
Genetic Analyses of Saprolegnia Strains Isolated from Salmonid Fish of Different Geographic Origin Document the Connection between Pathogenicity and Molecular Diversity

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Saprolegnia Material
2.2. Amplified Fragment Length Polymorphism (AFLP) Analysis
2.3. Data Analysis of the AFLP
2.4. Sequencing of the ITS
2.5. Phylogenetic Analyses
2.6. Cluster Analyses
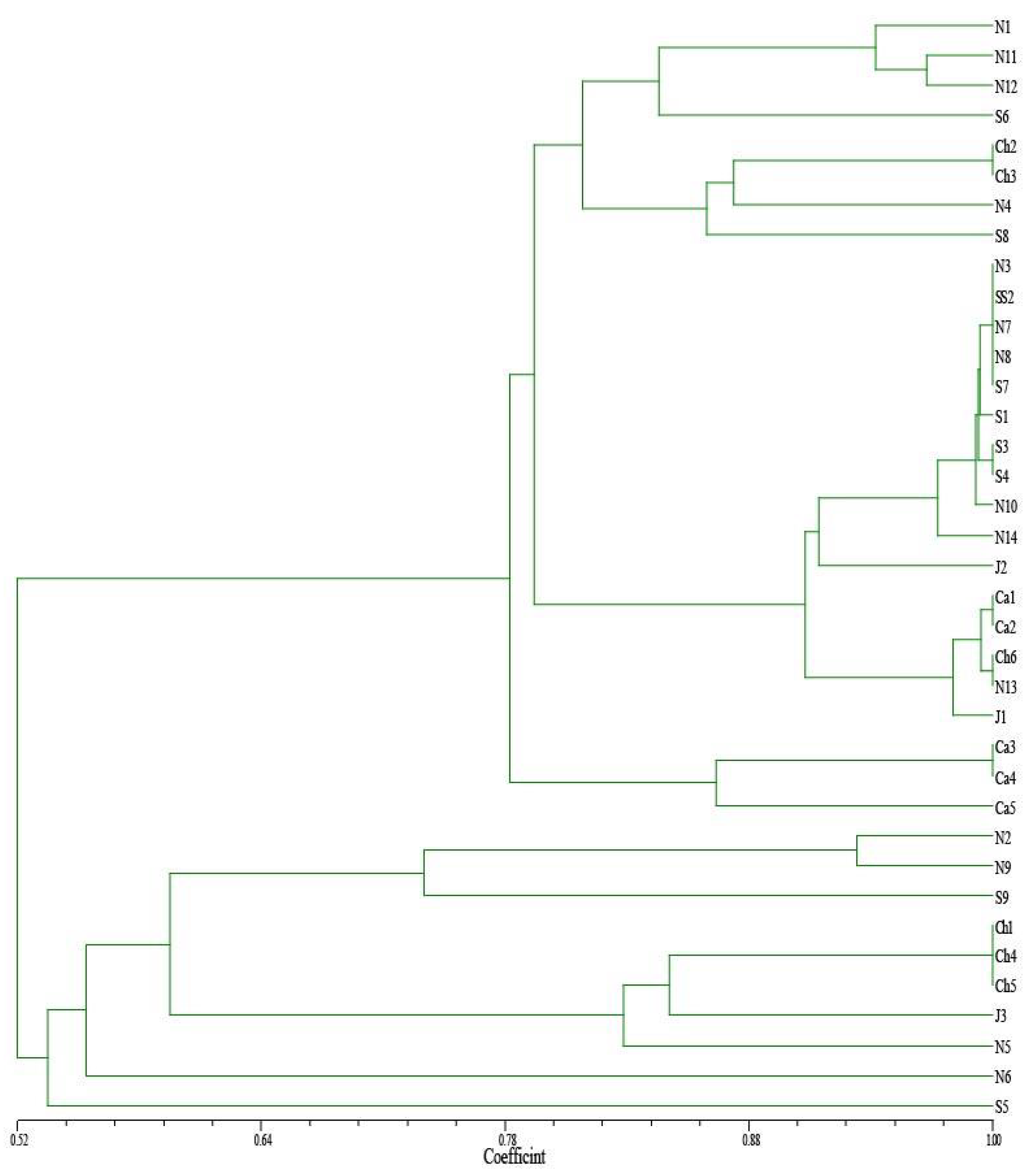
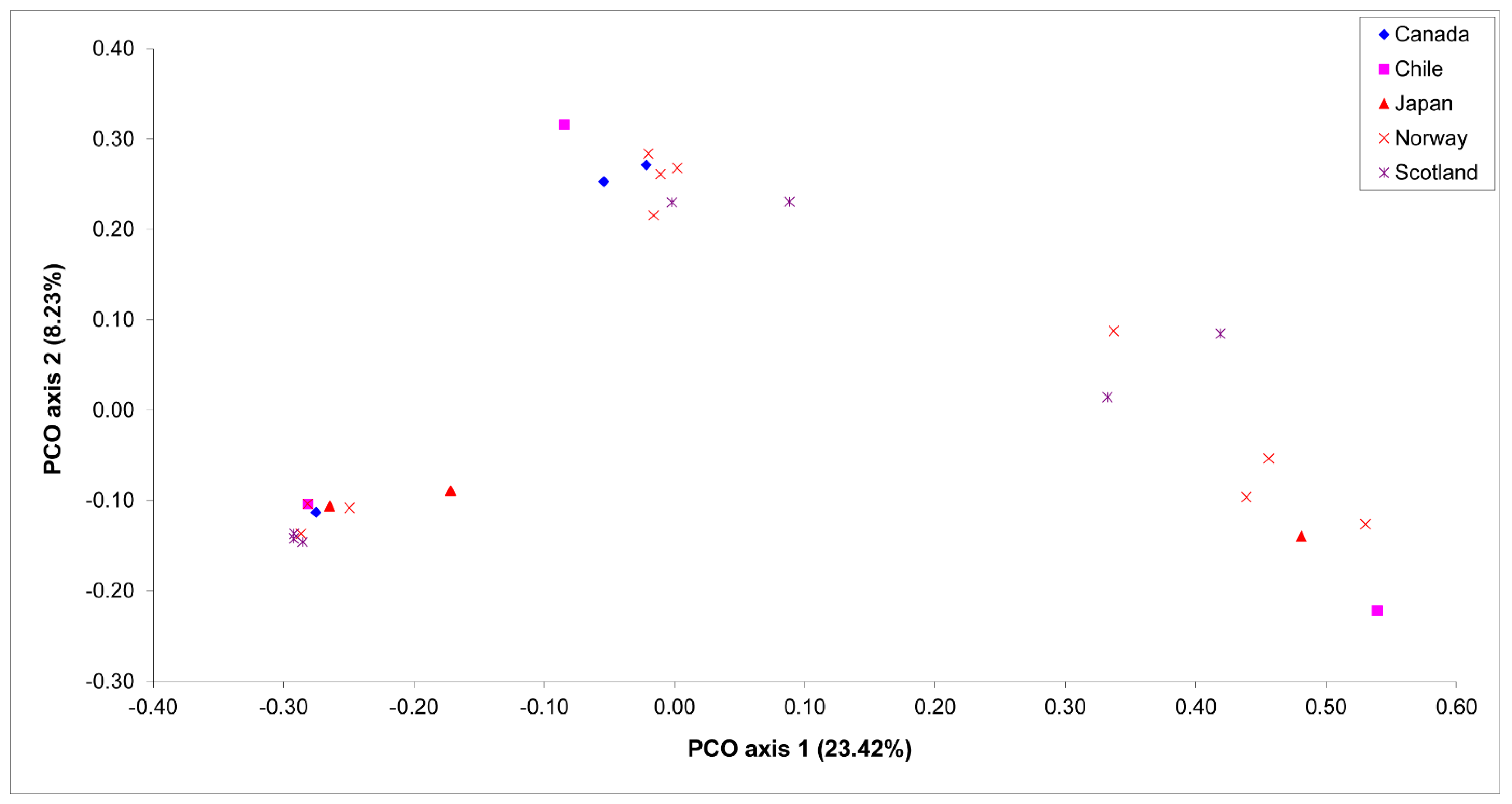
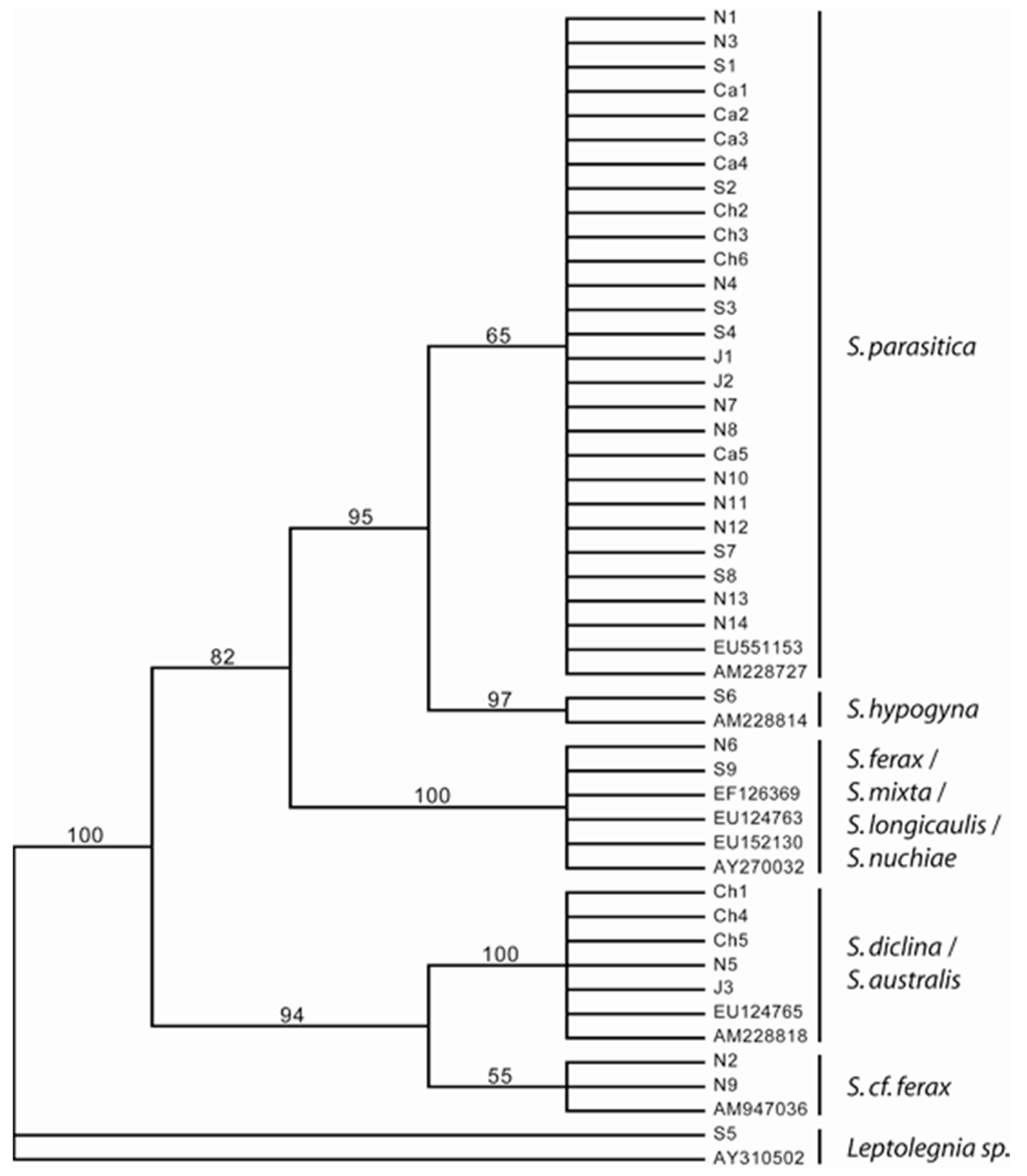
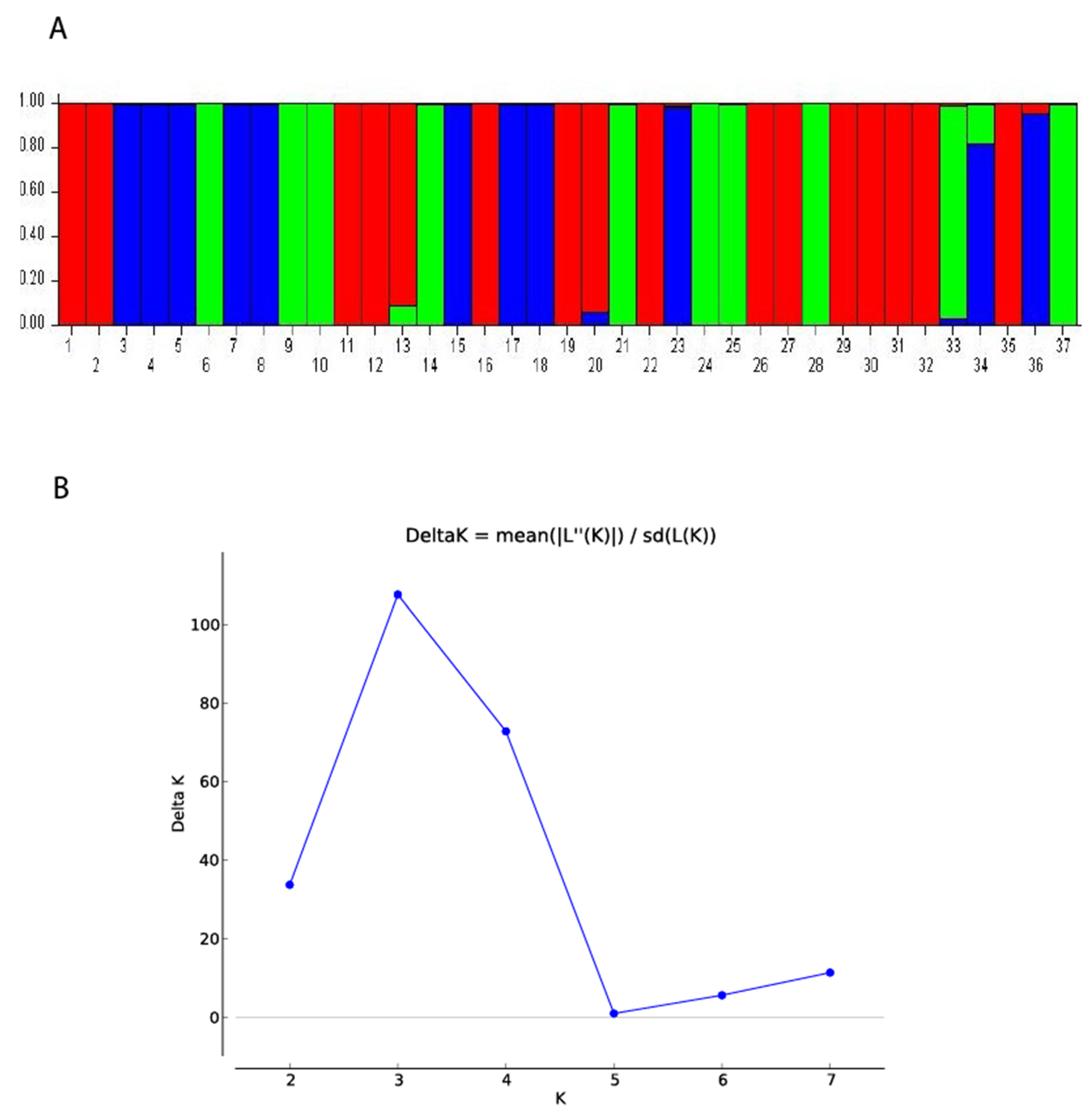
3. Results
4. Discussion
4.1. Geographical Origin
4.2. Morphology
4.3. Phylogeny
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van den Berg, A.H.; McLaggan, D.; Dieguez-Uribeond, J.; van West, P. The impact of the water moulds Saprolegnia diclina and Saprolegnia parasitica on natural ecosystems and the aquaculture industry. Fungal Biol. Rev. 2013, 27, 33–42. [Google Scholar] [CrossRef]
- Torto-Alalibo, T.; Tian, M.Y.; Gajendran, K.; Waugh, M.E.; van West, P.; Kamoun, S. Expressed sequence tags from the oomycete fish pathogen Saprolegnia parasitica reveal putative virulence factors. BMC Microbiol. 2005, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- Dick, M.W. Straminipilous Fungi: Systematics of the Peronosporomycetes Including Accounts of the Marine Straminipilous Protists, the Plasmodiophorids and Similar Organisms; Kluwer Academic, Springer Science & Business Media: Dordrecht, The Netherlands, 2001. [Google Scholar]
- Hughes, G. Saprolegniasis, Then and Now: A Retrospective. Salmon Saprolegniasis; Bonneville Power Administration: Portland, OR, USA, 1994; pp. 3–32. [Google Scholar]
