
First Documentation of Life Cycle Completion of the Alien Rust Pathogen Melampsoridium hiratsukanum in the Eastern Alps Proves Its Successful Establishment in This Mountain Range

 , , ,
, , ,  and
and
Abstract
:1. Introduction
- (1)
- confirm the identity of the rust fungus, putatively identified as the alien M. hiratsukanum, that occurs with its uredinial and telial stages on alder trees in the eastern Italian Alps; and
- (2)
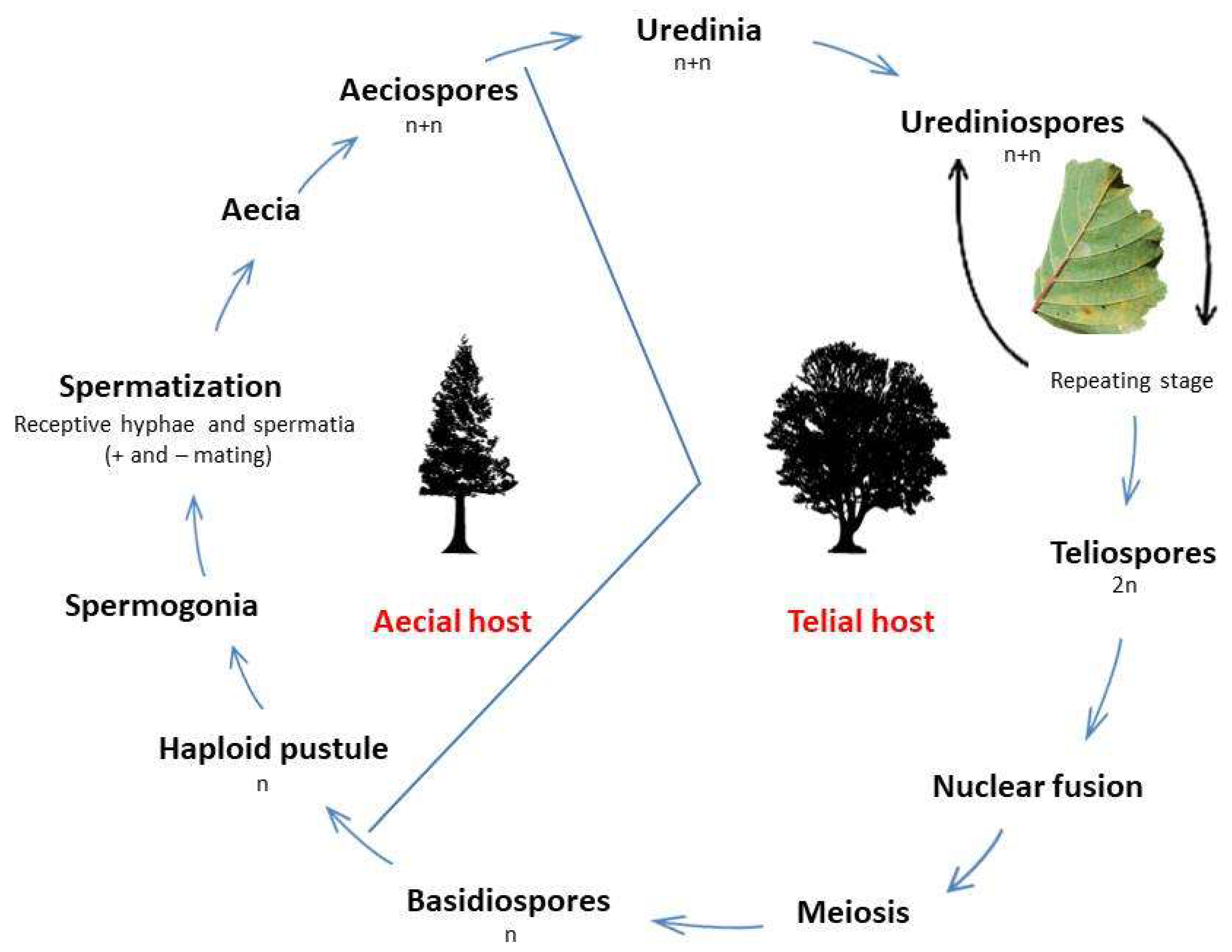
- ascertain if this M. hiratsukanum-like rust is the same rust that produces spermogonia and aecia on European larches (Larix decidua Mill.), growing in the Alps interspersed with, or adjacent to, infected alders.
