
Bot Gummosis of Lemon (Citrus × limon) Caused by Neofusicoccum parvum

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
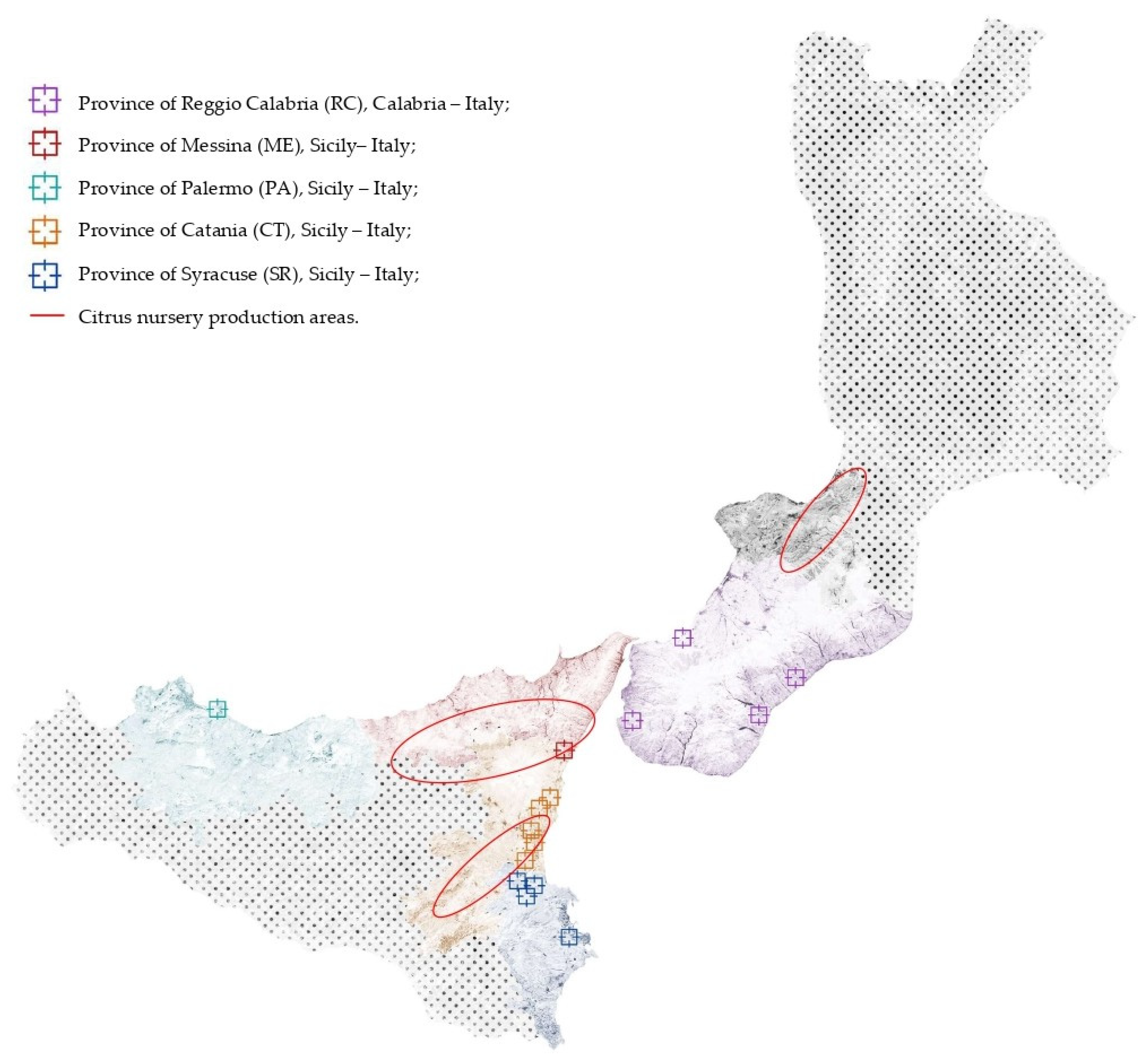
2.1. Fungal Collection and Isolation
2.2. Morphological Characteristics and Cardinal Temperatures for Growth of the Isolates
2.3. Amplification and Sequencing
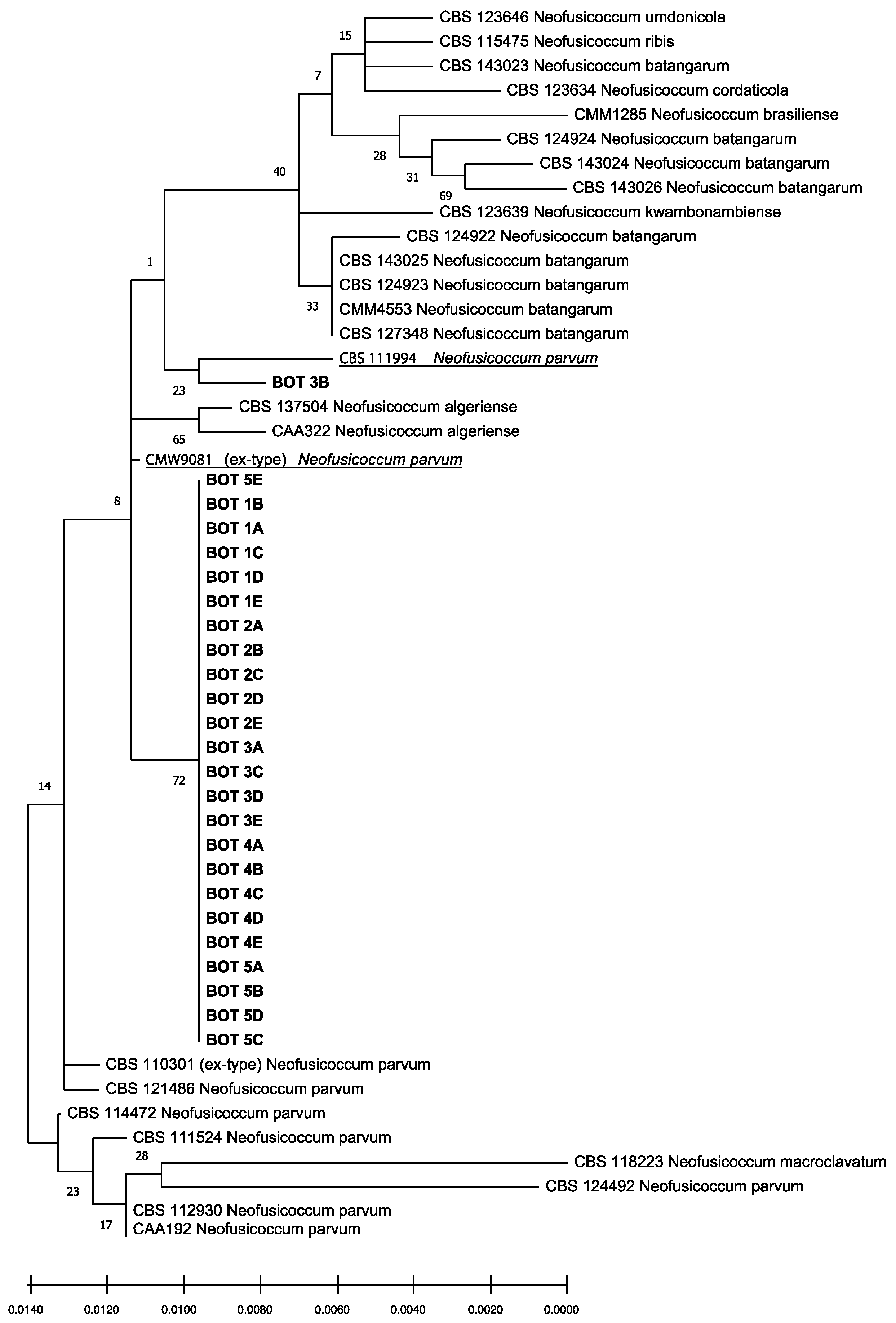
2.4. Molecular Identification and Phylogenetic Analyses
2.5. Pathogenicity Tests
3. Results
3.1. Symptoms
3.2. Fungus Isolation and Morphological Identification
3.3. Molecular Identification
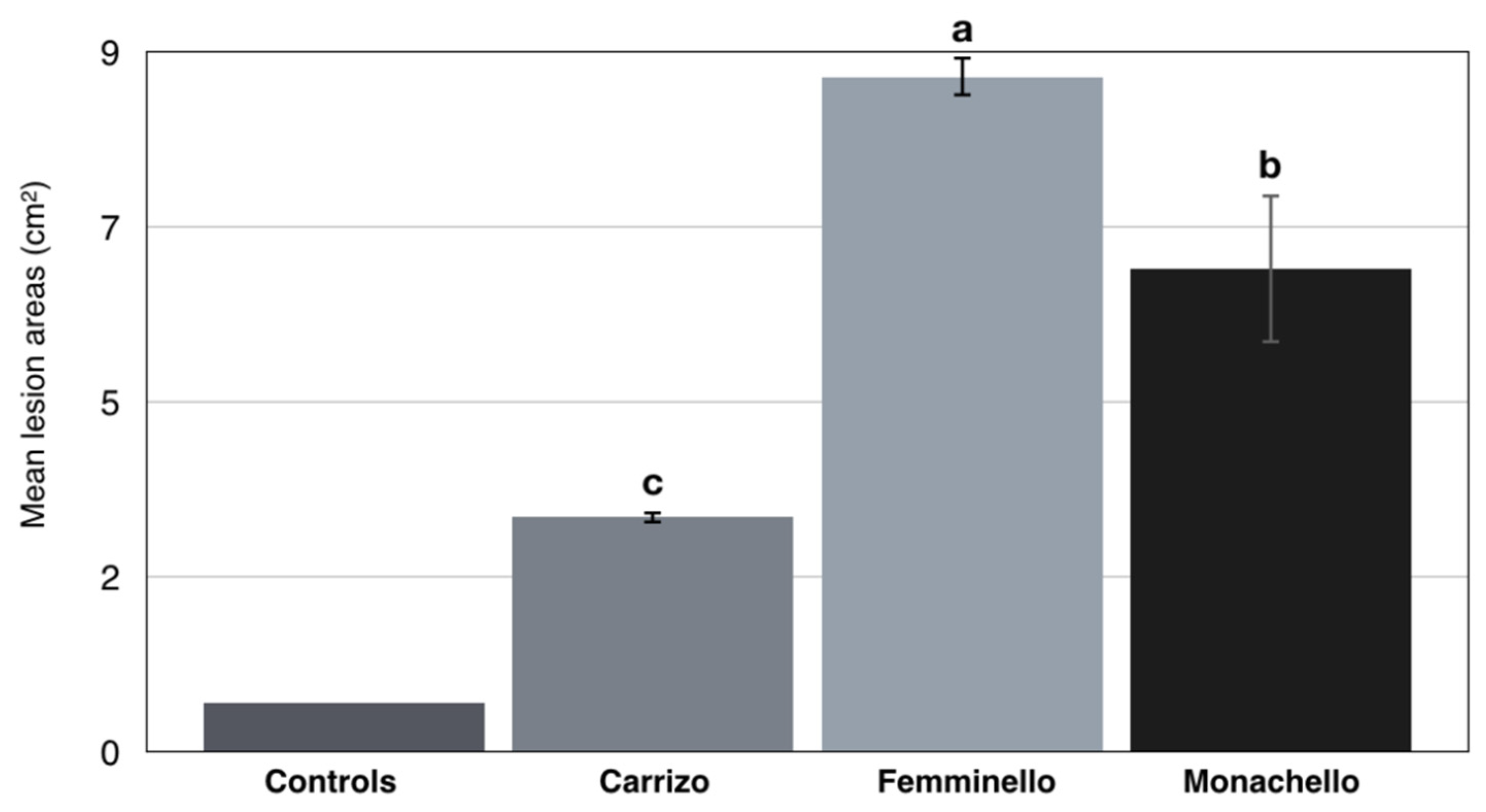
3.4. Pathogenicity Tests
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Klotz, L.J. Fungal, Bacterial, and Nonparasitic Diseases and Injuries Originating in the Seedbed, Nursery, and Orchard. In The Citrus Industry; Reuther, W., Calavan, E.C., Carman, G.E., Eds.; University of California-Division of Agricultural Sciences: Riverside, CA, USA, 1978; Volume 4, pp. 1–66. [Google Scholar]
- Scaramuzzi, G. Le Malattie degli Agumi; Edizioni Agricole: Bologna, Italy, 1965; pp. 12–17. [Google Scholar]
- Adesemoye, A.O.; Mayorquin, J.S.; Wang, D.H.; Twizeyimana, M.; Lynch, S.C.; Eskalen, A. Identification of Species of Botryosphaeriaceae Causing Bot gummosis in Citrus in California. Plant Dis. 2014, 98, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Phillips, A.J.L.