- Mueller, G.J. Salmon Saprolegniasis; Prepared for US Department of Energy; Bonneville Power Administration: Portland, OR, USA, 1994. [Google Scholar]
- Diéguez-Uribeondo, J.; Fregeneda-Grandes, J.M.; Cerenius, L.; Pérez-Iniesta, E.; Aller-Gancedo, J.M.; Tellería, M.T.; Söderhäll, K.; Martín, M.P. Re-evaluation of the enigmatic species complex Saprolegnia diclina–Saprolegnia parasitica based on morphological, physiological and molecular data. Fungal Genet. Biol. 2007, 44, 585–601. [Google Scholar] [CrossRef]
- Yuasa, K.; Hatai, K. Relationship between pathogenicity of Saprolegnia spp. isolates to rainbow trout and their biological characteristics. Fish Pathol. 1995, 30, 101–106. [Google Scholar] [CrossRef]
- Grandes, J.M.F.; Diez, M.F.; Gancedo, J.M.A. Ultrastructural analysis of Saprolegnia secondary zoospore cyst ornamentation from infected wild brown trout, Salmo trutta L. and river water indicates two distinct morphotypes amongst long-spined isolates. J. Fish Dis. 2000, 23, 147–160. [Google Scholar] [CrossRef]
- Hussein, M.M.; Hatai, K. Pathogenicity of Saprolegnia species associated with outbreaks of salmonid saprolegniosis in Japan. Fish. Sci. 2002, 68, 1067–1072. [Google Scholar] [CrossRef]
- Stueland, S.; Hatai, K.; Skaar, I. Morphological and physiological characteristics of Saprolegnia spp. strains pathogenic to Atlantic salmon, Salmo salar L. J. Fish Dis. 2005, 28, 445–453. [Google Scholar] [CrossRef]
- Seymour, R.L. The genus Saprolegnia. Nova Hedwig. 1970, 19, 1–124. [Google Scholar]