2. Materials and Methods
2.1. Field Surveys
2.2. Plant Sample Collection and Storage
2.3. Morphological Identification
2.3.1. LM Observations
Uredinial Stage on Alder Leaves
Aecial Stage on Larch Needles
Uredinio-Spore Germination Tests
2.3.2. SEM Observations
2.3.3. Molecular Identification
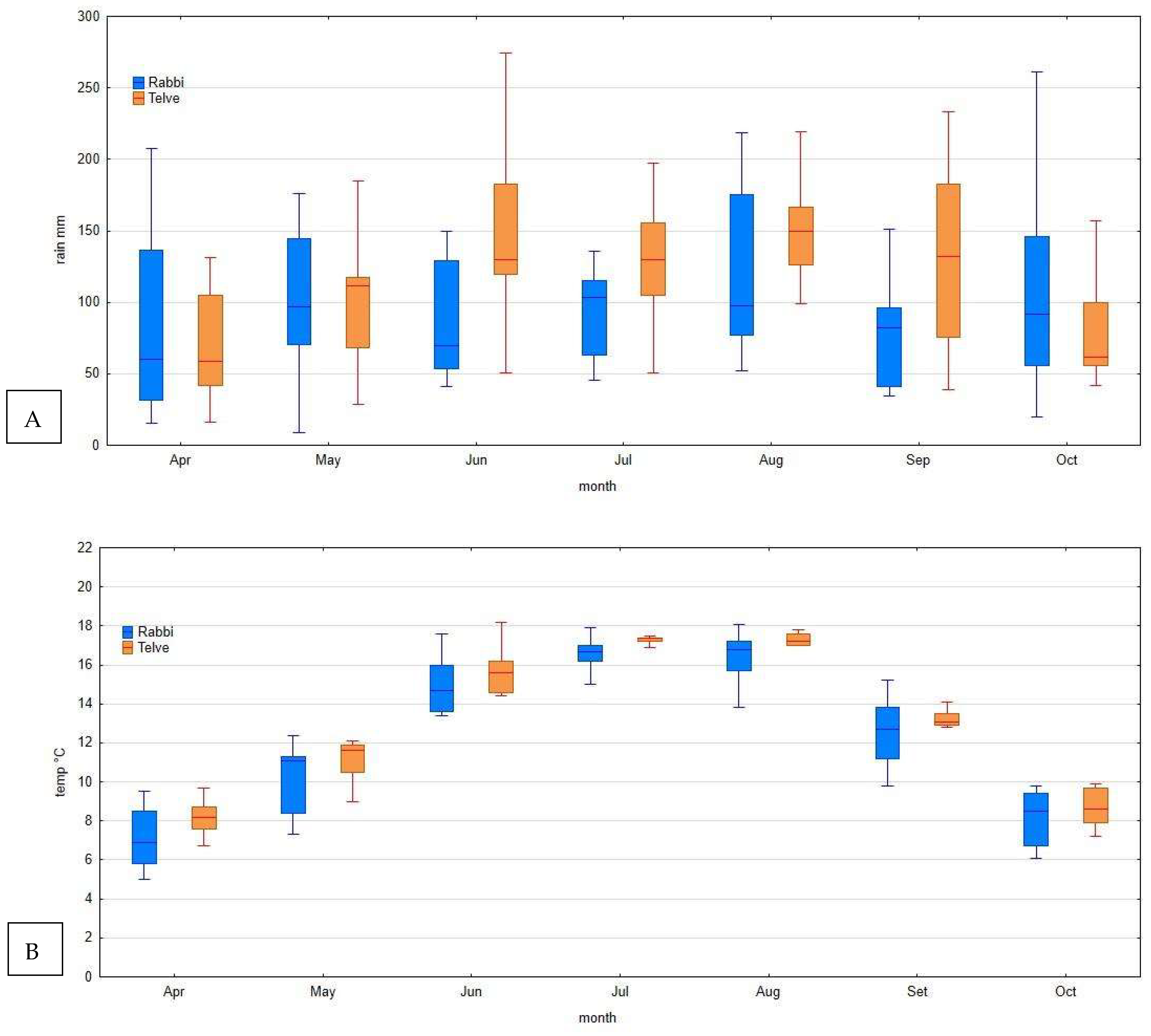
2.4. Meteorological Data
- -
- Telve (Pontarso) (Meteo Trentino meteorological network), 935 m a. s. l., placed at roughly 1 Km from the study sites of Val Campelle;
- -
- Rabbi (FEM agro-meteorological network), 1444 m a. s. l., located at 1 Km from the study site of Val di Rabbi.
