
Current Insight into Culture-Dependent and Culture-Independent Methods in Discovering Ascomycetous Taxa

 ,
,  and
and 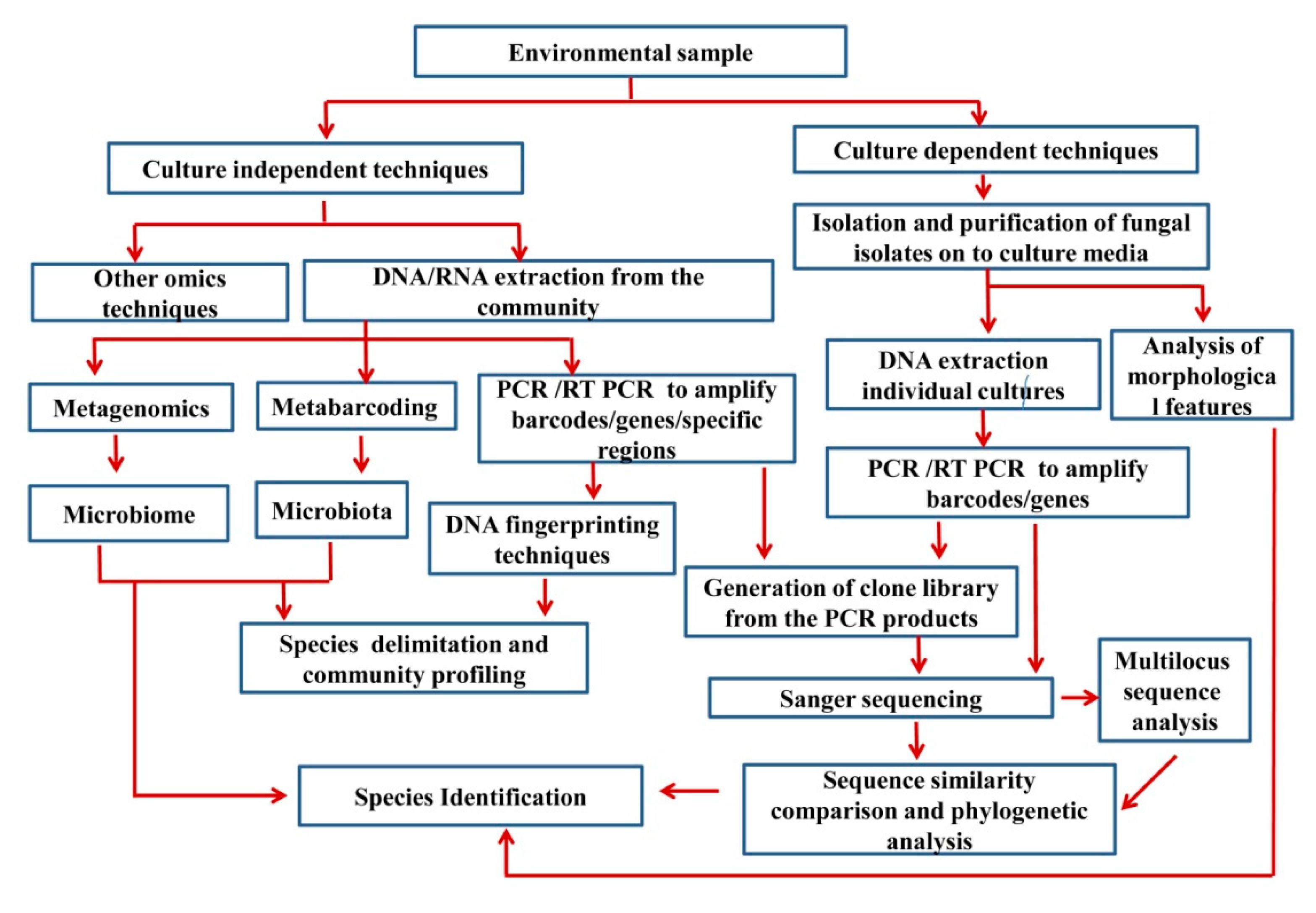{kind=link}
Abstract
:1. Introduction
2. Cultures in Traditional Taxonomy (i.e., Morphology-Based Taxonomy)
2.1. To Resolve Species Boundaries of Cryptic Species
2.2. To Reveal Pleomorphism and Propose a Single Name
2.3. To Isolate Mycobionts of Lichens
3. Cultures to Extract Secondary Metabolites
4. Cultures as a Source for DNA
4.1. Isolation
4.2. Culture Maintenance and Culture Collections
5. Limitations of Culture-Based Studies
6. Culture-Independent Methods
6.1. Dark Fungi and Culture-Independent Techniques to Detect Them
6.1.1. Endophytic Taxa; an Example of a Life Mode That Needs More Work
6.1.2. Marine Fungi as an Example
6.2. Studies on Older Herbarium Specimens
7. Towards Sequence-Based Nomenclature
8. Future Directions
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Species Fungorum. Available online: http://www.speciesfungorum.org/ (accessed on 20 July 2021).
- Hawksworth, D.L.; Lücking, R. Fungal diversity revisited: 2.2 to 3.8 million species. Microbiol. Spectr. 2017, 5, 79–95. [Google Scholar] [CrossRef]
- Nichols, D.; Cahoon, N.; Trakhtenberg, E.M.; Pham, L.; Mehta, A.; Belanger, A.; Kanigan, T.; Lewis, K.; Epstein, S.S. Use of ichip for highthroughput in situ cultivation of “uncultivable” microbial species. Appl. Environ. Microbiol. 2010, 76, 2445–2450. [Google Scholar] [CrossRef] [Green Version]
- Berdy, B.; Spoering, A.L.; Ling, L.L.; Epstein, S.S. In situ cultivation of previously uncultivable microorganisms using the ichip. Nat. Protoc. 2017, 12, 2232–2242. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Hussain, M.; Zhang, W.; Stadler, M.; Liu, X.; Xiang, M. Current insights into fungal species diversity and perspective on naming the environmental DNA sequences of fungi. Mycology 2019, 10, 127–140. [Google Scholar] [CrossRef] [Green Version]
- Dai, D.-Q.; Wijayawardene, N.N.; Tang, L.-Z.; Liu, C.; Han, L.-H.; Chu, H.-L.; Wang, H.-B.; Liao, C.-F.; Yang, E.-F.; Xu, R.-F.; et al. Rubroshiraia gen. nov., a second hypocrellin-producing genus in Shiraiaceae (Pleosporales). MycoKeys 2019, 58, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Guba, E.F. Monograph of Pestalotia and Monochaetia; Harvard University Press: Cambridge, MA, USA, 1961. [Google Scholar]
- Sutton, B.C. The Coelomycetes: Fungi Imperfecti with Pycnidia, Acervuli and Stromata; Commonwealth Mycological Institute: London, UK, 1980. [Google Scholar]
- Uecker, F.A.; Raj, T.R.N. Coelomycetous Anamorphs with Appendage-Bearing Conidia. Mycologia 1994, 86, 308. [Google Scholar] [CrossRef]
- Brown, W. Experiments on the growth of fungi on culture media. Ann. Bot. 1923, 37, 105–129. [Google Scholar] [CrossRef]
- Nobles, M.K. Studies in forest pathology: Identification of cultures of wood-rotting fungi. Can. J. Res. 1948, 26, 281–431. [Google Scholar] [CrossRef]
- Sutton, B.C. Colletotrichum dematium (Pers. ex Fr.) Grove and C. trichellum (Fr. ex Fr.) Duke. Trans. Br. Mycol. Soc. 1962, 45, 222–232. [Google Scholar] [CrossRef]
- Sutton, B.C. The appressoria of Colletotrichum graminicola and C. falcatum. Can. J. Bot. 1968, 46, 873–876. [Google Scholar] [CrossRef]
- Boerema, G.H.; Howeler, L.H. Phoma exigua Desm. and its varieties. Persoonia 1967, 5, 15–28. [Google Scholar]
- Boerema, G.H.; Dorenbosch, M.M.J.; van Kesteren, H.A. Remarks on Species of Phoma Referred to Peyronellaea, V. Kew Bull. 1977, 31, 533–544. [Google Scholar] [CrossRef]
- Boerema, G.H.; Dorenbosch, M.M.J.; van Kesteren, H.A. Remarks on species of Phoma referred to Peyronellaea. Persoonia 1965, 4, 47–68. [Google Scholar] [CrossRef]
- Boerema, G.H.; Dorenbosch, M.M.J.; van Kesteren, H.A. Remarks on species of Phoma referred to Peyronellaea—III. Persoonia 1971, 6, 171–177. [Google Scholar]
- Boerema, G.H.; Dorenbosch, M.M.J.; van Kesteren, H.A. Remarks on species of Phoma referred to Peyronellaea—IV. Persoonia 1973, 7, 131–139. [Google Scholar]
- Boerema, G.H.; Dorenbosch, M.M.J.; van Kesteren, H.A. Remarks on species of Phoma referred to Peyronellaea. Persoonia 1968, 5, 201–205. [Google Scholar] [CrossRef]
- Boerema, G.H.; Gruyter, J.; de Noordeloos, M.E.; Hamers, M.E.C. Phoma identifcation manual. In Differentiation of Specific and Infra-Specific Taxa in Culture; CABI Publishing: Wallingford, UK, 2004. [Google Scholar]
- Boerema, G.H.; Dorenbosch, M.M.J. The Phoma and Ascochyta species described by Wollenweber and Hochapfel in their study on fruit-rotting. Stud. Mycol. 1973, 3, 1–50. [Google Scholar]
- Boerema, G. The Phoma species studied in culture by Dr. R.W.G. Dennis. Trans. Br. Mycol. Soc. 1976, 67, 289–319. [Google Scholar] [CrossRef]
- Simmonds, J.H. A study of the species of Colletotrichum causing ripe fruit rots in Queensland. Qd. J. Agric. Anim. Sci. 1965, 22, 437–459. [Google Scholar]
- Sutton, B.; Alcorn, J. Additions to Hyphopolynema (Hyphomycetes). Aust. J. Bot. 1984, 32, 551–559. [Google Scholar] [CrossRef]
- Okada, G.; Tubaki, K. A new species and a new variety of Endocalyx (Deuteromycotina) from Japan. Mycologia 1984, 76, 300–313. [Google Scholar] [CrossRef]
- Holdenrieder, O.; Kowalski, T. Pycnidial formation and pathogenicity in Tubakia dryina. Mycol. Res. 1989, 92, 166–169. [Google Scholar] [CrossRef]
- Kendrick, W.B.; Nag Raj, T.R. Morphological terms in fungal imperfecti. In The Whole Fungus; Kendrick, W.B., Ed.; Nat. Mus.: Ottawa, ON, Canada, 1979; pp. 43–61. [Google Scholar]
- Walker, J.; Sutton, B.C. Dilophta Sacc. and Dilophospora Desm. Trans. Br. Mycol. Soc. 1974, 62, 231–241. [Google Scholar] [CrossRef]