- Molina, F.I.; Jong, S.-C.; Ma, G. Molecular characterization and identification of Saprolegnia by restriction analysis of genes coding for ribosomal RNA. Antonie Van Leeuwenhoek 1995, 68, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Bangyeekhun, E.; Pylkkö, P.; Vennerström, P.; Kuronen, H.; Cerenius, L. Prevalence of a single fish-pathogenic Saprolegnia sp. clone in Finland and Sweden. Dis. Aquat. Org. 2003, 53, 47–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulvey, J.P.; Padgett, D.E.; Bailey, J.C. Species boundaries within Saprolegnia (Saprolegniales, Oomycota) based on morphological and DNA sequence data. Mycologia 2007, 99, 421–429. [Google Scholar] [CrossRef]
- Paul, Y.; Leung, W.L.; Hintz, W.E. Species composition of the genus Saprolegnia in fin fish aquaculture environments, as determined by nucleotide sequence analysis of the nuclear rDNA ITS regions. Fungal Biol. 2015, 119, 27–43. [Google Scholar]
- Sandoval-Sierra, J.V.; Martin, M.P.; Dieguez-Uribeondo, J. Species identification in the genus Saprolegnia (Oomycetes): Defining DNA-based molecular operational taxonomic units. Fungal Biol. 2014, 118, 559–578. [Google Scholar] [CrossRef]
- Hatai, K.; Hoshiai, G.-I. Characteristics of two Saprolegnia species isolated from coho salmon with saprolegniosis. J. Aquat. Anim. Health 1993, 5, 115–118. [Google Scholar] [CrossRef]
- Hussein, M.M.; Hatai, K.; Nomura, T. Saprolegniosis in salmonids and their eggs in Japan. J. Wildl. Dis. 2001, 37, 204–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitancharoen, N.; Hatai, K. Experimental infection of Saprolegnia spp. in rainbow trout eggs. Fish Pathol. 1996, 31, 49–50. [Google Scholar] [CrossRef] [Green Version]
