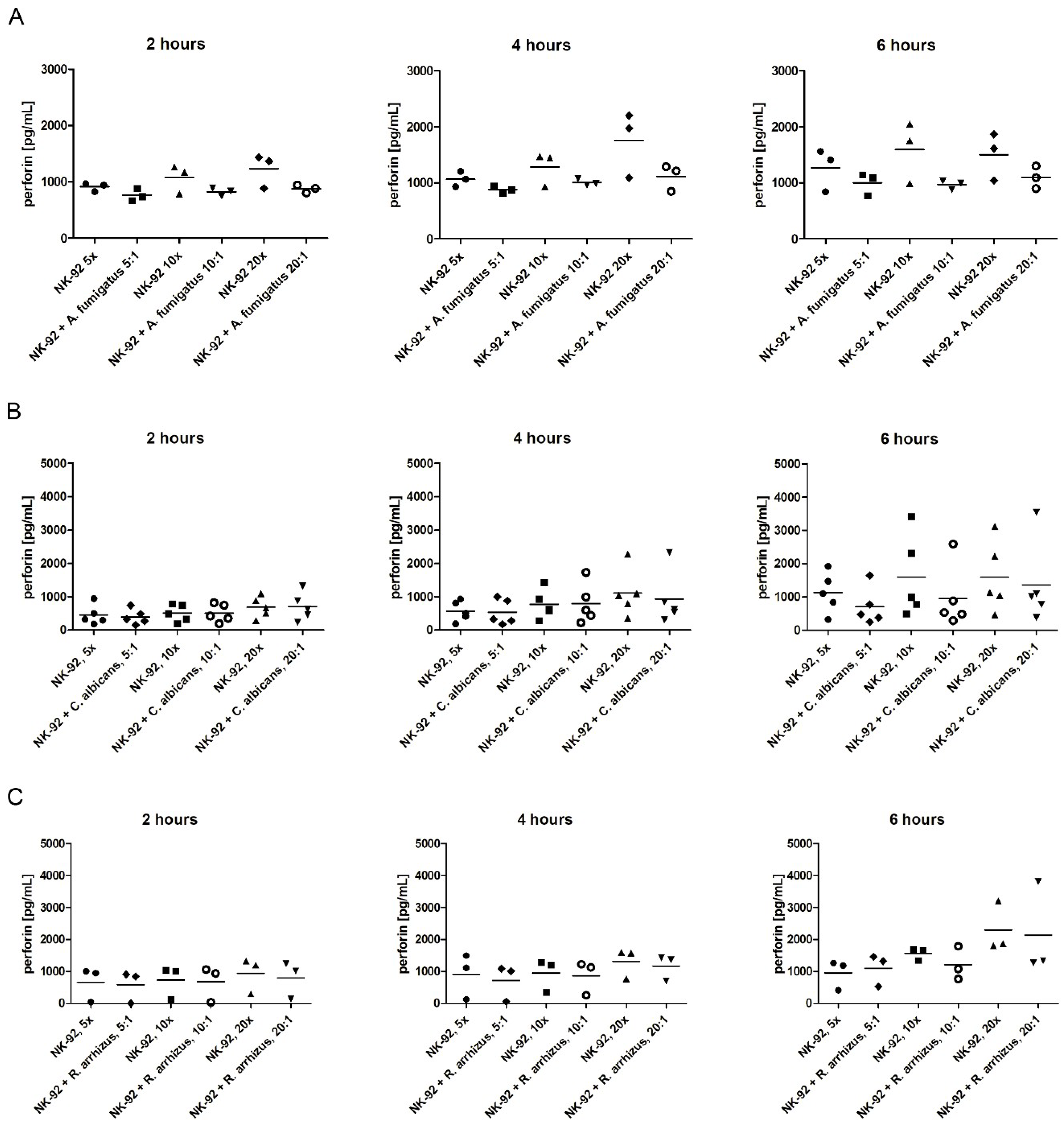
3.3. Assessment of Perforin Levels in the Supernatant and of the Degranulation Marker CD107a on the Surface of NK-92 Cells
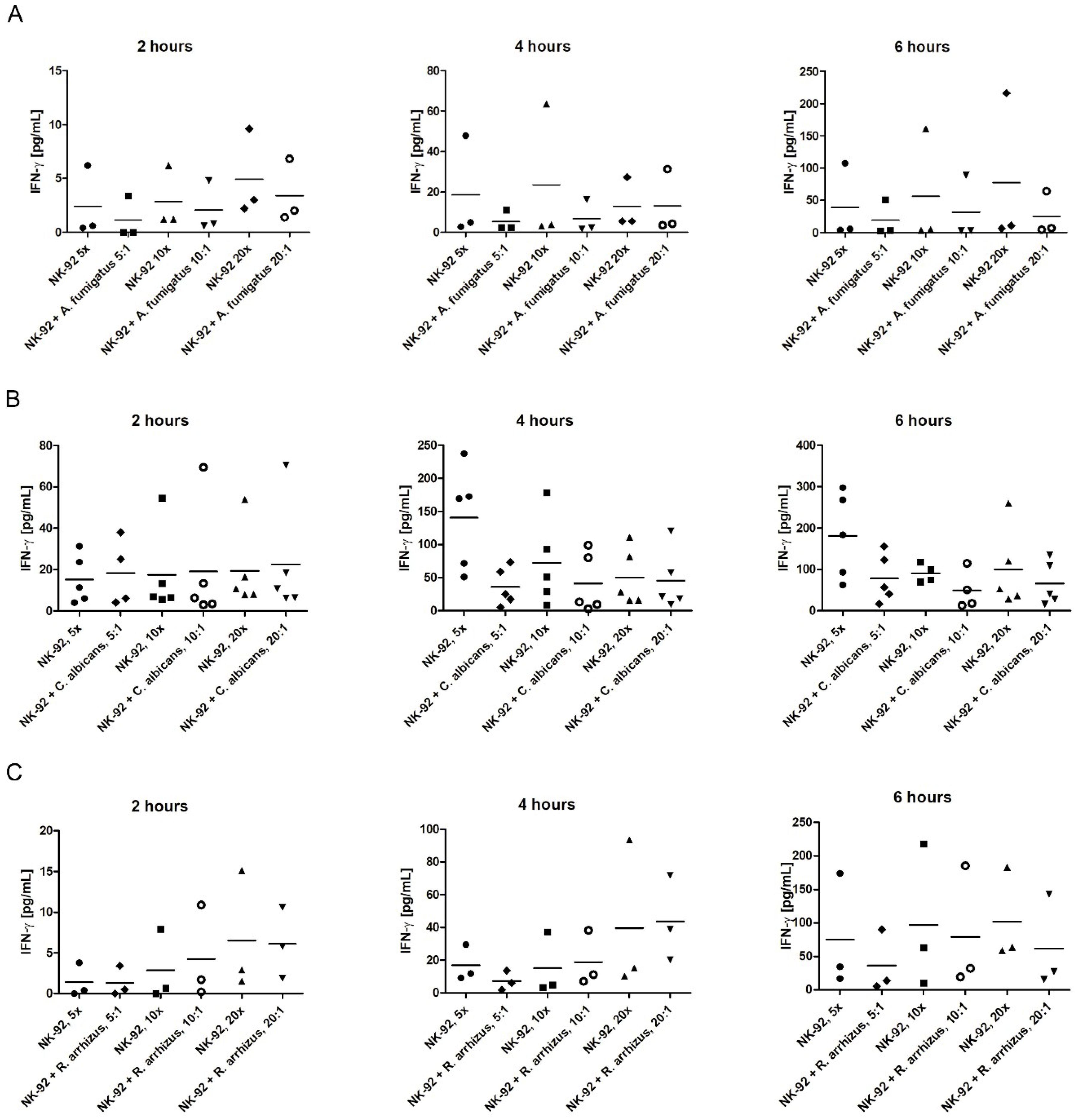
3.4. Assessment of IFN-γ Levels in the Supernatant
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Montesinos, P.; Rodríguez-Veiga, R.; Boluda, B.; Martínez-Cuadrón, D.; Cano, I.; Lancharro, A.; Sanz, J.; Arilla, M.J.; López-Chuliá, F.; Navarro, I.; et al. Incidence and risk factors of post-engraftment invasive fungal disease in adult allogeneic hematopoietic stem cell transplant recipients receiving oral azoles prophylaxis. Bone Marrow Transplant. 2015, 50, 1465–1472. [Google Scholar] [CrossRef] [Green Version]
- Lehrnbecher, T.; Schöning, S.; Poyer, F.; Georg, J.; Becker, A.; Gordon, K.; Attarbaschi, A.; Groll, A.H. Incidence and Outcome of Invasive Fungal Diseases in Children With Hematological Malignancies and/or Allogeneic Hematopoietic Stem Cell Transplantation: Results of a Prospective Multicenter Study. Front. Microbiol. 2019, 10, 681. [Google Scholar] [CrossRef]
- Neofytos, D.; Horn, D.; Anaissie, E.; Steinbach, W.; Olyaei, A.; Fishman, J.; Pfaller, M.; Chang, C.; Webster, K.; Marr, K. Epidemiology and outcome of invasive fungal infection in adult hematopoietic stem cell transplant recipients: Analysis of multicenter prospective antifungal therapy (PATH) alliance registry. Clin. Infect. Dis. 2009, 48, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Stanzani, M.; Lewis, R. Development and Applications of Prognostic Risk Models in the Management of Invasive Mold Disease. J. Fungi 2018, 4, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drgona, L.; Khachatryan, A.; Stephens, J.; Charbonneau, C.; Kantecki, M.; Haider, S.; Barnes, R. Clinical and economic burden of invasive fungal diseases in Europe: Focus on pre-emptive and empirical treatment of Aspergillus and Candida species. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 7–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuehler, C.; Kuenzli, E.; Jaeger, V.K.; Baettig, V.; Ferracin, F.; Rajacic, Z.; Kaiser, D.; Bernardini, C.; Forrer, P.; Weisser, M.; et al. Immune Reconstitution After Allogeneic Hematopoietic Stem Cell Transplantation and Association With Occurrence and Outcome of Invasive Aspergillosis. J. Infect. Dis. 2015, 212, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Fisher, B.T.; Robinson, P.D.; Lehrnbecher, T.; Steinbach, W.J.; Zaoutis, T.E.; Phillips, B.; Sung, L. Risk Factors for Invasive Fungal Disease in Pediatric Cancer and Hematopoietic Stem Cell Transplantation: A Systematic Review. J. Pediatric Infect. Dis. Soc. 2018, 7, 191–198. [Google Scholar] [CrossRef]
- Hebart, H.; Bollinger, C.; Fisch, P.; Sarfati, J.; Meisner, C.; Baur, M.; Loeffler, J.; Monod, M.; Latgé, J.P.; Einsele, H. Analysis of T-cell responses to Aspergillus fumigatus antigens in healthy individuals and patients with hematologic malignancies. Blood 2002, 100, 4521–4528. [Google Scholar] [CrossRef]
- Seidel, M.G.; Peters, C.; Wacker, A.; Northoff, H.; Moog, R.; Boehme, A.; Silling, G.; Grimminger, W.; Einsele, H. Randomized phase III study of granulocyte transfusions in neutropenic patients. Bone Marrow Transplant. 2008, 42, 679–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo, P.; Wright, K.E.; Kontoyiannis, D.P.; Walsh, T.; Patel, S.; Chorvinsky, E.; Bose, S.; Hazrat, Y.; Omer, B.; Albert, N.; et al. A New Method for Reactivating and Expanding T Cells Specific for Rhizopus oryzae. Mol. Ther. Methods Clin. Dev. 2018, 9, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Tramsen, L.; Schmidt, S.; Boenig, H.; Latgé, J.P.; Lass-Flörl, C.; Roeger, F.; Seifried, E.; Klingebiel, T.; Lehrnbecher, T. Clinical-scale generation of multi-specific anti-fungal T cells targeting Candida, Aspergillus and mucormycetes. Cytotherapy 2013, 15, 344–351. [Google Scholar] [CrossRef]
- Yan, Y.; Steinherz, P.; Klingemann, H.G.; Dennig, D.; Childs, B.H.; McGuirk, J.; O’Reilly, R.J. Antileukemia activity of a natural killer cell line against human leukemias. Clin. Cancer Res. 1998, 4, 2859–2868. [Google Scholar] [PubMed]