- Aime, M.C.; Miller, A.N.; Aoki, T.; Bensch, K.; Cai, L.; Crous, P.W.; Hawksworth, D.L.; Hyde, K.D.; Kirk, P.M.; Lücking, R.; et al. How to publish a new fungal species, or name, version 3.0. IMA Fungus 2021, 12, 1–5. [Google Scholar] [CrossRef]
- Wanasinghe, D.; Mortimer, P.; Xu, J. Insight into the systematics of microfungi colonizing dead woody twigs of Dodonaea viscosa in Honghe (China). J. Fungi 2021, 7, 180. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, M.M.; Wang, X.G.; Cheng, X.L.; Guo, J.J.; Bian, X.M.; Cai, L. Fungal communities in sediments of subtropical Chinese seas as estimated by DNA metabarcoding. Sci. Rep. 2016, 6, 26528. [Google Scholar] [CrossRef]
- Hughes, S.J. New Zealand Fungi 31. Capnobotrys, an anamorph of Metacapnodiaceae. N. Z. J. Bot. 1981, 19, 193–226. [Google Scholar] [CrossRef] [Green Version]
- Barber, P.A.; Burgess, T.J.; Hardy, G.; Slippers, B.; Keane, P.J.; Wingfield, M.J. Botryosphaeria species from Eucalyptus in Australia are pleoanamorphic, producing Dichomera synanamorphs in culture. Mycol. Res. 2005, 109, 1347–1363. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Summerell, B.A.; Carnegie, A.; Mohammed, C.; Himaman, W.; Groenewald, J.Z. Foliicolous Mycosphaerella spp. and their anamorphs on Corymbia and Eucalyptus. Fungal Divers. 2007, 26, 143–185. [Google Scholar]
- Crous, P.; Summerell, B.; Carnegie, A.; Wingfield, M.; Groenewald, J. Novel species of Mycosphaerellaceae and Teratosphaeriaceae. Pers. Mol. Phylogeny Evol. Fungi 2009, 23, 119–146. [Google Scholar] [CrossRef] [Green Version]
- Udayanga, D.; Liu, X.; McKenzie, E.H.C.; Chukeatirote, E.; Bahkali, A.H.A.; Hyde, K.D. The genus Phomopsis: Biology, applications, species concepts and names of common phytopathogens. Fungal Divers. 2011, 50, 189–225. [Google Scholar] [CrossRef]
- Mihaescu, C.; Dunea, D.; Bășa, A.G.; Frasin, L.N. Characteristics of Phomopsis juglandina (Sacc.) Hohn. Associated with dieback of walnut in the climatic conditions of Southern Romania. Agronomy 2020, 11, 46. [Google Scholar] [CrossRef]
- Hawksworth, D.L. Managing and coping with names of pleomorphic fungi in a period of transition. IMA Fungus 2012, 3, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Wijayawardene, N.N.; Crous, P.W.; Kirk, P.M.; Hawksworth, D.L.; Boonmee, S.; Braun, U.; Dai, D.-Q.; D’Souza, M.J.; Diederich, P.; Dissanayake, A.J.; et al. Naming and outline of Dothideomycetes–2014 including proposals for the protection or suppression of generic names. Fungal Divers. 2014, 69, 1–55. [Google Scholar] [CrossRef] [PubMed]
- Samuels, G.J.; Rogers, J.D. Botryohypoxylon gen. nov. and its anamorph, Iledon gen. nov. Mycotaxon 1986, 25, 629–637. [Google Scholar]
- Ahmadjian, V. Methods of isolating and culturing lichen symbionts and thalli. In The Lichens; Ahmadjian, V., Hale, M.E., Eds.; Academic Press: New York, NY, USA, 1973; pp. 653–659. [Google Scholar]
- Yamamoto, Y.; Kinoshita, Y.; Takahagi, T.; Kroken, S.; Kurokawa, T.; Yoshimura, I. Factors affecting discharge and germination of lichen ascospores. J. Hattori Bot. Lab. 1998, 85, 267–278. [Google Scholar]
- Yoshimura, I.; Yamamoto, Y.; Nakano, T.; Finnie, J. Isolation and culture of lichen photobionts and mycobionts. In Methods in Lichenology; Kranner, I., Becknett, R., Varma, A., Eds.; Springer: Heidelberg, Germany, 2001; pp. 3–33. [Google Scholar]
- Yamamoto, Y.; Mizuguchi, R.; Yamada, Y. Tissue cultures of Usnea rubescens and Ramalina yasudae and production of usnic acid in their cultures. Agric. Biol. Chem. 1985, 49, 3347–3348. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Kinoshita, Y.; Yoshimura, I. Culture of thallus fragments and redifferentiation of lichens. In Methods in Lichenology; Kranner, I., Becknett, R., Varma, A., Eds.; Springer: Heiderberg, Germany, 2001; pp. 34–46. [Google Scholar]
- Yamamoto, Y.; Miura, Y.; Higuchi, M.; Kinoshita, Y.; Yoshimura, I. Using lichen tissue cultures in modern biology. Bryologist 1993, 96, 384. [Google Scholar] [CrossRef]
- Kinoshita, Y.; Yamamoto, Y.; Okuyama, E.; Yamazaki, M. Screening in natural thalli and cultured mycobionts of lichens for inhibitory activity of monoamine oxidase. Lichenology 2006, 5, 1–6. [Google Scholar]
- Verma, N.; Behera, B.C. In Vitro culture of lichen partners: Need and Implications. Recent Adv. Lichenol. 2015, 147–159. [Google Scholar] [CrossRef]
- Behera, B.C.; Adawadkar, B.; Makhija, U. Tissue culture of Bulbothrix setschwanensis (lichenized ascomycetes) in vitro. Curr. Sci. 2000, 78, 781–783. [Google Scholar]
- Behera, B.C.; Verma, N.; Sonone, A.; Makhija, U. Retracted: Tissue culture of some lichens and screening of their antioxidant, antityrosinase and antibacterial properties. Phytother. Res. 2007, 21, 1159–1170. [Google Scholar] [CrossRef]
- Kono, M.; Kon, Y.; Ohmura, Y.; Satta, Y.; Terai, Y. In vitro resynthesis of lichenization reveals the genetic background of symbiosis-specific fungal-algal interaction in Usnea hakonensis. BMC Genom. 2020, 21, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Tanahashi, T.; Kuroishi, M.; Kuwahara, A.; Nagakura, N.; Hamada, N. Four phenols from the cultured lichen mycobiont of Graphis scripta var. pulverulenta. Chem. Pharm. Bull. 1997, 45, 1183–1185. [Google Scholar] [CrossRef] [Green Version]
- Miyagawa, H.; Hamada, N.; Sato, M.; Eeno, T. Pigments from the cultured lichen mycobionts of Graphis scripta and G. desquamescens. Phytochemistry 1994, 36, 1319–1322. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Matsubara, H.; Kinoshita, Y.; Kinoshita, K.; Koyama, K.; Takahashi, K.; Ahmadjiam, V.; Kurokawa, T.; Yoshimura, I. Naphthazarin derivatives from cultures of the lichen Cladonia cristatella. Phytochemistry 1996, 43, 1239–1242. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kinoshita, Y.; Thor, G.R.; Hasumi, M.; Kinoshita, K.; Koyama, K.; Takahashi, K.; Yoshimura, I. Isofuranonaphthoquinone derivatives from cultures of the lichen Arthonia cinnabarina (DC.) Wallr. Phytochemistry 2002, 60, 741–745. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kinoshita, Y.; Kinoshita, K.; Koyama, K.; Takahashi, K. A new zearalenone derivative from cultures of the lichen Baeomyces placophyllus. J. Hattori Bot. Lab. 2002, 92, 285–289. [Google Scholar]