- Elameen, A.; Fjellheim, S.; Larsen, A.; Rognli, O.A.; Sundheim, L.; Msolla, S.; Masumba, E.; Mtunda, K.; Klemsdal, S.S. Analysis of genetic diversity in a sweet potato (Ipomoea batatas L.) germplasm collection from Tanzania as revealed by AFLP. Genet. Resour. Crop Evol. 2008, 55, 397–408. [Google Scholar] [CrossRef]
- Dice, L.R. Measures of the amount of ecologic association between species. Ecology 1945, 26, 297–302. [Google Scholar] [CrossRef]
- Jaccard, P. Nouvelles Recherches sur la Distribution Florale; Bull. Soc. Vaud. Sci. Nat. 1908, 44, 223–270. [Google Scholar]
- Sneath, P.H.; Sokal, R.R. Numerical Taxonomy. The Principles and Practice of Numerical Classification; Taylor & Francis, Ltd.: Oxford, UK, 1973. [Google Scholar]
- Rohlf, F.J. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System; Applied Biostatistics: New York, NY, USA, 1992. [Google Scholar]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef]
- Schneider, S.; Roessli, D.; Excoffier, L. Arlequin: A software for population genetics data analysis. User Man. Ver 2000, 2, 2496–2497. [Google Scholar]
- Excoffier, L.; Smouse, P.E. Using allele frequencies and geographic subdivision to reconstruct gene trees within a species: Molecular variance parsimony. Genetics 1994, 136, 343–359. [Google Scholar] [CrossRef]
- Whitlock, M.C.; McCauley, D.E. Indirect measures of gene flow and migration: FST≠ 1/(4Nm+ 1). Heredity 1999, 82, 117–125. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT; Nucleic acids symposium series; ScienceOpen: Burlington, NJ, USA, 1999; pp. 95–98. [Google Scholar]