- Schmidt, S.; Tramsen, L.; Rais, B.; Ullrich, E.; Lehrnbecher, T. Natural killer cells as a therapeutic tool for infectious diseases—Current status and future perspectives. Oncotarget 2018, 9, 20891–20907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.J.; Hughes, M.A.; Burdick, M.; Strieter, R.M.; Mehrad, B. Early NK cell-derived IFN-γ is essential to host defense in neutropenic invasive aspergillosis. J. Immunol. 2009, 182, 4306–4312. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Ruiz, M.; López-Medrano, F.; San Juan, R.; Allende, L.M.; Paz-Artal, E.; Aguado, J.M. Low Natural Killer Cell Counts and Onset of Invasive Fungal Disease After Solid Organ Transplantation. J. Infect. Dis. 2016, 213, 873–874. [Google Scholar] [CrossRef]
- Soe, W.M.; Hui, J.; Lim, J.; Williams, D.L.; Goh, J.G.; Tan, Z.; Sam, Q.H.; Chotirmall, S.H.; A’tikah Binte, N.; Ali, M.; et al. Using Expanded Natural Killer Cells as Therapy for Invasive Aspergillosis. J. Fungi 2020, 6, 231. [Google Scholar] [CrossRef] [PubMed]
- Tonn, T.; Becker, S.; Esser, R.; Schwabe, D.; Seifried, E. Cellular Immunotherapy of Malignancies Using the Clonal Natural Killer Cell Line NK-92. J. Hematother. Stem Cell Res. 2001, 10, 535–544. [Google Scholar] [CrossRef]
- Tam, Y.K.; Miyagawa, B.; Ho, V.C.; Klingemann, H.G. Immunotherapy of malignant melanoma in a SCID mouse model using the highly cytotoxic natural killer cell line NK-92. J. Hematother. 1999, 8, 281–290. [Google Scholar] [CrossRef]
- Gong, J.H.; Maki, G.; Klingemann, H.G. Characterization of a human cell line (NK-92) with phenotypical and functional characteristics of activated natural killer cells. Leukemia 1994, 8, 652–658. [Google Scholar]
- Klingemann, H.G.; Wong, E.; Maki, G. A cytotoxic NK-cell line (NK-92) for ex vivo purging of leukemia from blood. Biol. Blood Marrow Transplant. 1996, 2, 68–75. [Google Scholar]
- Tonn, T.; Schwabe, D.; Klingemann, H.G.; Becker, S.; Esser, R.; Koehl, U.; Suttorp, M.; Seifried, E.; Ottmann, O.G.; Bug, G. Treatment of patients with advanced cancer with the natural killer cell line NK-92. Cytotherapy 2013, 15, 1563–1570. [Google Scholar] [CrossRef]
- Arai, S.; Meagher, R.; Swearingen, M.; Myint, H.; Rich, E.; Martinson, J.; Klingemann, H. Infusion of the allogeneic cell line NK-92 in patients with advanced renal cell cancer or melanoma: A phase I trial. Cytotherapy 2008, 10, 625–632. [Google Scholar] [CrossRef]