3. Results
3.1. Field Surveys
3.1.1. Rust Occurrence on A. incana
3.1.2. Rust Occurrence on L. decidua
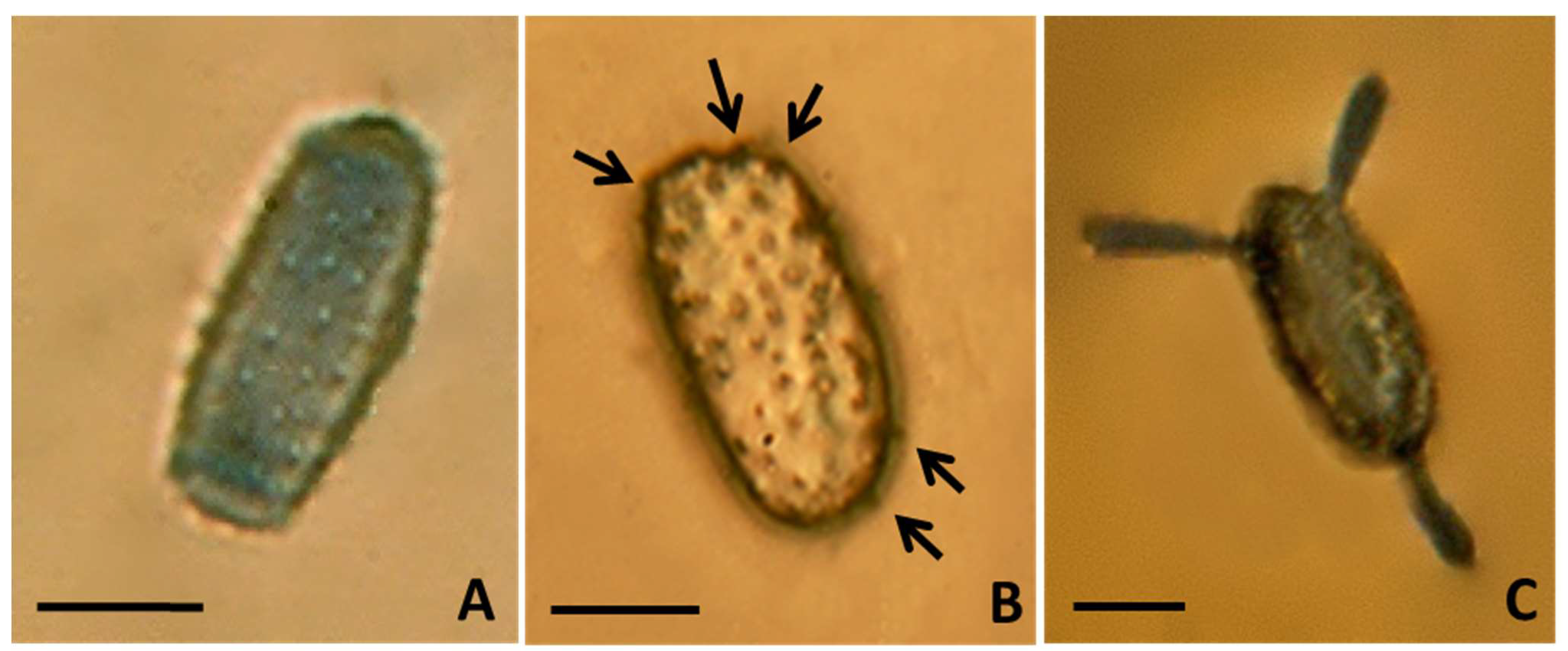
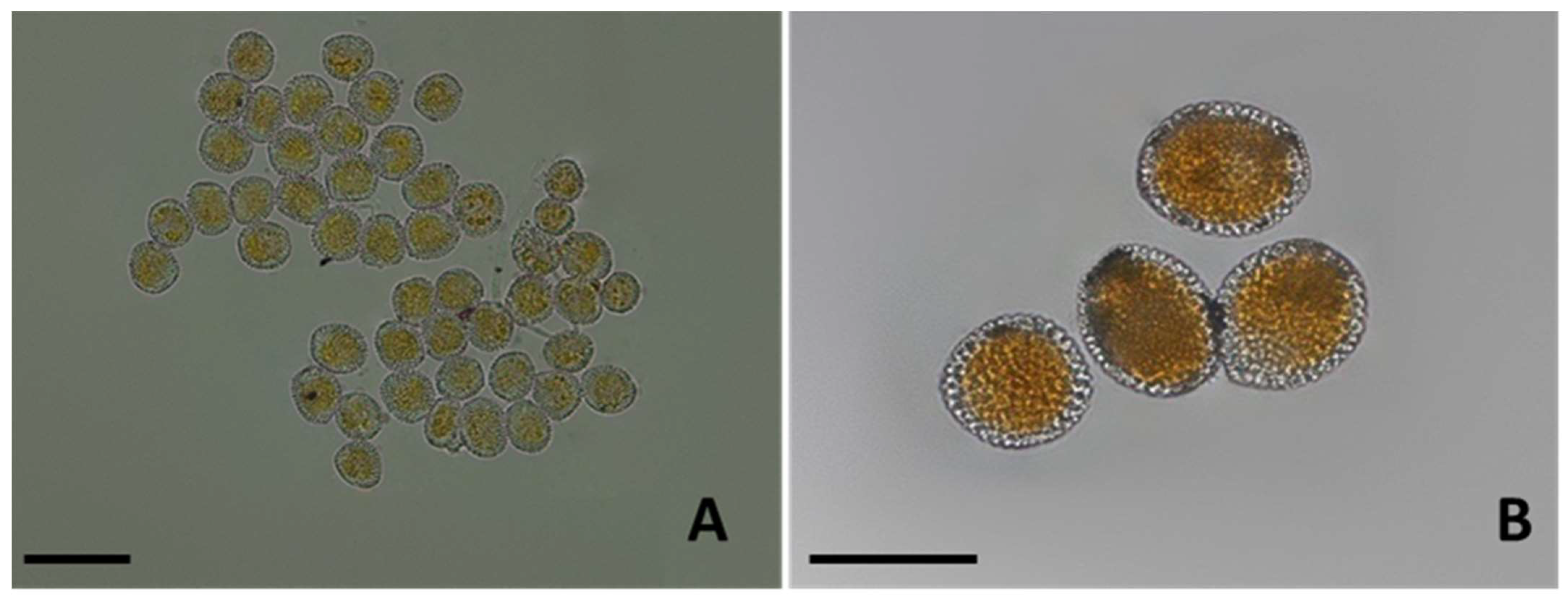
3.2. Morphological Identification
3.2.1. Micromorphological Analysis
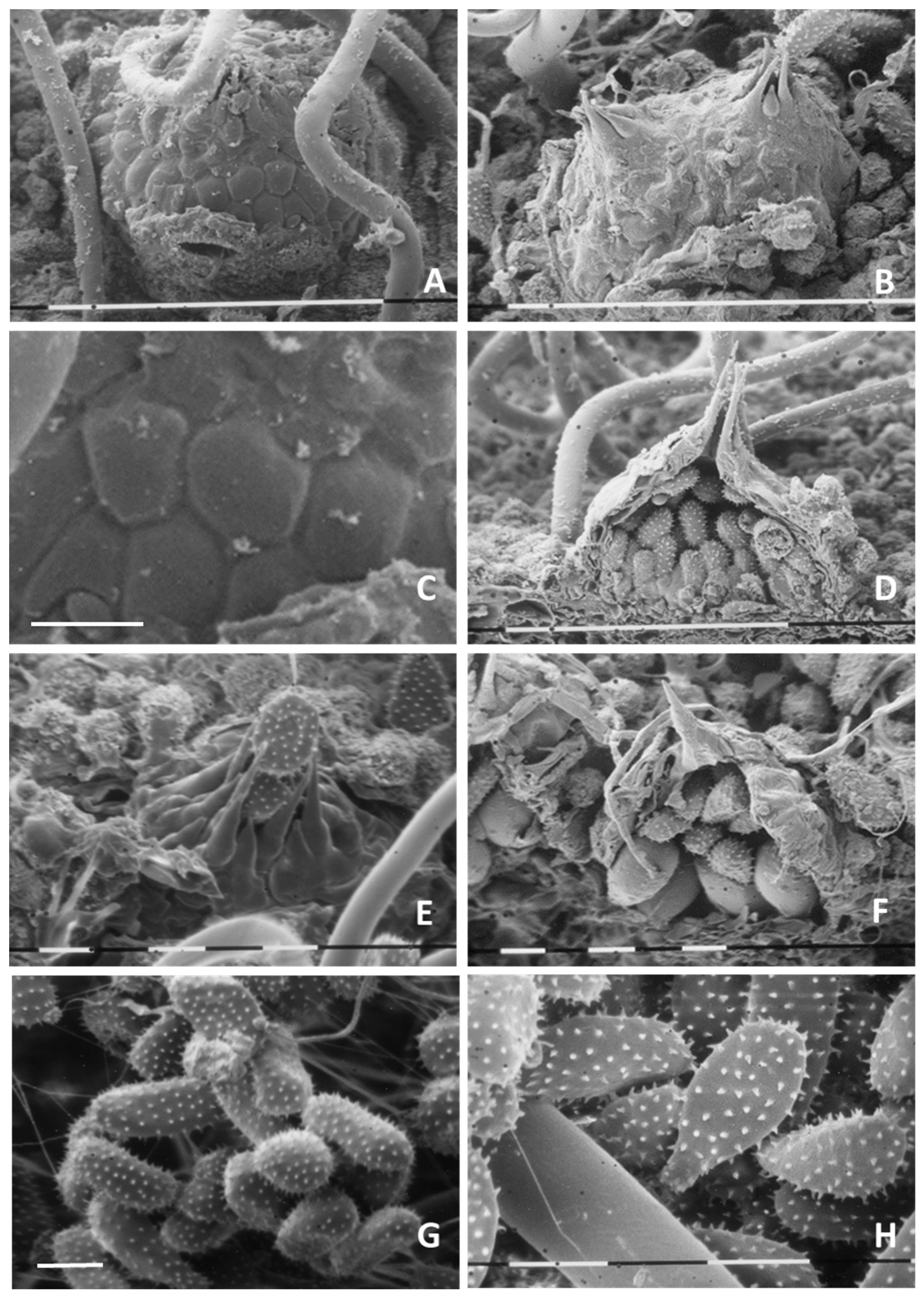
3.2.2. Ultrastructural Analysis
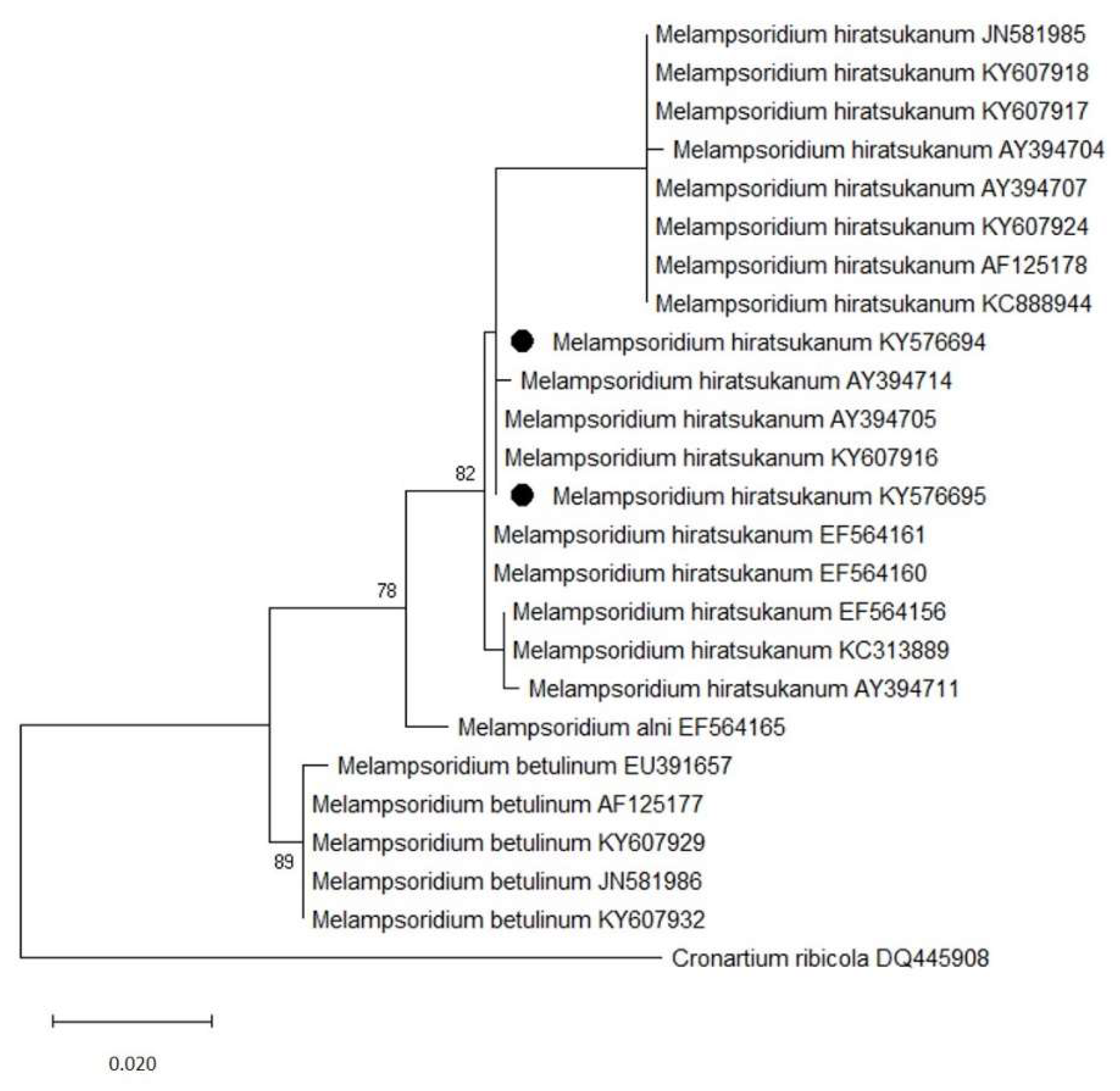
3.3. Molecular Identification
3.4. Meteorological Data
4. Discussion
- a marked propagule pressure (high production of rust inoculum on the telial host);
- host connectivity, with the broadleaf and the conifer hosts growing in contiguity;
- an environment conducive to disease, with high moisture during summer months, especially in July and August (meteorological data suggest the alpine environment to be highly favorable to rust survival, allowing the infection to intensify on the telial host and thus the complete colonization of this part of the grey alder range). This is different from what happens in northern European alder stands where the disease has sometimes a variable and erratic impact);
- larch infection is not required for rust survival: this rust can survive as mycelium in the scales of dormant alder buds [1] and thus persist in the environment regardless of the presence/absence of its aecial host (which becomes, however, crucial for the completion of the rust life cycle).
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hantula, J.; Scholler, M. NOBANIS—Invasive Alien Species Fact Sheet—Melampsoridium hiratsukanum. From: Online Database of the European Network on Invasive Alien Species—NOBANIS 2013. Available online: https://www.nobanis.org/fact-sheets/ (accessed on 23 March 2021).