- Matthews, E.; Ellison, A.; Cable, J. Saprolegnia parasitica zoospore activity and host survival indicates isolate variation in host preference. Fungal Biol. 2021, 125, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. Paup-a computer-program for phylogenetic inference using maximum parsimony. J. Gen. Physiol. 1993, 102, 9–10. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earl, D.A. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Paul, Y.; Naumann, C.; Hintz, W.E. Assessment of intra-specific variability in Saprolegnia parasitica populations of aquaculture facilities in British Columbia, Canada. Dis. Aquat. Org. 2018, 128, 235–248. [Google Scholar]
- Willoughby, L. Rapid preliminary screening of Saprolegnia on fish. J. Fish Dis. 1985, 8, 473–476. [Google Scholar] [CrossRef]
- Beakes, G. Features which characterize Saprolegnia isolates from salmonid fish lesions-a review. Salmon Saprolegniasis 1994, 33–66. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Id | Species | Strain Number * | EMBL Accession Number | Origin | Host | Cyst Coat ** | Germ Rate *** | Path Test **** |
|---|---|---|---|---|---|---|---|---|
| N1 | S. parasitica | VI 02388 | FN186015 | Norway | As parr | + | Slow | - |
| N2 | S. cf. ferax | VI 02389 | FN186016 | Norway | AS parr | − | NT | NT |
| N3 | S. parasitica | VI 02391 | FN186017 | Norway | As parr | + | Fast | ++ |
| N4 | S. parasitica | VI 02750 | FN186018 | Norway | As parr | + | NT | NT |
| N5 | S. diclina | VI 02753 | FN186019 | Norway | As parr | − | NT | NT |
| N6 | S. ferax | VI 02756 | FN186020 | Norway | As parr | − | Slow | − |
| N7 | S. parasitica | VI 02763 | FN186021 | Norway | Bt broodstock | + | NT | NT |
| N8 | S. parasitica | VI 02770 | FN186022 | Norway | As parr | + | Fast | NT |