- Boyiadzis, M.; Agha, M.; Redner, R.L.; Sehgal, A.; Im, A.; Hou, J.Z.; Farah, R.; Dorritie, K.A.; Raptis, A.; Lim, S.H.; et al. Phase 1 clinical trial of adoptive immunotherapy using “off-the-shelf” activated natural killer cells in patients with refractory and relapsed acute myeloid leukemia. Cytotherapy 2017, 19, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Tramsen, L.; Hanisch, M.; Latgé, J.-P.; Huenecke, S.; Koehl, U.; Lehrnbecher, T. Human natural killer cells exhibit direct activity against Aspergillus fumigatus hyphae, but not against resting conidia. J. Infect. Dis. 2011, 203, 430–435. [Google Scholar] [CrossRef] [Green Version]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal Identification Using Molecular Tools: A Primer for the Natural Products Research Community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Tramsen, L.; Perkhofer, S.; Lass-Flörl, C.; Hanisch, M.; Roger, F.; Klingebiel, T.; Koehl, U.; Lehrnbecher, T. Rhizopus oryzae hyphae are damaged by human natural killer (NK) cells, but suppress NK cell mediated immunity. Immunobiology 2013, 218, 939–944. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Schneider, A.; Demir, A.; Lass-Flörl, C.; Lehrnbecher, T. Natural killer cell-mediated damage of clinical isolates of mucormycetes. Mycoses 2016, 59, 34–38. [Google Scholar] [CrossRef]
- Aktas, E.; Kucuksezer, U.C.; Bilgic, S.; Erten, G.; Deniz, G. Relationship between CD107a expression and cytotoxic activity. Cell. Immunol. 2009, 254, 149–154. [Google Scholar] [CrossRef]
- Zhang, T.; Li, Y.; Wang, Y.-J.; Wang, X.; Young, M.; Douglas, S.D.; Ho, W.-Z. Natural killer cell inhibits human immunodeficiency virus replication in chronically infected immune cells. Antiviral Res. 2007, 73, 132–139. [Google Scholar] [CrossRef]
- Iversen, A.; Norris, P.S.; Ware, C.F.; Benedict, C.A. Human NK cells inhibit cytomegalovirus replication through a noncytolytic mechanism involving lymphotoxin-dependent induction of IFN-beta. J. Immunol. 2005, 175, 7568–7574. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, T.; Ho, C.; Orange, J.S.; Douglas, S.D.; Ho, W.-Z. Natural killer cells inhibit hepatitis C virus expression. J. Leukoc. Biol. 2004, 76, 1171–1179. [Google Scholar] [CrossRef]