- Kinoshita, K.; Yamamoto, Y.; Koyama, K.; Takahashi, K.; Yoshimura, I. Novel fluorescent Isoquinoline pigments, panaefluoroline A–C from cultured mycobiont of a lichen, Amygdalaria panaeola. Tetrahedron Lett. 2003, 44, 8009–8011. [Google Scholar] [CrossRef]
- Tanahashi, T.; Takenaka, Y. Sesquiterpene derivatives from cultured lichen mycobionts of Diorygma sp. Planta Med. 2015, 81, PM_55. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, J.; Taylor, S.B. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Mühlbauer, A.; Triebel, D.; Peršoh, D.; Wollweber, H.; Seip, S.; Stadler, M. Macrocarpones, novel metabolites from stromata of Hypoxylon macrocarpum, and new evidence on the chemotaxonomy of Hypoxylon species. Mycol. Prog. 2002, 1, 235–248. [Google Scholar] [CrossRef]
- Stadler, M.; Ju, Y.-M.; Rogers, J.D. Chemotaxonomy of Entonaema, Rhopalostroma and other Xylariaceae. Mycol. Res. 2004, 108, 239–256. [Google Scholar] [CrossRef] [PubMed]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Hawksworth, D.L. Problems and prospects in the systematics of the Ascomycotina. Proc. Indian Acad. Sci. 1985, 94, 319–339. [Google Scholar]
- Hawksworth, D.L. Fungal diversity and its implications for genetic resource collections. Stud. Mycol. 2004, 50, 9–17. [Google Scholar]
- Humber, R.A. Fungi: Identification. In Manual of Techniques in Insect Pathology; Academic Press: Cambridge, MA, USA, 1997; pp. 153–185. [Google Scholar]
- Seifert, K.A.; Rossman, A.Y. How to describe a new fungal species. IMA Fungus 2010, 1, 109–111. [Google Scholar] [CrossRef] [PubMed]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code); Regnum Vegetabile 159. Glashütten; Koeltz Botanical Books: Glahutten, Germany, 2018. [Google Scholar]
- Abd-Elsalam, K.; Yassin, M.A.; Moslem, M.A.; Bahkali, A.; de Wit, P.; McKenzie, E.H.C.; Stephenson, S.L.; Cai, L.; Hyde, K.D. Culture collections, the new herbaria for fungal pathogens. Fungal Divers. 2010, 45, 21–32. [Google Scholar] [CrossRef]
- Damm, U.; Cannon, P.; Woudenberg, J.H.; Crous, P. The Colletotrichum acutatum species complex. Stud. Mycol. 2012, 73, 37–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariyawansa, H.; Hawksworth, D.L.; Hyde, K.D.; Jones, E.B.G.; Maharachchikumbura, S.; Manamgoda, D.S.; Thambugala, K.M.; Udayanga, D.; Camporesi, E.; Daranagama, A.; et al. Epitypification and neotypification: Guidelines with appropriate and inappropriate examples. Fungal Divers. 2014, 69, 57–91. [Google Scholar] [CrossRef]
- Tejesvi, M.V.; Kajula, M.; Mattila, S.; Pirttilä, A.M. Bioactivity and genetic diversity of endophytic fungi in Rhododendron tomentosum Harmaja. Fungal Divers. 2011, 47, 97–107. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; Bhat, J.D.; Mortimer, P.E.; Xu, J.; Promputtha, I.; Doilom, M.; Jun-Bo, Y.; Tang, A.M.C.; Karunarathna, S.C. Identification of endophytic fungi from leaves of Pandanaceae based on their morphotypes and DNA sequence data from southern Thailand. MycoKeys 2018, 33, 25–67. [Google Scholar] [CrossRef] [PubMed]
- Crahay, C.; Declerck, S.; Colpaert, J.V.; Pigeon, M.; Munaut, F. Viability of ectomycorrhizal fungi following cryopreservation. Fungal Biol. 2013, 117, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Itazaki, H.; Nagashima, K.; Kawamura, Y.; Matsumoto, K.; Nakai, H.; Terui, Y. Cinatrins, a novel family of phospholipase A2 inhibitors. I. Taxonomy and fermentation of the producing culture; isolation and structures of cinatrins. J. Antibiot. 1992, 45, 38–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bills, G.; Polishook, J.D. Abundance and diversity of microfungi in leaf litter of a lowland rain forest in Costa Rica. Mycologia 1994, 86, 187. [Google Scholar] [CrossRef]
- Hajek, A.E.; Humber, R.A.; Griggs, M.H. Decline in virulence of Entomophaga maimaiga (Zygomycetes: Entomophthorales) with repeated in vitro subculture. J. Invertebr. Pathol. 1990, 56, 91–97. [Google Scholar] [CrossRef]
- Krokene, P.; Solheim, H. Loss of pathogenicity in the blue-stain fungus Ceratocystis polonica. Plant Pathol. 2001, 50, 497–502. [Google Scholar] [CrossRef]
- Kashket, E.R.; Cao, Z.Y. Clostridial strain degeneration. FEMS Microbiol. Rev. 1995, 17, 307–315. [Google Scholar] [CrossRef]
- Forin, N.; Nigris, S.; Voyron, S.; Girlanda, M.; Vizzini, A.; Casadoro, G.; Baldan, B. Next generation sequencing of ancient fungal specimens: The Case of the Saccardo mycological herbarium. Front. Ecol. Evol. 2018, 6, 129. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Schumacher, R.K.; Wood, A.R.; Groenewald, J.Z. The genera of fungi–G5: Arthrinium, Ceratosphaeria, Dimerosporiopsis, Hormodochis, Lecanostictopsis, Lembosina, Neomelanconium, Phragmotrichum, Pseudomelanconium, Rutola, and Trullula. Fungal Syst. Evol. 2020, 5, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Forin, N.; Vizzini, A.; Nigris, S.; Ercole, E.; Voyron, S.; Girlanda, M.; Baldan, B. Illuminating type collections of nectriaceous fungi in Saccardo’s fungarium. Pers. Mol. Phylogeny Evol. Fungi 2020, 45, 221–249. [Google Scholar] [CrossRef]
- Tedersoo, L.; Liiv, I.; Kivistik, P.A.; Anslan, S.; Kõljalg, U.; Bahram, M. Genomics and metagenomics technologies to recover ribosomal DNA and single-copy genes from old fruit-body and ectomycorrhiza specimens. MycoKeys 2016, 13, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibbett, D.S.; Fisher, M. Phylogenetic species recognition and species concepts in fungi. Fungal Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booth, C. Methods in Microbiology; Academic Press INC LTD: London, UK, 1971. [Google Scholar]