| N9 | S. cf. ferax | VI 04808 | FN186023 | Norway | As fry | NT | NT | NT |
| N10 | S. parasitica | VI 04810 | FN186024 | Norway | As parr | NT | NT | NT |
| N11 | S. parasitica | VI 04811 | FN186025 | Norway | As parr | NT | NT | NT |
| N12 | S. parasitica | VI 04812 | FN186026 | Norway | As parr | NT | NT | NT |
| N13 | S. parasitica | VI 04817 | FN186027 | Norway | As broodstock | NT | NT | NT |
| N14 | S. parasitica | VI 04819 | FN186028 | Norway | Cwf | NT | NT | NT |
| S1 | S. parasitica | VI 02392 | FN186029 | Scotland | As parr | + | Fast | NT |
| S2 | S. parasitica | VI 02736 | FN186030 | Scotland | As parr | + | Fast | ++ |
| S3 | S. parasitica | VI 02757 | FN186031 | Scotland | As parr | + | Fast | NT |
| S4 | S. parasitica | VI 02758 | FN186032 | Scotland | As parr | + | Fast | NT |
| S5 | Leptolegnia sp. | VI 04813 | FN186033 | Scotland | As parr | NT | NT | NT |
| S6 | S. hypogyna | VI 04814 | FN186034 | Scotland | As parr | NT | NT | NT |
| S7 | S. parasitica | VI 04815 | FN186035 | Scotland | As parr | NT | NT | NT |
| S8 | S. parasitica | VI 04816 | FN186036 | Scotland | As parr | NT | NT | NT |
| S9 | Saprolegnia sp. | VI 04818 | FN186037 | Scotland | As parr | NT | NT | NT |
| Ca1 | S. parasitica | VI 02393 | FN186038 | Canada | As parr | + | Slow | NT |
| Ca2 | S. parasitica | VI 02394 | FN186039 | Canada | As parr | + | Fast | NT |
| Ca3 | S. parasitica | VI 02395 | FN186040 | Canada | As parr | + | Medium | − |
| Ca4 | S. parasitica | VI 02437 | FN186041 | Canada | As parr | + | NT | NT |
| Ca5 | S. parasitica | VI 04809 | FN186042 | Canada | Ps | NT | NT | NT |