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and Species Known from Culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, A.J.; Camporesi, E.; Hyde, K.D.; Phillips, A.J.L.; Fu, C.Y.; Yan, J.Y.; Li, X. Dothiorella Species Associated with Woody Hosts in Italy. Mycosphere 2016, 7, 51–63. [Google Scholar] [CrossRef]
- Phillips, A.J.L.; Hyde, K.D.; Alves, A.; Liu, J.K. Families in Botryosphaeriales: A Phylogenetic, Morphological and Evolutionary Perspective. Fungal Divers. 2019, 94, 1–22. [Google Scholar] [CrossRef]
- Zhang, W.; Groenewald, J.Z.; Lombard, L.; Schumacher, R.K.; Phillips, A.J.L.; Crous, P.W. Evaluating Species in Botryosphaeriales. Pers. Mol. Phylogeny Evol. Fungi 2021, 46, 63–115. [Google Scholar]
- Adesemoye, A.O.; Eskalen, A. First Report of Spencermartinsia viticola, Neofusicoccum australe, and N. parvum Causing Branch Canker of Citrus in California. Plant Dis. 2011, 95, 770. [Google Scholar] [CrossRef] [PubMed]
- Mayorquin, J.S.; Wang, D.H.; Twizeyimana, M.; Eskalen, A. Identification, Distribution, and Pathogenicity of Diatrypaceae and Botryosphaeriaceae Associated with Citrus Branch Canker in the Southern California Desert. Plant Dis. 2016, 100, 2402–2413. [Google Scholar] [CrossRef] [Green Version]
- Vakalounakis, D.J.; Ntougias, S.; Kavroulakis, N.; Protopapadakis, E. Neofusicoccum parvum and Diaporthe foeniculina Associated with Twig and Shoot Blight and Branch Canker of Citrus in Greece. J. Phytopathol. 2019, 167, 527–537. [Google Scholar] [CrossRef]