- Polishook, J.D.; Bills, G.; Lodge, D.J. Microfungi from decaying leaves of two rain forest trees in Puerto Rico. J. Ind. Microbiol. Biotechnol. 1996, 17, 284–294. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Hyde, K.D. Succession: Where do we go from here? Fungal Divers. 2002, 10, 241–253. [Google Scholar]
- Promputtha, I.; Lumyong, S.; Dhanasekaran, V.; McKenzie, E.H.C.; Hyde, K.D.; Jeewon, R. A Phylogenetic evaluation of whether endophytes become saprotrophs at host senescence. Microb. Ecol. 2007, 53, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Promputtha, I.; Lumyong, S.; Lumyong, P.; McKenzie, E.C.; Hyde, K.D. Fungal succession on senescent leaves of Manglietia garrettii in Doi Suthep-Pui National Park, northern Thailand. Fungal Divers. 2002, 10, 89–100. [Google Scholar]
- Hawksworth, D.L. Microbial collections as a tool in biodiversity and biosystematic research. In Culture Collections to Improve the Quality of Life; Samson, R.A., Stalpers, J.A., van der Mei, D., Stouthamer, A.H., Eds.; Centraalbureau voor Schimmelcultures and World Federation of Culture Collections: Baarn, The Netherlands, 1996; pp. 26–35. [Google Scholar]
- Quaedvlieg, W.; Binder, M.; Groenewald, J.; Summerell, B.; Carnegie, A.; Burgess, T.; Crous, P. Introducing the Consolidated Species Concept to resolve species in the Teratosphaeriaceae. Pers. Mol. Phylogeny Evol. Fungi 2014, 33, 1–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lücking, R.; Aime, M.C.; Robbertse, B.; Miller, A.N.; Ariyawansa, H.A.; Aoki, T.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Unambiguous identification of fungi: Where do we stand and how accurate and precise is fungal DNA barcoding? IMA Fungus 2020, 11, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D.L. Lessons from 50 years describing and classifying fungi. Kavaka 2020, 55, 1–11. [Google Scholar] [CrossRef]
- Hongsanan, S.; Jeewon, R.; Purahong, W.; Xie, N.; Liu, J.K.; Jayawardena, R.S.; Ekanayaka, A.H.; Dissanayake, A.; Raspé, O.; Hyde, K.D.; et al. Can we use environmental DNA as holotypes? Fungal Divers. 2018, 92, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Nilsson, R.H.; James, T.Y.; Dai, Y.; Townsend, J.P. Future perspectives and challenges of fungal systematics in the age of big data. Biol. Microfungi 2016, 25–46. [Google Scholar] [CrossRef]
- Mitchell, J.I.; Zuccaro, A. Sequences, the environment and fungi. Mycologist 2006, 20, 62–74. [Google Scholar] [CrossRef]
- Seifert, K.A. Progress towards DNA barcoding of fungi. Mol. Ecol. Resour. 2009, 9, 83–89. [Google Scholar] [CrossRef]
- Parr, C.S.; Guralnick, R.; Cellinese, N.; Page, R. Evolutionary informatics: Unifying knowledge about the diversity of life. Trends Ecol. Evol. 2012, 27, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Grossart, H.-P.; Wurzbacher, C.; James, T.Y.; Kagami, M. Discovery of dark matter fungi in aquatic ecosystems demands a reappraisal of the phylogeny and ecology of zoosporic fungi. Fungal Ecol. 2016, 19, 28–38. [Google Scholar] [CrossRef] [Green Version]
- Page, R.D.M. DNA barcoding and taxonomy: Dark taxa and dark texts. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150334. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; Smith, M.E. Ectomycorrhizal Fungal Lineages: Detection of four new groups and notes on consistent recognition of ectomycorrhizal taxa in High-Throughput Sequencing studies. In Biogeography of Mycorrhizal Symbiosis; Tedersoo, L., Ed.; Springer: Cham, Switzerland, 2017; pp. 125–142. [Google Scholar] [CrossRef]
- Ryberg, M.; Nilsson, R.H. New light on names and naming of dark taxa. MycoKeys 2018, 30, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Sanger, F.; Air, G.; Barrell, B.G.; Brown, N.L.; Coulson, A.R.; Fiddes, J.C.; Hutchison, C.A.; Slocombe, P.M.; Smith, M.A. Nucleotide sequence of bacteriophage φX174 DNA. Nature 1977, 265, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shokralla, S.; Spall, J.L.; Gibson, J.; Hajibabaei, M. Next-generation sequencing technologies for environmental DNA research. Mol. Ecol. 2012, 21, 1794–1805. [Google Scholar] [CrossRef]
- Margulies, M.; Egholm, M.; Altman, W.E.; Attiya, S.; Bader, J.; Bemben, L.A.; Berka, J.; Braverman, M.S.; Chen, Y.-J.; Chen, Z.; et al. Genome sequencing in microfabricated high-density picolitre reactors. Nature 2005, 437, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Shendure, J.; Porreca, G.J.; Reppas, N.B.; Lin, X.; McCutcheon, J.P.; Rosenbaum, A.M.; Wang, M.D.; Zhang, K.; Mitra, R.D.; Church, G.M. Accurate Multiplex Polony Sequencing of an evolved bacterial genome. Science 2005, 309, 1728–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, S.C. Next-generation sequencing transforms today’s biology. Nat. Methods 2008, 5, 16–18. [Google Scholar] [CrossRef] [PubMed]
- Shendure, J.; Ji, H. Next-generation DNA sequencing. Nat. Biotechnol. 2008, 26, 1135–1145. [Google Scholar] [CrossRef] [PubMed]
- Bartnicki-Garcia, S. The cell wall in fungal evolution. In Evolutionary Biology of the Fungi; Rayner, A.D.M., Brasier, C.M., Moore, D., Eds.; Cambridge University Press: New York, NY, USA, 1987. [Google Scholar]
- Handelsman, J.; Rondon, M.R.; Brady, S.F.; Clardy, J.; Goodman, R.M. Molecular biological access to the chemistry of unknown soil microbes: A new frontier for natural products. Chem. Biol. 1998, 5, R245–R249. [Google Scholar] [CrossRef] [Green Version]
- Eisen, J.A. Environmental Shotgun Sequencing: Its potential and challenges for studying the hidden world of microbes. PLoS Biol. 2007, 5, e82. [Google Scholar] [CrossRef] [PubMed]