| Ch1 | S. diclina | VI 02739 | FN186043 | Chile | As eggs | − | Slow | − |
| Ch2 | S. parasitica | VI 02740 | FN186044 | Chile | As broodstock | + | Medium | − |
| Ch3 | S. parasitica | VI 02741 | FN186045 | Chile | As eggs | + | Medium | NT |
| Ch4 | S. diclina | VI 02744 | FN186046 | Chile | As eggs | NT | Slow | NT |
| Ch5 | S. diclina | VI 02746 | FN186047 | Chile | Cs eggs | NT | NT | NT |
| Ch6 | S. parasitica | VI 02748 | FN186048 | Chile | As eggs | + | Fast | NT |
| J1 | S. salmonis | NJM 9851 | FN186049 | Japan | Ss | + | Fast | ++ |
| J2 | S. parasitica | ATCC90213 | FN186050 | Japan | Cs parr | + | Fast | ++ |
| J3 | S. diclina | ATCC90215 | FN186051 | Japan | Cs parr | − | Slow | − |
| Primer Combination | EcoRI Primer 5′ → 3′ | MseI Primer 5′ → 3′ | Number of Polymorphic Bands |
|---|---|---|---|
| E12 × M17 | GAC-TGC-GTA-CCA-ATT-CAC | GAT-GAG-TCC-TGA-GTA-ACG | 27 |
| E13 × M15 | GAC-TGC-GTA-CCA-ATT-CAG | GAT-GAG-TCC-TGA-GTA-ACA | 28 |
| E11 × M16 | GAC-TGC-GTA-CCA-ATT-CAA | GAT-GAG-TCC-TGA-GTA-ACC | 23 |
| E20 × M17 | GAC-TGC-GTA-CCA-ATT-CGC | GAT-GAG-TCC-TGA-GTA-ACG | 31 |
| E21 × M17 | GAC-TGC-GTA-CCA-ATT-CGG | GAT-GAG-TCC-TGA-GTA-ACG | 30 |
| E22 × M17 | GAC-TGC-GTA-CCA-ATT-CGT | GAT-GAG-TCC-TGA-GTA-ACG | 24 |
| Country of Origin | No. Polymorphic Bands | % Polymorphic Bands |
|---|---|---|
| Canada | 48 | 29.45 |
| Chile | 86 | 52.76 |
| Norway | 137 | 84.05 |
| Scotland | 91 | 55.83 |
| Mean | 90.5 | 55.52 |
| Regions | Canada | Chile | Norway | Scotland |
|---|---|---|---|---|
| Canada | 0.00000 | |||
| Chile | 0.25198 | 0.00000 | ||
| Norway | 0.12585 | 0.08865 | 0.00000 | |
| Scotland | 0.14977 | 0.15440 | 0.04209 | 0.00000 |