- Hamrouni, N.; Nouri, M.T.; Trouillas, F.P.; Said, A.; Sadfi-Zouaoui, N.; Hajlaoui, M.R. Dothiorella gummosis Caused by Dothiorella viticola, First Record from Citrus in Tunisia. New Dis. Rep. 2018, 38, 10. [Google Scholar] [CrossRef] [Green Version]
- Schena, L.; Burruano, S.; Giambra, S.; Surico, G.; Pane, A.; Evoli, M.; Magnano Di San Lio, G.; Cacciola, S.O. First Report of Neofusicoccum batangarum as Causal Agent of Scabby Cankers of Cactus Pear (Opuntia ficus-indica) in Minor Islands of Sicily. Plant Dis. 2018, 102, 445. [Google Scholar] [CrossRef]
- Aloi, F.; Giambra, S.; Schena, L.; Surico, G.; Pane, A.; Gusella, G.; Stracquadanio, C.; Burruano, S.; Cacciola, S.O. New Insights into Scabby Canker of Opuntia ficus-indica, Caused by Neofusicoccum batangarum. Phytopathol. Mediterr. 2020, 59, 269–284. [Google Scholar]
- Guarnaccia, V.; Crous, P.W. Emerging Citrus Diseases in Europe Caused by Species of Diaporthe. IMA Fungus 2017, 8, 317–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, A.J.L. Botryosphaeria Species Associated with Diseases of Grapevines in Portugal. Phytopathol. Mediterr. 2002, 41, 3–18. [Google Scholar]