- Brown, J.K.M.; Hovmøller, M.S. Aerial dispersal of pathogens on the global and continental scales and its impact on plant disease. Science 2002, 297, 537–541. [Google Scholar] [CrossRef] [Green Version]
- Mundt, C.C.; Sackett, K.E.; Wallace, L.D.; Cowger, C.; Dudley, J.P. Aerial dispersal and multiple-scale spread of epidemic disease. EcoHealth 2009, 6, 546–552. [Google Scholar] [CrossRef]
- Hantula, J.; Kurkela, T.; Hendry, S.; Yamaguchi, T. Morphological measurements and ITS sequences show that the new alder rust in Europe is conspecific with Melampsoridium hiratsukanum in eastern Asia. Mycologia 2009, 101, 622–631. [Google Scholar] [CrossRef]
- Wolczanska, A. Melampsoridium hiratsukanum (Uredinales), a new species for Poland. Acta Mycol. 1999, 34, 345–347. [Google Scholar] [CrossRef] [Green Version]
- Szabo, I. First report of Melampsoridium hiratsukanum on common alder in Hungary. Plant Pathol. 2002, 51, 804. [Google Scholar] [CrossRef]
- Riegler-Hager, H.; Scheuer, C.; Zwetko, P. Der Erlen-Rost Melampsoridium hiratsukanum in Österreich. Wulfenia 2003, 10, 135–143. [Google Scholar]
- Sert, H.; Sumbul, H. First report of Melampsoridium hiratsukanum infecting alder (Alnus orientalis var. orientalis) in Turkey. Plant Pathol. 2005, 54, 241. [Google Scholar] [CrossRef]
- Saura, S.; Bodin, Ö.; Fortin, M.J. Stepping stones are crucial for species’ long-distance dispersal and range expansion through habitat networks. J. Appl. Ecol. 2014, 51, 171–182. [Google Scholar] [CrossRef]
- Philibert, A.; Desprez-Loustau, M.-L.; Fabre, B.; Frey, P.; Halkett, F.; Husson, C.; Lung-Escarmant, B.; Marçais, B.; Robin, C.; Vacher, C.; et al. Predicting invasion success of forest pathogenic fungi from species traits. J. Appl. Ecol. 2011, 48, 1381–1390. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Hyakumachi, M. Feeding on plant pathogenic fungi by invertebrates: Comparison with saprotrophic and mycorrhizal systems. In Biotic Interactions in Plant-Pathogen Associations; Jeger, M.J., Spence, N.J., Eds.; CABI Publishing: Wallingford, UK, 2001; pp. 63–85. [Google Scholar]
- Moricca, S.; Maresi, G. Melampsoridium hiratsukanum reported for the first time on grey alder in Italy. New Dis. Rep. 2010, 21, 17. [Google Scholar] [CrossRef] [Green Version]
- Kuprevich, V.T.; Tranzschel, V. Flora Plantarum Cryptogamarum URSS; Fungi (1) Uredinales. Fasc. 1. Familia Melampsoraceae; Typis et Impensis Academiae Scientiarum URSS: Moscow, Russia, 1957; Volume 4. [Google Scholar]
- Gäumann, E. Die Rostpilze Mitteleuropas; Beitr Cryptogamenfl Schweiz 12; Buchdruckerei Büchler: Bern, Switzerland, 1959; p. 1407. [Google Scholar]
- Roll-Hansen, F.; Roll-Hansen, H. Melampsoridium on Alnus in Europe. M. alni conspecific with M. betulinum. Eur. J. For. Pathol. 1981, 11, 77–87. [Google Scholar]
- Wilson, M.; Henderson, D.M. British Rust Fungi; Cambridge University Press: Cambridge, UK, 1966. [Google Scholar]
- Henderson, D.M.; Bennel, A.P. British rust fungi: Additions and corrections. Notes R. Bot. Gard. Edinb. 1979, 37, 475–501. [Google Scholar]
- Hantula, J.; Nigel Stringer, R.; Lilja, A.; Kurkela, T. Alder rust, Melampsoridium hiratsukanum Ito, identified from Wales, UK and British Columbia, Canada. For. Pathol. 2012, 42, 348–350. [Google Scholar] [CrossRef]
- Kaneko, S.; Hiratsuka, N. Classification of the Melampsoridium species based on the position of urediniospore germ pores. Trans. Mycol. Soc. Jpn. 1981, 22, 463–473. [Google Scholar]
- Kurkela, T.; Hanso, M.; Hantula, J. Differentiating characteristics between Melampsoridium rusts infecting birch and alder leaves. Mycologia 1999, 91, 987–992. [Google Scholar] [CrossRef]
- Pisetta, M.; Montecchio, L.; Longa, C.M.O.; Salvadori, C.; Zottele, F.; Maresi, G. Green alder decline in the Italian Alps. For. Ecol. Manag. 2012, 281, 75–83. [Google Scholar] [CrossRef]