| Regions | d.f. | Sum of Squares | Variance Components | % of Variation | FST Value |
|---|---|---|---|---|---|
| Among countries | 3 | 101.763 | 1.86936 | 8.99 | 0.089 |
| Within countries | 30 | 567.884 | 18.92947 | 91.01 | |
| Total | 669.647 | 669.647 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elameen, A.; Stueland, S.; Kristensen, R.; Fristad, R.F.; Vrålstad, T.; Skaar, I. Genetic Analyses of Saprolegnia Strains Isolated from Salmonid Fish of Different Geographic Origin Document the Connection between Pathogenicity and Molecular Diversity. J. Fungi 2021, 7, 713. https://doi.org/10.3390/jof7090713
Elameen A, Stueland S, Kristensen R, Fristad RF, Vrålstad T, Skaar I. Genetic Analyses of Saprolegnia Strains Isolated from Salmonid Fish of Different Geographic Origin Document the Connection between Pathogenicity and Molecular Diversity. Journal of Fungi. 2021; 7(9):713. https://doi.org/10.3390/jof7090713
Chicago/Turabian StyleElameen, Abdelhameed, Svein Stueland, Ralf Kristensen, Rosa F. Fristad, Trude Vrålstad, and Ida Skaar. 2021. "Genetic Analyses of Saprolegnia Strains Isolated from Salmonid Fish of Different Geographic Origin Document the Connection between Pathogenicity and Molecular Diversity" Journal of Fungi 7, no. 9: 713. https://doi.org/10.3390/jof7090713
APA StyleElameen, A., Stueland, S., Kristensen, R., Fristad, R. F., Vrålstad, T., & Skaar, I. (2021). Genetic Analyses of Saprolegnia Strains Isolated from Salmonid Fish of Different Geographic Origin Document the Connection between Pathogenicity and Molecular Diversity. Journal of Fungi, 7(9), 713. https://doi.org/10.3390/jof7090713