- Schmidt, S.; Schubert, R.; Tramsen, L.; Lehrnbecher, T. Impact of antifungal compounds on viability and anti-Aspergillus activity of human natural killer cells. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [Green Version]
- Antachopoulos, C.; Demchok, J.P.; Roilides, E.; Walsh, T.J. Fungal biomass is a key factor affecting polymorphonuclear leucocyte-induced hyphal damage of filamentous fungi. Mycoses 2009, 53, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Voigt, J.; Hünniger, K.; Bouzani, M.; Jacobsen, I.D.; Barz, D.; Hube, B.; Löffler, J.; Kurzai, O. Human Natural Killer Cells Acting as Phagocytes Against Candida albicans and Mounting an Inflammatory Response That Modulates Neutrophil Antifungal Activity. J. Infect. Dis. 2014, 209, 616–626. [Google Scholar] [CrossRef] [Green Version]
- Hellwig, D.; Voigt, J.; Bouzani, M.; Löffler, J.; Albrecht-Eckardt, D.; Weber, M.; Brunke, S.; Martin, R.; Kurzai, O.; Hünniger, K. Candida albicans Induces Metabolic Reprogramming in Human NK Cells and Responds to Perforin with a Zinc Depletion Response. Front. Microbiol. 2016, 7, 750. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.L.; Wang, C.L.C.; Neely, G.G.; Epelman, S.; Krensky, A.M.; Mody, C.H. NK cells use perforin rather than granulysin for anticryptococcal activity. J. Immunol. 2004, 173, 3357–3365. [Google Scholar] [CrossRef] [Green Version]
- Schneider, A.; Blatzer, M.; Posch, W.; Schubert, R.; Lass-Flörl, C.; Schmidt, S.; Lehrnbecher, T. Aspergillus fumigatus responds to natural killer (NK) cells with upregulation of stress related genes and inhibits the immunoregulatory function of NK cells. Oncotarget 2016, 7, 71062–71071. [Google Scholar] [CrossRef]
- Gil-lamaignere, C.; Simitsopoulou, M.; Roilides, E.; Maloukou, A.; Winn, R.M.; Walsh, T.J. Interferon-γ and Granulocyte-Macrophage Colony-Stimulating Factor Augment the Activity of Polymorphonuclear Leukocytes against Medically Important Zygomycetes. J. Infect. Dis. 2005, 191, 1180–1187. [Google Scholar] [CrossRef] [Green Version]
- Lehrnbecher, T.; Kalkum, M.; Champer, J.; Tramsen, L.; Schmidt, S.; Klingebiel, T. Immunotherapy in Invasive Fungal Infection—Focus on Invasive Aspergillosis. Curr. Pharm. Des. 2013, 19, 3689–3712. [Google Scholar] [CrossRef] [PubMed]