- Slippers, B.; Fourie, G.; Crous, P.W.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Multiple Gene Sequences Delimit Botryosphaeria australis sp. nov. from B. lutea. Mycologia 2004, 96, 1030–1041. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Slippers, B.; Wingfield, M.J.; Rheeder, J.; Marasas, W.F.O.; Phillips, A.J.L.; Alves, A.; Burgess, T.; Barber, P.; Groenewald, J.Z. Phylogenetic Lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–253. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.F.; Harmon, P.F. Identification of Species in the Botryosphaeriaceae Family Causing Stem Blight on Southern Highbush Blueberry in Florida. Plant Dis. 2010, 94, 966–971. [Google Scholar] [CrossRef] [Green Version]
- McDonald, V.; Eskalen, A. Botryosphaeriaceae Species Associated with Avocado Branch Cankers in California. Plant Dis. 2011, 95, 1465–1473. [Google Scholar] [CrossRef] [Green Version]
- Schena, L.; Cooke, D.E.L. Assessing the Potential of Regions of the Nuclear and Mitochondrial Genome to Develop a “Molecular Tool Box” for the Detection and Characterization of Phytophthora Species. J. Microbiol. Methods 2006, 67, 70–85. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A Method for Designing Primer Sets for Speciation Studies in Filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of Primer Sets Designed for Use with the PCR to Amplify Conserved Genes from Filamentous Ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- FinchTV v.1.4.0. Available online: https://digitalworldbiology.com/FinchTV (accessed on 16 February 2021).