- Ghannoum, M.A.; Jurevic, R.J.; Mukherjee, P.K.; Cui, F.; Sikaroodi, M.; Naqvi, A.; Gillevet, P.M. Characterization of the oral fungal microbiome (Mycobiome) in healthy individuals. PLoS Pathog. 2010, 6, e1000713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gweon, H.S.; Oliver, A.; Taylor, J.; Booth, T.; Gibbs, M.; Read, D.S.; Griffiths, R.I.; Schonrogge, K. PIPITS: An automated pipeline for analyses of fungal internal transcribed spacer sequences from the Illumina sequencing platform. Methods Ecol. Evol. 2015, 6, 973–980. [Google Scholar] [CrossRef]
- White, J.R.; Maddox, C.; White, O.; Angiuoli, S.V.; Fricke, W.F. CloVR-ITS: Automated internal transcribed spacer amplicon sequence analysis pipeline for the characterization of fungal microbiota. Microbiome 2013, 1, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anslan, S.; Bahram, M.; Hiiesalu, I.; Tedersoo, L. PipeCraft: Flexible open-source toolkit for bioinformatics analysis of custom high-throughput amplicon sequencing data. Mol. Ecol. Resour. 2017, 17, e234–e240. [Google Scholar] [CrossRef] [PubMed]
- Donovan, P.; Gonzalez, G.; Higgins, D.G.; Butler, G.; Ito, K. Identification of fungi in shotgun metagenomics datasets. PLoS ONE 2018, 13, e0192898. [Google Scholar] [CrossRef] [PubMed]
- Madhavan, A.; Sindhu, R.; Parameswaran, B.; Sukumaran, R.K.; Pandey, A. Metagenome Analysis: A powerful tool for enzyme bioprospecting. Appl. Biochem. Biotechnol. 2017, 183, 636–651. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Anslan, S.; Bahram, M.; Wurzbacher, C.; Baldrian, P.; Tedersoo, L. Mycobiome diversity: High-throughput sequencing and identification of fungi. Nat. Rev. Genet. 2018, 17, 95–109. [Google Scholar] [CrossRef]
- Brumfield, K.D.; Hasan, N.A.; Leddy, M.B.; Cotruvo, J.A.; Rashed, S.M.; Colwell, R.R.; Huq, A. A comparative analysis of drinking water employing metagenomics. PLoS ONE 2020, 15, e0231210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderón-Ezquerro, M.D.C.; Serrano-Silva, N.; Brunner-Mendoza, C. Aerobiological study of bacterial and fungal community composition in the atmosphere of Mexico City throughout an annual cycle. Environ. Pollut. 2021, 278, 116858. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.; Rajput, V.; Shah, M.; Yadav, R.; Sarode, P.; Dastager, S.G.; Dharne, M.S.; Khairnar, K. Deciphering taxonomic and functional diversity of fungi as potential bioindicators within confluence stretch of Ganges and Yamuna Rivers, impacted by anthropogenic activities. Chemosphere 2020, 252, 126507. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Williams, L.J.; Vincent, J.B.; Stefanski, A.; Cavender-Bares, J.; Messier, C.; Paquette, A.; Gravel, D.; Reich, P.; Kennedy, P.G. Ectomycorrhizal fungal diversity and saprotrophic fungal diversity are linked to different tree community attributes in a field-based tree experiment. Mol. Ecol. 2016, 25, 4032–4046. [Google Scholar] [CrossRef]
- Yang, T.; Adams, J.M.; Shi, Y.; He, J.S.; Jing, X.; Chen, L.; Chu, H. Plant diversity and productivity drive soil fungal richness in natural grasslands of the Tibetan Plateau. New Phytol. 2017, 215, 756–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, R.A.; Noverr, M. Fungal interactions with the human host: Exploring the spectrum of symbiosis. Curr. Opin. Microbiol. 2017, 40, 58–64. [Google Scholar] [CrossRef]
- Hager, C.L.; Ghannoum, M.A. The mycobiome: Role in health and disease, and as a potential probiotic target in gastrointestinal disease. Dig. Liver Dis. 2017, 49, 1171–1176. [Google Scholar] [CrossRef] [PubMed]
- Lücking, R.; Hawksworth, D.L. Formal description of sequence-based voucherless Fungi: Promises and pitfalls, and how to resolve them. IMA Fungus 2018, 9, 143–165. [Google Scholar] [CrossRef] [Green Version]
- Lepere, C.; Demura, M.; Kawachi, M.; Romac, S.; Probert, I.; Vaulot, D. Whole-genome amplification (WGA) of marine photosynthetic eukaryote populations. FEMS Microbiol. Ecol. 2011, 76, 513–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tedersoo, L.; Bahram, M.; Puusepp, R.; Nilsson, H.; James, T.Y. Novel soil-inhabiting clades fill gaps in the fungal tree of life. Microbiome 2017, 5, 1–10. [Google Scholar] [CrossRef]
- Tedersoo, L.; Anslan, S.; Bahram, M.; Kõljalg, U.; Abarenkov, K. Identifying the ‘unidentified’ fungi: A global-scale long-read third-generation sequencing approach. Fungal Divers. 2020, 103, 273–293. [Google Scholar] [CrossRef]
- Martin, F.; Kohler, A.; Murat, C.; Balestrini, R.; Coutinho, P.M.; Jaillon, O.; Montanini, B.; Morin, E.; Noel, B.; Percudani, R.; et al. Périgord black truffle genome uncovers evolutionary origins and mechanisms of symbiosis. Nature 2010, 464, 1033–1038. [Google Scholar] [CrossRef]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; van Bodegom, P.; Bengtsson-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H.; et al. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef]
- Bahram, M.; Vanderpool, D.; Pent, M.; Hiltunen, M.; Ryberg, M. The genome and microbiome of a dikaryotic fungus (Inocybe terrigena, Inocybaceae) revealed by metagenomics. Environ. Microbiol. Rep. 2018, 10, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Nilsson, H.; Abarenkov, K.; Jairus, T.; Sadam, A.; Saar, I.; Bahram, M.; Bechem, E.; Chuyong, G.; Kõljalg, U. 454 Pyrosequencing and Sanger sequencing of tropical mycorrhizal fungi provide similar results but reveal substantial methodological biases. New Phytol. 2010, 188, 291–301. [Google Scholar] [CrossRef]