- Uicker, W.C.; Doyle, H.A.; McCracken, J.P.; Langlois, M.; Buchanan, K.L. Cytokine and chemokine expression in the central nervous system associated with protective cell-mediated immunity against Cryptococcus neoformans. Med. Mycol. 2005, 43, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Santiago, V.; Rezvani, K.; Sekine, T.; Stebbing, J.; Kelleher, P.; Armstrong-James, D. Human NK cells develop an exhaustion phenotype during polar degranulation at the aspergillus fumigatus hyphal synapse. Front. Immunol. 2018, 9, 2344. [Google Scholar] [CrossRef]
- Tam, Y.K.; Maki, G.; Miyagawa, B.; Hennemann, B.; Tonn, T.; Klingemann, H.-G. Characterization of Genetically Altered, Interleukin 2-Independent Natural Killer Cell Lines Suitable for Adoptive Cellular Immunotherapy. Hum. Gene Ther. 1999, 10, 1359–1373. [Google Scholar] [CrossRef] [PubMed]
- Sahm, C.; Schönfeld, K.; Wels, W.S. Expression of IL-15 in NK cells results in rapid enrichment and selective cytotoxicity of gene-modified effectors that carry a tumor-specific antigen receptor. Cancer Immunol. Immunother. 2012, 61, 1451–1461. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, J.; Niu, J.; Zhang, J.; Tian, Z. Interleukin-15 improves cytotoxicity of natural killer cells via up-regulating NKG2D and cytotoxic effector molecule expression as well as STAT1 and ERK1/2 phosphorylation. Cytokine 2008, 42, 128–136. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmidt, S.; Luckowitsch, M.; Hogardt, M.; Lehrnbecher, T. Natural Killer Cell Line NK-92-Mediated Damage of Medically Important Fungi. J. Fungi 2021, 7, 144. https://doi.org/10.3390/jof7020144
Schmidt S, Luckowitsch M, Hogardt M, Lehrnbecher T. Natural Killer Cell Line NK-92-Mediated Damage of Medically Important Fungi. Journal of Fungi. 2021; 7(2):144. https://doi.org/10.3390/jof7020144
Chicago/Turabian StyleSchmidt, Stanislaw, Marie Luckowitsch, Michael Hogardt, and Thomas Lehrnbecher. 2021. "Natural Killer Cell Line NK-92-Mediated Damage of Medically Important Fungi" Journal of Fungi 7, no. 2: 144. https://doi.org/10.3390/jof7020144
APA StyleSchmidt, S., Luckowitsch, M., Hogardt, M., & Lehrnbecher, T. (2021). Natural Killer Cell Line NK-92-Mediated Damage of Medically Important Fungi. Journal of Fungi, 7(2), 144. https://doi.org/10.3390/jof7020144