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Slippers, B.; Boissin, E.; Phillips, A.J.L.; Groenewald, J.Z.; Lombard, L.; Wingfield, M.J.; Postma, A.; Burgess, T.; Crous, P.W. Phylogenetic Lineages in the Botryosphaeriales: A Systematic and Evolutionary Framework. Stud. Mycol. 2013, 76, 31–49. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.; Phillips, A.J.L.; Alves, A. Mating Type Genes in the Genus Neofusicoccum: Mating Strategies and Usefulness in Species Delimitation. Fungal Biol. 2017, 121, 394–404. [Google Scholar] [CrossRef]
- Yang, T.; Groenewald, J.Z.; Cheewangkoon, R.; Jami, F.; Abdollahzadeh, J.; Lombard, L.; Crous, P.W. Families, Genera, and Species of Botryosphaeriales. Fungal Biol. 2017, 121, 322–346. [Google Scholar] [CrossRef]
- Lopes, A.; Barradas, C.; Phillips, A.J.L.; Alves, A. Diversity and Phylogeny of Neofusicoccum Species Occurring in Forest and Urban Environments in Portugal. Mycosphere 2016, 7, 906–920. [Google Scholar] [CrossRef]
- Vaidya, N.H.; Hadjicostis, C.N.; Dominguez-Garcia, A.D. Distributed Algorithms for Consensus and Coordination in the Presence of Packet-Dropping Communication Links–Part II: Coefficients of Ergodicity Analysis Approach. arXiv 2011, arXiv:1109.6392. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmer, L.W.; Garnsey, S.M.; Graham, J.H. Compendium of Citrus Diseases, 2nd ed.; American Phytopathological Society Press: Saint Paul, MN, USA, 2000; p. 92. [Google Scholar]
- Migheli, Q.; Cacciola, S.O.; Balmas, V.; Pane, A.; Ezra, D.; Magnano di San Lio, G. Mal Secco Disease Caused by Phoma tracheiphila: A Potential Threat to Lemon Production Worldwide. Plant Dis. 2009, 93, 852–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roccotelli, A.; Schena, L.; Sanzani, S.M.; Cacciola, S.O.; Mosca, S.; Faedda, R.; Ippolito, A.; Magnano di San Lio, G. Characterization of Basidiomycetes Associated with Wood Rot of Citrus in Southern Italy. Phytopathology 2014, 104, 851–858. [Google Scholar] [CrossRef] [Green Version]
- Golzar, H.; Burgess, T.I. Neofusicoccum parvum, a Causal Agent Associated with Cankers and Decline of Norfolk Island Pine in Australia. Australas. Plant Pathol. 2011, 40, 484. [Google Scholar] [CrossRef]
- Iturritxa, E.; Slippers, B.; Mesanza, N.; Wingfield, M.J. First Report of Neofusicoccum parvum Causing Canker and Die-back of Eucalyptus in Spain. Australas. Plant Dis. Notes 2011, 6, 57. [Google Scholar] [CrossRef] [Green Version]
- Spagnolo, A.; Marchi, G.; Peduto, F.; Phillips, A.J.L.; Surico, G. Detection of Botryosphaeriaceae Species within Grapevine Woody Tissues by Nested PCR, with Particular Emphasis on the Neofusicoccum parvum/N. ribis complex. Eur. J. Plant Pathol. 2011, 129, 485–500. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Gubler, W.D. Susceptibility of Grapevine Pruning Wounds to Infection by Lasiodiplodia theobromae and Neofusicoccum parvum. Plant Pathol. 2011, 60, 261–270. [Google Scholar] [CrossRef]
- Molina-Gayosso, E.; Silva-Rojas, H.V.; García-Morales, S.; Avila-Quezada, G. First Report of Black Spots on Avocado Fruit Caused by Neofusicoccum parvum in Mexico. Plant Dis. 2012, 96, 287. [Google Scholar] [CrossRef] [PubMed]
- Sakalidis, M.L.; Slippers, B.; Wingfield, B.D.; Hardy, G.E.S.J.; Burgess, T.I. The Challenge of Understanding the Origin, Pathways and Extent of Fungal Invasions: Global Populations of the Neofusicoccum parvum-N. ribis Species Complex. Divers. Distrib. 2013, 19, 873–883. [Google Scholar] [CrossRef]
- Moral, J.; Morgan, D.; Trapero, A.; Michailides, T.J. Ecology and Epidemiology of Diseases of Nut Crops and Olives Caused by Botryosphaeriaceae Fungi in California and Spain. Plant Dis. 2019, 103, 1809–1827. [Google Scholar] [CrossRef] [Green Version]