- Cuadros-Orellana, S.; Leite, L.R.; Smith, A.; Medeiros, J.D.; Badotti, A.; Fonseca, P.; Vaz, A.B.; Oliveira, G.; Góes-Neto, A. Assessment of fungal diversity in the environment using metagenomics: A decade in review. Fungal Genom. Biol. 2013, 3, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Escobar-Zepeda, A.; de León, A.V.-P.; Sanchez-Flores, A.; Escobar-Zepeda, A.; de León, A.V.-P.; Sanchez-Flores, A. The Road to metagenomics: From microbiology to DNA sequencing technologies and bioinformatics. Front. Genet. 2015, 6, 348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dijk, E.L.; Auger, H.; Jaszczyszyn, Y.; Thermes, C. Ten years of next-generation sequencing technology. Trends Genet. 2014, 30, 418–426. [Google Scholar] [CrossRef]
- Wooley, J.C.; Godzik, A.; Friedberg, I. A Primer on Metagenomics. PLoS Comput. Biol. 2010, 6, e1000667. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth & Bisby’s Dictionary of the Fungi, 10th ed.; CABI: Wallingford, UK, 2008. [Google Scholar]
- Raghuwanshi, R.; Singh, S.; Aamir, M.; Saxena, A.; Gupta, V.K.; Upadhyay, R.S. Biology of microfungi in biofuel. Bioenergy Res. 2016, 543–571. [Google Scholar] [CrossRef]
- Andersson, A.F.; Lindberg, M.; Jakobsson, H.; Bäckhed, F.; Nygren, P.; Engstrand, L. Comparative analysis of human gut microbiota by barcoded pyrosequencing. PLoS ONE 2008, 3, e2836. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; DiBaise, J.K.; Zuccolo, A.; Kudrna, D.; Braidotti, M.; Yu, Y.; Parameswaran, P.; Crowell, M.; Wing, R.; Rittmann, B.E.; et al. Human gut microbiota in obesity and after gastric bypass. Proc. Natl. Acad. Sci. USA 2009, 106, 2365–2370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huseyin, C.; O’Toole, P.; Cotter, P.; Scanlan, P. Forgotten fungi—the gut mycobiome in human health and disease. FEMS Microbiol. Rev. 2017, 41, 479–511. [Google Scholar] [CrossRef] [Green Version]
- Shivaji, S.; Jayasudha, R.; Prashanthi, G.S.; Chakravarthy, S.K.; Sharma, S. The human ocular surface fungal microbiome. Investig. Opthalmol. Vis. Sci. 2019, 60, 451–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berruti, A.; Borriello, R.; Orgiazzi, A.; Barbera, A.C.; Lumini, E.; Bianciotto, E.L.A.V. Arbuscular mycorrhizal fungi and their value for ecosystem management. In Biodiversity: The Dynamic Balance of the Planet; Grillo, O., Ed.; InTech: Rijeta, Croacia, 2014; pp. 159–191. [Google Scholar] [CrossRef] [Green Version]
- Frąc, M.; Hannula, S.E.; Bełka, M.; Jędryczka, M. Fungal biodiversity and their role in soil health. Front. Microbiol. 2018, 9, 707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellemain, E.; Davey, M.; Kauserud, H.; Epp, L.S.; Boessenkool, S.; Coissac, E.; Geml, J.; Edwards, M.; Willerslev, E.; Gussarova, G.; et al. Fungal palaeodiversity revealed using high-throughput metabarcoding of ancient DNA from arctic permafrost. Environ. Microbiol. 2012, 15, 1176–1189. [Google Scholar] [CrossRef]
- Buée, M.; de Boer, W.; Martin, F.; van Overbeek, L.; Jurkevitch, E. The rhizosphere zoo: An overview of plant-associated communities of microorganisms, including phages, bacteria, archaea, and fungi, and of some of their structuring factors. Plant Soil 2009, 321, 189–212. [Google Scholar] [CrossRef]
- Jumpponen, A.; Jones, K.L. Massivelyparallel454 sequencing indicates hyper diverse fungal communities in temperate Quercus macrocarpa phyllosphere. New Phytol. 2009, 184, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Öpik, M.; Metsis, M.; Daniell, T.J.; Zobel, M.; Moora, M. Large-scale parallel 454 sequencing reveals host ecological group specificity of arbuscular mycorrhizal fungi in a boreonemoral forest. New Phytol. 2009, 184, 424–437. [Google Scholar] [CrossRef]
- Jumpponen, A.R.; Jones, K.L.; David Mattox, J.; Yaege, C. Massively parallel 454 sequencing of fungal communities in Quercus spp. ectomycorrhizas indicates seasonal dynamics in urban and rural sites. Mol. Ecol. 2010, 19, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Hibbett, D.S.; Ohman, A.; Glotzer, D.; Nuhn, M.; Kirk, P.; Nilsson, R.H. Progress in molecular and morphological taxon discovery in Fungi and options for formal classification of environmental sequences. Fungal Biol. Rev. 2011, 25, 38–47. [Google Scholar] [CrossRef]
- Schadt, C.W.; Martin, A.P.; Lipson, D.A.; Schmidt, S.K. Seasonal dynamics of previously unknown fungal lineages in tundra soils. Science 2003, 301, 1359–1361. [Google Scholar] [CrossRef] [Green Version]
- Rosling, A.; Cox, F.; Cruz-Martinez, K.; Ihrmark, K.; Grelet, G.-A.; Lindahl, B.D.; Menkis, A.; James, T.Y. Archaeorhizomycetes: Unearthing an ancient class of ubiquitous soil fungi. Science 2011, 333, 876–879. [Google Scholar] [CrossRef]
- Menkis, A.; Urbina, H.; James, T.Y.; Rosling, A. Archaeorhizomyces borealis sp. nov. and a sequence-based classification of related soil fungal species. Fungal Biol. 2014, 118, 943–955. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.D.M.; Richards, T.; Hawksworth, D.L.; Bass, D. Validation and justification of the phylum name Cryptomycota phyl. nov. IMA Fungus 2011, 2, 173–175. [Google Scholar] [CrossRef]
- Jones, M.D.M.; Forn, I.; Gadelha, C.; Egan, M.J.; Bass, D.; Massana, R.; Richards, T. Discovery of novel intermediate forms redefines the fungal tree of life. Nature 2011, 474, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Livermore, J.A.; Mattes, T.E. Phylogenetic detection of novel Cryptomycota in an Iowa (United States) aquifer and from previously collected marine and freshwater targeted high-throughput sequencing sets. Environ. Microbiol. 2013, 15, 2333–2341. [Google Scholar] [CrossRef]
- De Beer, Z.W.; Marincowitz, S.; Duong, T.A.; Kim, J.-J.; Rodrigues, A.; Wingfield, M.J. Hawksworthiomyces gen. nov. (Ophiostomatales), illustrates the urgency for a decision on how to name novel taxa known only from environmental nucleic acid sequences (ENAS). Fungal Biol. 2016, 120, 1323–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrini, O. Fungal endophytes of tree leaves. In Microbial Ecology of Leaves; Springer: New York, NY, USA, 1991; pp. 179–197. [Google Scholar]
