
Discovery and Extrolite Production of Three New Species of Talaromyces Belonging to Sections Helici and Purpurei from Freshwater in Korea

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation
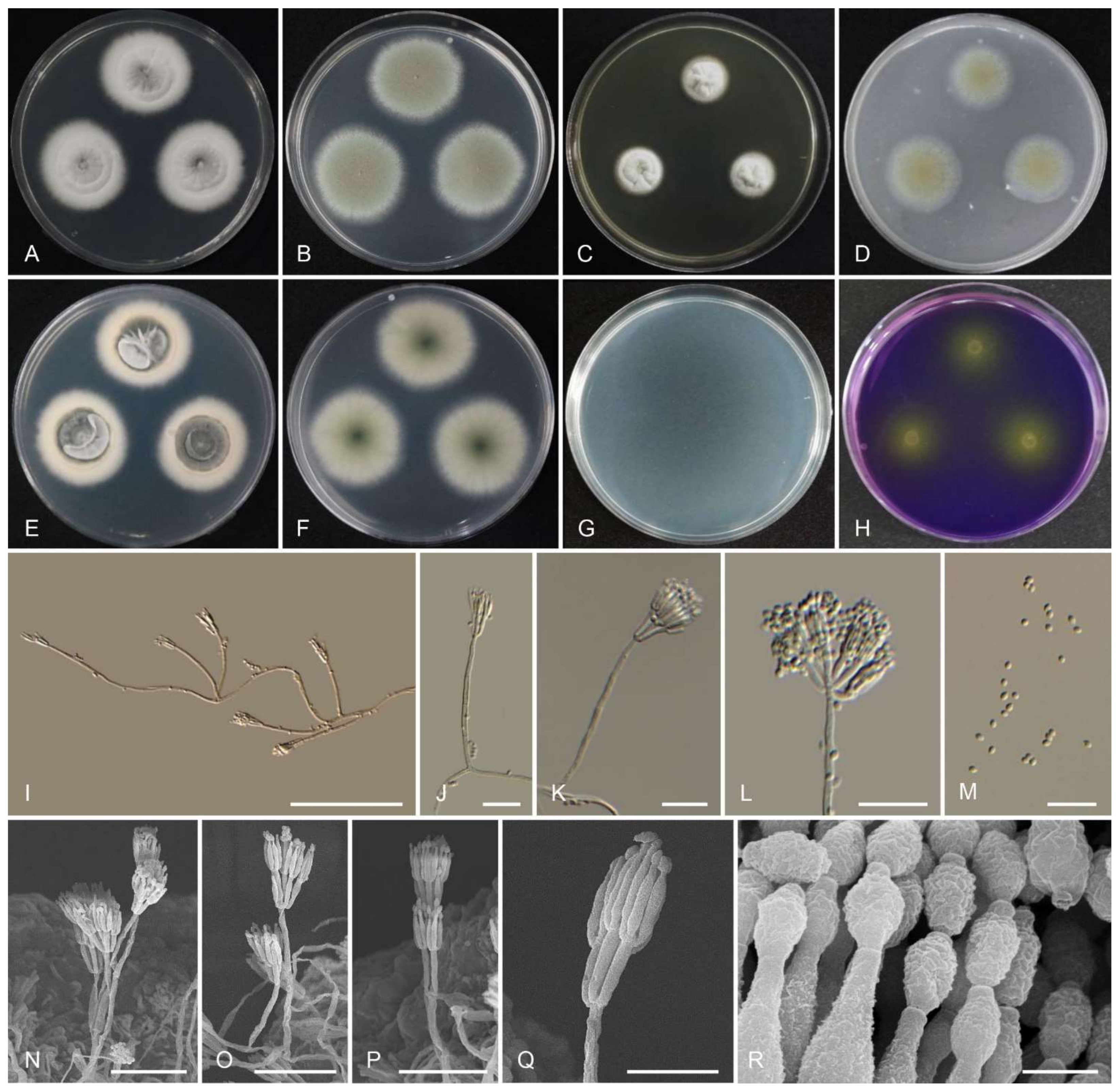
2.2. Morphology
2.3. DNA Extraction, PCR, and Sequencing
2.4. Molecular Analysis
2.5. Extrolite Analysis
3. Results
3.1. Phylogenetic Analysis
3.2. Taxonomy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Benjamin, C.R. Ascocarps of Aspergillus and Penicillium. Mycologia 1995, 47, 669–687. [Google Scholar] [CrossRef]
- Samson, R.A.; Yilmaz, N.; Houbraken, J.; Spierenburg, H.; Seifert, K.A.; Peterson, S.W.; Varga, J.; Frisvad, J.C. Phylogeny and nomenclature of the genus Talaromyces and taxa accommodated in Penicillium subgenus Biverticillium. Stud. Mycol. 2011, 70, 159–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yilmaz, N.; Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of the genus Talaromyces. Stud. Mycol. 2014, 78, 175–341. [Google Scholar] [CrossRef] [Green Version]
- Visagie, C.M.; Yilmaz, N.; Frisvad, J.C.; Houbraken, J.; Seifert, K.A.; Samson, R.A.; Jacobs, K. Five new Talaromyces species with ampulliform-like phialides and globose rough walled conidia resembling T. verruculosus. Mycoscience 2015, 56, 486–502. [Google Scholar] [CrossRef]
- Chen, A.J.; Sun, B.D.; Houbraken, J.; Frisvad, J.C.; Yilmaz, N.; Zhou, Y.G.; Samson, R.A. New Talaromyces species from indoor environments in China. Stud. Mycol. 2016, 84, 119–144. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Lu, X.; Bi, W.; Liu, F.; Gao, W. Talaromyces rubrifaciens, a new species discovered from heating, ventilation and air conditioning systems in China. Mycologia 2016, 108, 773–779. [Google Scholar] [CrossRef]
- Romero, S.M.; Romero, A.I.; Barrera, V.; Comerio, R. Talaromyces systylus, a new synnematous species from Argentinean semiarid soil. Nova Hedwigia 2016, 102, 241–256. [Google Scholar] [CrossRef]