- Elvira-Recuenco, M.; Cacciola, S.O.; Sanz-Ros, A.V.; Garbelotto, M.; Aguayo, J.; Solla, A.; Mullet, M.; Drenkhan, T.; Oskay, F.; Aday Kaya, A.G.; et al. Potential Interactions between Invasive Fusarium circinatum and Other Pine Pathogens in Europe. Forests 2020, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Haenzi, M.; Cochard, B.; Chablais, R.; Crovadore, J.; Lefort, F. Neofusicoccum parvum, a New Agent of Sequoia Canker and Dieback Identified in Geneva, Switzerland. Forests 2021, 12, 434. [Google Scholar] [CrossRef]
- Úrbez-Torres, J.R.; Gubler, W.D. Pathogenicity of Botryosphaeriaceae Species Isolated from Grapevine Cankers in California. Plant Dis. 2009, 93, 584–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondello, V.; Songy, A.; Battiston, E.; Pinto, C.; Coppin, C.; Trotel-Aziz, P.; Clément, C.; Mugnai, L.; Fontaine, F. Grapevine Trunk Diseases: A Review of Fifteen Years of Trials for Their Control with Chemicals and Biocontrol Agents. Plant Dis. 2018, 102, 1189–1217. [Google Scholar] [CrossRef] [Green Version]
- Cunnington, J.H.; Priest, M.J.; Powney, R.A.; Cother, N.J. Diversity of Botryosphaeria Species on Horticultural Plants in Victoria and New South Wales. Austral. Plant Pathol. 2007, 157–159. [Google Scholar] [CrossRef]
- Bezerra, J.D.P.; Crous, P.W.; Aiello, D.; Gullino, M.L.; Polizzi, G.; Guarnaccia, V. Genetic Diversity and Pathogenicity of Botryosphaeriaceae Species Associated with Symptomatic Citrus Plants in Europe. Plants 2021, 10, 492. [Google Scholar] [CrossRef]
- Salerno, M.; Cutuli, G. Guida Illustrate di Patologia degli Agrumi; Edagrigole–Edizioni Agricole della Calderini: Bologna, Italy, 1994; pp. 30–35. [Google Scholar]
- Abou-Mansour, E.; Débieux, J.L.; Ramírez-Suero, M.; Bénard-Gellon, M.; Magnin-Robert, M.; Spagnolo, A.; Chong, J.; Farine, S.; Bertsch, C.; L’Haridon, F.; et al. Phytotoxic Metabolites from Neofusicoccum parvum, a Pathogen of Botryosphaeria dieback of Grapevine. Phytochemistry 2015, 115, 2017–2215. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Aloi, F.; Nocera, P.; Cacciola, S.O.; Surico, G.; Evidente, A. Phytotoxic Metabolites Isolated from Neufusicoccum batangarum, the Causal Agent of the Scabby Canker of Cactus Pear (Opuntia ficus-indica L.). Toxins 2020, 12, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvatore, M.M.; Alves, A.; Andolfi, A. Secondary Metabolites Produced by Neofusicoccum Species Associated with Plants: A Review. Agriculture 2021, 11, 149. [Google Scholar] [CrossRef]
- Blanco-Ulate, B.; Rolshausen, P.; Cantu, D. Draft Genome Sequence of Neofusicoccum parvum Isolate UCR-NP2, a Fungal Vascular Pathogen Associated with Grapevine Cankers. Genome Announc. 2013, 1, e00339-13. [Google Scholar] [CrossRef] [Green Version]
- Massonnet, M.; Figueroa-Balderas, R.; Galarneau, E.R.; Miki, S.; Lawrence, D.P.; Sun, Q.; Wallis, C.M.; Baumgartner, K.; Cantu, D. Neofusicoccum parvum Colonization of the Grapevine Woody Stem Triggers Asynchronous Host Responses at the Site of Infection and in the Leaves. Front. Plant. Sci. 2017, 8, 1117. [Google Scholar] [CrossRef] [Green Version]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as Endophytes and Latent Pathogens of Woody Plants: Diversity, Ecology and Impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Pavlic-Zupanc, D.; Wingfield, M.J.; Boissin, E.; Slippers, B. The Distribution of Genetic Diversity in the Neofusicoccum parvum/N. ribis Complex Suggests Structure Correlated with Level of Disturbance. Fungal Ecol. 2015, 13, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Cacciola, S.O.; Gilardi, G.; Faedda, R.; Schena, L.; Pane, A.; Garibaldi, A.; Gullino, M.L. Characterization of Colletotrichum ocimi Population Associated with Black Spot of Sweet Basil (Ocimum basilicum) in Northern Italy. Plants 2020, 9, 654. [Google Scholar] [CrossRef]
- Haridas, S.; Albert, R.; Binder, M.; Bloem, J.; LaButti, K.; Salamov, A.; Andreopoulos, B.; Baker, S.E.; Barry, K.; Bills, G. 101 Dothideomycetes Genomes: A Test Case for Predicting Lifestyles and Emergence of Pathogens. Stud. Mycol. 2020, 96, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Riolo, M.; Aloi, F.; Pane, A.; Cara, M.; Cacciola, S.O. Twig and Shoot Dieback of Citrus, a New Disease Caused by Colletotrichum Species. Cells 2021, 10, 449. [Google Scholar] [CrossRef] [PubMed]