- Malloch, D.W.; Pirozynski, K.A.; Raven, P.H. Ecological and evolutionary significance of mycorrhizal symbioses in vascular plants (A Review). Proc. Natl. Acad. Sci. USA 1980, 77, 2113–2118. [Google Scholar] [CrossRef] [Green Version]
- Petrini, O.; Stone, J.; Carroll, F.E. Endophytic fungi in evergreen shrubs in western Oregon: A preliminary study. Can. J. Bot. 1982, 60, 789–796. [Google Scholar] [CrossRef]
- Rodrigues, K.F.; Samuels, G.J. Preliminary study of endophytic fungi in a tropical palm. Mycol. Res. 1990, 94, 827–830. [Google Scholar] [CrossRef]
- Meletiadis, J.; Meis, J.F.G.M.; Mouton, J.W.; Verweij, P.E. Analysis of growth characteristics of filamentous fungi in different nutrient media. J. Clin. Microbiol. 2001, 39, 478–484. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.K.; Sharma, V.K.; Kumar, J.; Mishra, A.; Verma, S.K.; Sieber, T.N.; Kharwar, R.N. Diversity of endophytic mycobiota of tropical tree Tectona grandis Linn.f.: Spatiotemporal and tissue type effects. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, X.; Xu, F.; Qin, D.; Gao, T.; Shen, W.; Zuo, S.; Yu, B.; Xu, J.; Peng, Y.; Dong, J. Diversity and bioactivities of fungal endophytes from Distylium chinense, a rare waterlogging tolerant plant endemic to the Three Gorges Reservoir. BMC Microbiol. 2019, 19, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.-L. Effect of Host, Environment and fungal growth on fungal leaf endophyte communities in Taiwan. J. Fungi 2020, 6, 244. [Google Scholar] [CrossRef]
- Arnold, A.E.; Henk, D.A.; Eells, R.L.; Lutzoni, F.; Vilgalys, R. Diversity and phylogenetic affinities of foliar fungal endophytes in loblolly pine inferred by culturing and environmental PCR. Mycologia 2007, 99, 185–206. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Purahong, W.; Wubet, T.; Hyde, K.D.; Zhang, W.; Xu, H.; Zhang, G.; Fu, C.; Liu, M.; Xing, Q.; et al. Direct comparison of culture-dependent and culture-independent molecular approaches reveal the diversity of fungal endophytic communities in stems of grapevine (Vitis vinifera). Fungal Divers. 2018, 90, 85–107. [Google Scholar] [CrossRef]
- Parmar, S.; Li, Q.; Wu, Y.; Li, X.; Yan, J.; Sharma, V.K.; Wei, Y.; Li, H. Endophytic fungal community of Dysphania ambrosioides from two heavy metal-contaminated sites: Evaluated by culture-dependent and culture-independent approaches. Microb. Biotechnol. 2018, 11, 1170–1183. [Google Scholar] [CrossRef]
- Luypaert, T.; Hagan, J.G.; McCarthy, M.L.; Poti, M. Status of marine biodiversity in the Anthropocene. In YOUMARES 9—The Oceans: Our Research, Our Future; Jungblut, S., Liebich, V., Bode-Dalby, M., Eds.; Springer: Cham, Switzerland, 2020; pp. 57–82. [Google Scholar] [CrossRef] [Green Version]
- Bubnova, E.N.; Georgieva, M.; Grum-Grzhimailo, O.A. Method for isolation and enumeration of fungi developing in marine sediments. Microbiology 2018, 87, 777–782. [Google Scholar] [CrossRef]
- Overy, D.P.; Rämä, T.; Oosterhuis, R.; Walker, A.K.; Pang, K.-L. The neglected marine fungi, sensu stricto, and their isolation for natural products’ discovery. Mar. Drugs 2019, 17, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghukumar, S. Methods to Study Marine Fungi. In Fungi in Coastal and Oceanic Marine Ecosystems; Springer: Cham, Switzerland, 2017; pp. 323–343. [Google Scholar] [CrossRef]
- Singh, P.; Raghukumar, C.; Verma, P.; Shouche, Y. Fungal community analysis in the deep-sea sediments of the Central Indian basin by culture-independent approach. Microb. Ecol. 2010, 61, 507–517. [Google Scholar] [CrossRef]
- Luo, Y.; Wei, X.; Yang, S.; Gao, Y.H.; Luo, Z.H. Fungal diversity in deep-sea sediments from the Magellan seamounts as revealed by a metabarcoding approach targeting the ITS2 regions. Mycology 2020, 11, 214–229. [Google Scholar] [CrossRef]
- Zhang, L.; Kang, M.; Huang, Y.; Yang, L. Fungal communities from the calcareous deep-sea sediments in the Southwest India Ridge revealed by Illumina sequencing technology. World J. Microbiol. Biotechnol. 2016, 32, 78. [Google Scholar] [CrossRef]
- Grassle, J.F. Marine ecosystems. In Encyclopedia of Biodiversity; Levin, S.A., Ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 45–55. [Google Scholar] [CrossRef]
- Nagahama, T.; Nagano, Y. Cultured and uncultured fungal diversity in deep-sea environments. Prog. Mol. Subcell. Biol. 2011, 53, 173–187. [Google Scholar] [CrossRef]
- Xu, W.; Pang, K.-L.; Luo, Z.-H. High fungal diversity and abundance recovered in the deep-sea sediments of the Pacific Ocean. Microb. Ecol. 2014, 68, 688–698. [Google Scholar] [CrossRef]
- Nagano, Y.; Miura, T.; Nishi, S.; Lima, A.; Nakayama, C.R.; Pellizari, V.; Fujikura, K. Fungal diversity in deep-sea sediments associated with asphalt seeps at the Sao Paulo Plateau. Deep Sea Res. Part II Top. Stud. Oceanogr. 2017, 146, 59–67. [Google Scholar] [CrossRef]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Rossman, A.Y. Where are all the undescribed fungi? Phytopathology 1997, 87, 888–891. [Google Scholar] [CrossRef] [Green Version]
- Brock, P.; Döring, H.; Bidartondo, M.I. How to know unknown fungi: The role of a herbarium. New Phytol. 2008, 181, 719–724. [Google Scholar] [CrossRef] [Green Version]
- Bebber, D.P.; Carine, M.A.; Wood, J.R.I.; Wortley, A.H.; Harris, D.; Prance, G.T.; Davidse, G.; Paige, J.; Pennington, T.D.; Robson, N.K.B.; et al. Herbaria are a major frontier for species discovery. Proc. Natl. Acad. Sci. USA 2010, 107, 22169–22171. [Google Scholar] [CrossRef] [Green Version]