- Moricca, S.; Ragazzi, A. Axenic culture of the aecial state of Cronartium flaccidum from Italy. Mycol. Res. 1994, 98, 1258–1262. [Google Scholar] [CrossRef]
- Moricca, S.; Ragazzi, A.; Mitchelson, K.R. Molecular and conventional detection and identification of Cladosporium tenuissimum on two-needle pine rust aeciospores. Can. J. Bot. 1999, 77, 339–347. [Google Scholar]
- Assante, G.; Maffi, D.; Saracchi, M.; Farina, G.; Moricca, S.; Ragazzi, A. Hystological studies on the mycoparasitism of Cladosporium tenuissimum on urediniospores of Uromyces appendiculatus. Mycol. Res. 2004, 108, 170–183. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; the UGENE Team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [Green Version]
- Moricca, S.; Ginetti, B. Saprophytic Growth of the Alder Rust Fungus Melampsoridium hiratsukanum on Artificial Media. Fungal Biol. UK 2015, 119, 568–579. [Google Scholar] [CrossRef] [Green Version]
- Howie, J.A. Adult Plant Resistance to Fungal Pathogens of Wheat. Ph.D. Thesis, Open University, Milton Keynes, UK, 1997. [Google Scholar]
- Maguire, D.A.; Kanaskie, A.; Voelker, W.; Johnson, R.; Johnson, G. Growth of young Douglas-fir plantations across a gradient in Swiss needle cast severity. West. J. Appl. For. 2002, 17, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Prosser, F.; Bertolli, A. Atlante di 50 Specie Esotiche del Trentino; LIFE + T.E.N Azione C18; Provincia Autonoma di Trento: Trento, Italy, 2015; 112p. [Google Scholar]
- Bregant, C.; Sanna, G.P.; Bottos, A.; Maddau, L.; Montecchio, L.; Linaldeddu, B.T. Diversity and Pathogenicity of Phytophthora Species Associated with Declining Alder Trees in Italy and Description of Phytophthora alpina sp. nov. Forests 2020, 11, 848. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diagnostic Features | Melampsoridium alni | Melampsoridium betulinum | Melampsoridium hiratsukanum |
|---|---|---|---|
| Urediniospore morphology | From subfusoid to subclavate | From subfusoid to subclavate | Ovoid to oblong-obovoid |
| Urediniospore size (average, μm) | 33.5 × 11.5 | 30.9 × 13.4 | 26.0 × 13.3 |
| Distribution of echinulae | Absent at the two spore ends | Absent at the two spore ends | All over the spore surface |
| Length of uredinial ostiolar cells (range, μm) | 20–25 | 20–25 | 30–45 |
| Number and arrangement of germ pores | Two, one at each spore end | Up to six, on the two spore ends | Up to six, on the two spore ends |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moricca, S.; Benigno, A.; Oliveira Longa, C.M.; Cacciola, S.O.; Maresi, G. First Documentation of Life Cycle Completion of the Alien Rust Pathogen Melampsoridium hiratsukanum in the Eastern Alps Proves Its Successful Establishment in This Mountain Range. J. Fungi 2021, 7, 617. https://doi.org/10.3390/jof7080617
Moricca S, Benigno A, Oliveira Longa CM, Cacciola SO, Maresi G. First Documentation of Life Cycle Completion of the Alien Rust Pathogen Melampsoridium hiratsukanum in the Eastern Alps Proves Its Successful Establishment in This Mountain Range. Journal of Fungi. 2021; 7(8):617. https://doi.org/10.3390/jof7080617
Chicago/Turabian StyleMoricca, Salvatore, Alessandra Benigno, Claudia Maria Oliveira Longa, Santa Olga Cacciola, and Giorgio Maresi. 2021. "First Documentation of Life Cycle Completion of the Alien Rust Pathogen Melampsoridium hiratsukanum in the Eastern Alps Proves Its Successful Establishment in This Mountain Range" Journal of Fungi 7, no. 8: 617. https://doi.org/10.3390/jof7080617
APA StyleMoricca, S., Benigno, A., Oliveira Longa, C. M., Cacciola, S. O., & Maresi, G. (2021). First Documentation of Life Cycle Completion of the Alien Rust Pathogen Melampsoridium hiratsukanum in the Eastern Alps Proves Its Successful Establishment in This Mountain Range. Journal of Fungi, 7(8), 617. https://doi.org/10.3390/jof7080617