- Pancher, M.; Ceol, M.; Corneo, P.E.; Longa, C.M.O.; Yousaf, S.; Pertot, I.; Campisano, A. Fungal Endophytic Communities in Grapevines (Vitis vinifera L.) Respond to Crop Management. Appl. Environ. Microbiol. 2012, 78, 4308–4317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, C.A.; Wingfield, M.J.; Slippers, B.; Altier, N.A.; Blanchette, R.A. Endophytic and Canker-associated Botryosphaeriaceae Occurring on Non-native Eucalyptus and Native Myrtaceae Trees in Uruguay. Fungal Divers. 2010, 41, 53. [Google Scholar] [CrossRef] [Green Version]
- Marsberg, A.; Kemler, M.; Jami, F.; Nagel, J.H.; Postma-Smidt, A.; Naidoo, S.; Wingfield, M.J.; Crous, P.W.; Spatafora, J.W.; Hesse, C.N.; et al. Botryosphaeria dothidea: A Latent Pathogen of Global Importance to Woody Plant Health. Mol. Plant. Pathol. 2017, 18, 477–488. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Species | Origin | Host, Variety | Accession Numbers | ||
|---|---|---|---|---|---|---|
| ITS | β-Tubulin | EF-1α | ||||
| BOT 1A | Neofusicoccum parvum | Catania, Siciliy-Italy | Citrus ×limon | MW727244 | MW789889 | MW789904 |
| BOT 1B | N. parvum | Catania, Siciliy-Italy | C. ×limon | MW727245 | MW789890 | MW789905 |
| BOT 1C | N. parvum | Catania, Siciliy-Italy | C. ×limon | MW727246 | MW789891 | MW789906 |
| BOT 1D | N. parvum | Catania, Siciliy-Italy | C. ×limon | MW727247 | MW789892 | MW789907 |
| BOT 1E | N. parvum | Catania, Siciliy-Italy | C. ×limon | MW727248 | MW789893 | MW789908 |
| BOT 2A | N. parvum | Syracuse, Siciliy-Italy | C. ×limon | MW727249 | MW789894 | MW789909 |
| BOT 2B | N. parvum | Syracuse, Siciliy-Italy | C. ×limon | MW727250 | MW789895 | MW789910 |
| BOT 2C | N. parvum | Syracuse, Siciliy-Italy | C. ×limon | MW727251 | MW789896 | MW789911 |
| BOT 2D | N. parvum | Syracuse, Siciliy-Italy | C. ×limon | MW727252 | MW789897 | MW789912 |
| BOT 2E | N. parvum | Syracuse, Siciliy-Italy | C. ×limon | MW727253 | MW789898 | MW789913 |
| BOT 4A | N. parvum | Messina, Siciliy-Italy | C. ×limon | MW788562 | MW789929 | MW789919 |
| BOT 4B | N. parvum | Messina, Siciliy-Italy | C. ×limon | MW788563 | MW789930 | MW789920 |
| BOT 4C | N. parvum | Messina, Siciliy-Italy | C. ×limon | MW788564 | MW789931 | MW789921 |
| BOT 4D | N. parvum | Messina, Siciliy-Italy | C. ×limon | MW788565 | MW789932 | MW789922 |
| BOT 4E | N. parvum | Messina, Siciliy-Italy | C. ×limon | MW788566 | MW789933 | MW789923 |
| BOT 5A | N. parvum | Palermo, Siciliy-Italy | C. ×limon | MW788567 | MW789934 | MW789924 |
| BOT 5B | N. parvum | Palermo, Siciliy-Italy | C. ×limon | MW788568 | MW789935 | MW789925 |
| BOT 5C | N. parvum | Palermo, Siciliy-Italy | C. ×limon | MW788569 | MW789936 | MW789926 |
| BOT 5D | N. parvum | Palermo, Siciliy-Italy | C. ×limon | MW788570 | MW789937 | MW789927 |
| BOT 5E | N. parvum | Palermo, Siciliy-Italy | C. ×limon | MW788571 | MW789938 | MW789928 |
| BOT 3A | N. parvum | Reggio Calabria, Calabria-Italy | C. ×limon | MW727254 | MW789899 | MW789914 |
| BOT 3B | N. parvum | Reggio Calabria, Calabria-Italy | C. ×limon | MW727255 | MW789900 | MW789915 |
| BOT 3C | N. parvum | Reggio Calabria, Calabria-Italy | C. ×limon | MW727256 | MW789901 | MW789916 |
| BOT 3D | N. parvum | Reggio Calabria, Calabria-Italy | C. ×limon | MW727257 | MW789902 | MW789917 |
| BOT 3E | N. parvum | Reggio Calabria, Calabria-Italy | C. ×limon | MW727258 | MW789903 | MW789918 |
| Species | Isolate | Origin | Host | Source | GenBank Accession Number | ||
|---|---|---|---|---|---|---|---|
| ITS | tef1 | β-Tubulin | |||||
| N. algeriense | CAA 322 | Portugal | Malus domestica | [29] | KX505906 | KX505894 | KX505916 |
| N. algeriense | CBS 137504 | Algeria | Vitis vinifera | [29] | KJ657702 | KX505893 | KX505915 |