- Bruns, T.D.; Fogel, R.; Taylor, J.W. Amplification and sequencing of DNA from fungal herbarium specimens. Mycologia 1990, 82, 175–184. [Google Scholar] [CrossRef]
- Smith, R.L.; Sawbridge, T.; Mann, R.; Kaur, J.; May, T.W.; Edwards, J. Rediscovering an old foe: Optimised molecular methods for DNA extraction and sequencing applications for fungarium specimens of powdery mildew (Erysiphales). PLoS ONE 2020, 15, e0232535. [Google Scholar] [CrossRef]
- Daru, B.H.; Bowman, E.A.; Pfister, D.H.; Arnold, A.E. A novel proof of concept for capturing the diversity of endophytic fungi preserved in herbarium specimens. Philos. Trans. R. Soc. B Biol. Sci. 2018, 374, 20170395. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.L.; May, T.W.; Kaur, J.; Sawbridge, T.; Mann, R.; Edwards, J. Molecular data from up to 130-year-old herbarium specimens do not support the presence of cherry powdery mildew in Australia. Plant Pathol. 2020, 70, 689–698. [Google Scholar] [CrossRef]
- Larkin, P.M.; Lawson, K.L.; Contreras, D.A.; Le, C.Q.; Trejo, M.; Realegeno, S.; Hilt, E.E.; Chandrasekaran, S.; Garner, O.B.; Fishbein, G.A.; et al. Amplicon-based next-generation sequencing for detection of fungi in formalin-fixed, paraffin-embedded tissues: Correlation with histopathology and clinical applications. J. Mol. Diag. 2020, 22, 1287–1293. [Google Scholar] [CrossRef]
- Tedersoo, L.; Albertsen, M.; Anslan, S.; Callahan, B. Perspectives and benefits of high-throughput long-read sequencing in microbial ecology. Appl. Environ. Microbiol. 2021, 87, e00626-21. [Google Scholar] [CrossRef]
- Leinonen, R.; Sugawara, H.; Shumway, M. On behalf of the international nucleotide sequence database collaboration. The sequence read archive. Nucleic Acids Res. 2010, 39, D19–D21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kodama, Y.; Shumway, M.; Leinonen, R.; On behalf of the International Nucleotide Sequence Database Collaboration. The sequence read archive: Explosive growth of sequencing data. Nucleic Acids Res. 2011, 40, D54–D56. [Google Scholar] [CrossRef] [Green Version]
- Hawksworth, D.L.; Hibbett, D.S.; Kirk, P.M.; Lücking, R. (308–310) Proposals to permit DNA sequence data to serve as types of names of fungi. Taxon 2016, 65, 899–900. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [Green Version]
- Kõljalg, U.; Tedersoo, L.; Nilsson, H.; Abarenkov, K. Digital identifiers for fungal species. Science 2016, 352, 1182–1183. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, H.R.; Schigel, D.; Tedersoo, L.; Larsson, K.-H.; May, T.W.; Taylor, A.F.S.; Jeppesen, T.S.; Frøslev, T.G.; Lindahl, B.D.; et al. The taxon hypothesis paradigm—On the unambiguous detection and communication of taxa. Microorganisms 2020, 8, 1910. [Google Scholar] [CrossRef] [PubMed]
- Thines, M.; Crous, P.W.; Aime, M.C.; Aoki, T.; Cai, L.; Hyde, K.D.; Miller, A.N.; Zhang, N.; Stadler, M. Ten reasons why a sequence-based nomenclature is not useful for fungi anytime soon. IMA Fungus 2018, 9, 177–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawksworth, D. The fungal dimension of biodiversity: Magnitude, significance, and conservation. Mycol. Res. 1991, 95, 641–655. [Google Scholar] [CrossRef]
- Hawksworth, D.L. The magnitude of fungal diversity: The 1.5 million species estimate revisited. Mycol. Res. 2001, 105, 1422–1432. [Google Scholar] [CrossRef] [Green Version]
- Hawksworth, D.L. Global species numbers of fungi: Are tropical studies and molecular approaches contributing to a more robust estimate? Biodivers Conserv. 2012, 21, 2425–2433. [Google Scholar] [CrossRef]
- Rossman, A.Y. A strategy for an all-taxa inventory of fungal diversity. In Biodiversity and Terrestrial Ecosystems; Peng, C.I., Chou, C.H., Eds.; [Monograph Series No. 14]; Institute of Botany Academia Sinica: Taipei, Taiwan, 1994; pp. 169–194. [Google Scholar]
- Hyde, K.D.; Jeewon, R.; Chen, Y.J.; Bhunjun, C.S.; Calabon, M.S.; Jiang, H.B.; Lin, C.G.; Norphanphoun, C.; Sysouphanthong, P.; Pem, D.; et al. The numbers of fungi: Is the descriptive curve flattening? Fungal Divers. 2020, 103, 219–271. [Google Scholar] [CrossRef]
- Tennakoon, D.S.; Gentekaki, E.; Jeewon, R.; Kuo, C.H.; Promputtha, I.; Hyde, K.D. Life in leaf litter: Fungal community succession during decomposition. Mycosphere 2021, 12, 406–429. [Google Scholar]
- Hibbett, D.S.; Taylor, J.W. Fungal systematics: Is a new age of enlightenment at hand? Nat. Rev. Microbiol. 2013, 11, 129–133. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wijayawardene, N.N.; Bahram, M.; Sánchez-Castro, I.; Dai, D.-Q.; Ariyawansa, K.G.S.U.; Jayalal, U.; Suwannarach, N.; Tedersoo, L. Current Insight into Culture-Dependent and Culture-Independent Methods in Discovering Ascomycetous Taxa. J. Fungi 2021, 7, 703. https://doi.org/10.3390/jof7090703
Wijayawardene NN, Bahram M, Sánchez-Castro I, Dai D-Q, Ariyawansa KGSU, Jayalal U, Suwannarach N, Tedersoo L. Current Insight into Culture-Dependent and Culture-Independent Methods in Discovering Ascomycetous Taxa. Journal of Fungi. 2021; 7(9):703. https://doi.org/10.3390/jof7090703
Chicago/Turabian StyleWijayawardene, Nalin N., Mohammad Bahram, Iván Sánchez-Castro, Dong-Qin Dai, Kahandawa G. S. U. Ariyawansa, Udeni Jayalal, Nakarin Suwannarach, and Leho Tedersoo. 2021. "Current Insight into Culture-Dependent and Culture-Independent Methods in Discovering Ascomycetous Taxa" Journal of Fungi 7, no. 9: 703. https://doi.org/10.3390/jof7090703
APA StyleWijayawardene, N. N., Bahram, M., Sánchez-Castro, I., Dai, D. -Q., Ariyawansa, K. G. S. U., Jayalal, U., Suwannarach, N., & Tedersoo, L. (2021). Current Insight into Culture-Dependent and Culture-Independent Methods in Discovering Ascomycetous Taxa. Journal of Fungi, 7(9), 703. https://doi.org/10.3390/jof7090703