| N. batangarum | CBS 124922 | Cameroon | Terminalia catappa | [29] | FJ900606 | FJ900652 | FJ900633 |
| N. batangarum | CBS 143023 | Italy, Favignana | Opuntia ficus-indica | [13] | MF414730 | MF414768 | MF414749 |
| N. batangarum | CBS 143025 | Italy, Linosa | Opuntia ficus-indica | [13] | MF414747 | MF414785 | MF414766 |
| N. batangarum | CBS 127348 | USA, Florida | Schinus terebinthifolius | [28] | HM357636 | KX464674 | KX464952 |
| N. batangarum | CMM4553 | Brasil | Anacardium sp. | Unpublished | KT728917 | KT728921 | KT728913 |
| N. batangarum | CBS 124923 | Cameroon | Terminalia catappa | [29] | FJ900608 | FJ900654 | FJ900635 |
| N. batangarum | CBS 124924 | Cameroon | Terminalia catappa | [29] | FJ900607 | FJ900653 | FJ900634 |
| N. batangarum | CBS 143026 | Italy, Lampedusa | Opuntia ficus-indica | [13] | MF414748 | MF414786 | MF414767 |
| N. batangarum | CBS 143024 | Italy, Ustica | Opuntia ficus-indica | [13] | MF414738 | MF414776 | MF414757 |
| N. brasiliense | CMM1285 | Brazil | Mangifera indica | [29] | JX513628 | JX513608 | KC794030 |
| N. cordaticola | CBS 123634 | South Africa | Syzygium cordatum | [29] | EU821898 | EU821868 | EU821838 |
| N. kwambonambiense | CBS 123639 | South Africa | Syzygium cordatum | [29] | EU821900 | EU821870 | EU821840 |
| N. macroclavatum | CBS 118223 | Australia | Eucalyptus globulus | [29] | DQ093196 | DQ093217 | DQ093206 |
| N. parvum | CBS 111994 | Australia | Telopea sp. | [28] | AF452519 | KX464702 | KX464982 |
| N. parvum | CAA 192 | Portugal | Ferula communis | [29] | KX505905 | KX505892 | KX505913 |
| N. parvum | CBS 112930 | South Africa | Vitis vinifera | [28] | AY343467 | AY343359 | KX464983 |
| N. parvum | CBS 121486 | Spain | Vitis vinifera cv. Parellada | [28] | EU650672 | KX464707 | KX464992 |
| N. parvum | CBS 114472 | USA | Leucadendron sp. | [28] | AF452523 | FJ150710 | KX464987 |
| N. parvum | CBS 111524 | USA, Hawaii | Protea cynaroides | [28] | AF452524 | FJ150709 | KX465009 |
| N. parvum | CBS 124492 | Zambia | Syzygium guineense | [28] | FJ655000 | KX464684 | KX464962 |
| N. parvum | CBS 110301(ex-type) | Portugal | Vitis vinifera | [29] | AY259098 | AY573221 | EU673095 |
| N. parvum | CMW9081 (ex-type) | New Zealand | Populus nigra | [29] | AY236943 | AY236888 | AY236917 |
| N. ribis | CBS 115475 | USA | Ribes sp. | [29] | AY236935 | AY236877 | AY236906 |
| N. umdonicola | CBS 123646 | South Africa | Syzygium cordatum | [29] | EU821905 | EU821875 | EU821845 |
| Isolates of N. parvum | 15 °C (mm d-1) ± S.D. a | 25 °C (mm d-1) ± S.D. a | 30 °C (mm d-1) ± S.D. a |
|---|---|---|---|
| BOT 1A | 3.73 ± 0.07 | 7.90 ± 1.08 | 6.71 ± 0.27 |
| BOT 1D | 3.11 ± 0.91 | 7.73 ± 0.28 | 7.38 ± 0.47 |
| BOT 2A | 3.41 ± 0.05 | 7.55 ± 0.32 | 7.06 ± 0.28 |
| BOT 2D | 3.53 ± 0.62 | 6.68 ± 0.90 | 5.58 ± 0.28 |
| BOT 3A | 3.45 ± 0.39 | 7.06 ± 0.28 | 6.23 ± 0.20 |
| BOT 3D | 3.30 ± 0.17 | 6.83 ± 0.32 | 6.63 ± 0.46 |
| BOT 4A | 3.42 ± 0.06 | 6.58 ± 0.62 | 7.38 ± 0.47 |
| BOT 5D | 3.28± 0.17 | 7.22 ± 0.42 | 5.37 ± 0.38 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aloi, F.; Riolo, M.; Parlascino, R.; Pane, A.; Cacciola, S.O. Bot Gummosis of Lemon (Citrus × limon) Caused by Neofusicoccum parvum. J. Fungi 2021, 7, 294. https://doi.org/10.3390/jof7040294
Aloi F, Riolo M, Parlascino R, Pane A, Cacciola SO. Bot Gummosis of Lemon (Citrus × limon) Caused by Neofusicoccum parvum. Journal of Fungi. 2021; 7(4):294. https://doi.org/10.3390/jof7040294
Chicago/Turabian StyleAloi, Francesco, Mario Riolo, Rossana Parlascino, Antonella Pane, and Santa Olga Cacciola. 2021. "Bot Gummosis of Lemon (Citrus × limon) Caused by Neofusicoccum parvum" Journal of Fungi 7, no. 4: 294. https://doi.org/10.3390/jof7040294
APA StyleAloi, F., Riolo, M., Parlascino, R., Pane, A., & Cacciola, S. O. (2021). Bot Gummosis of Lemon (Citrus × limon) Caused by Neofusicoccum parvum. Journal of Fungi, 7(4), 294. https://doi.org/10.3390/jof7040294