- Wang, X.C.; Chen, K.; Xia, Y.W.; Wang, L.; Li, T.; Zhuang, W.Y. A new species of Talaromyces (Trichocomaceae) from the Xisha Islands, Hainan, China. Phytotaxa 2016, 267, 187–200. [Google Scholar] [CrossRef]
- Yilmaz, N.; López-Quintero, C.A.; Vasco-Palacios, A.M.; Frisvad, J.C.; Theelen, B.; Boekhout, T.; Samson, R.A.; Houbraken, J. Four novel Talaromyces species isolated from leaf litter from Colombian Amazon rain forests. Mycol. Prog. 2016, 15, 1041–1056. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, N.; Visagie, C.M.; Frisvad, J.C.; Houbraken, J.; Jacobs, K.; Samson, R.A. Taxonomic re-evaluation of species in Talaromyces section Islandici, using a polyphasic approach. Persoonia 2016, 36, 37–56. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Carnegie, A.J.; Hardy, G.S.; Smith, D.; Summerell, B.A.; Cano-Lira, J.F.; Guarro, J.; Houbraken, J.; et al. Fungal Planet description sheets 625–715. Persoonia 2017, 39, 460–461. [Google Scholar] [CrossRef] [PubMed]
- Guevara-Suarez, M.; Sutton, D.A.; Gené, J.; García, D.; Wiederhold, N.; Guarro, J.; Cano-Lira, J.F. Four new species of Talaromyces from clinical sources. Mycoses 2017, 60, 651–662. [Google Scholar] [CrossRef]
- Peterson, S.W.; Jurjević, Ž. New species of Talaromyces isolated from maize, indoor air, and other substrates. Mycologia 2017, 109, 537–556. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.C.; Chen, K.; Qin, W.T.; Zhuang, W.Y. Talaromyces heiheensis and T.mangshanicus, two new species from China. Mycol. Prog. 2017, 16, 73–81. [Google Scholar]
- Barbosa, R.N.; Bezerra, J.D.; Souza-Motta, C.M.; Frisvad, J.C.; Samson, R.A.; Oliveira, N.T.; Houbraken, J. New Penicillium and Talaromyces species from honey, pollen and nests of stingless bees. Antonie van Leeuwenhoek 2018, 13, 1883–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.S.; Gené, J.; Guarro, J.; Baseia, I.G.; García, D.; Gusmão, L.F.; Souza-Motta, C.M.; et al. Fungal Planet description sheets: 716–784. Persoonia 2018, 40, 239–392. [Google Scholar] [CrossRef]
- Crous, P.W.; Luangsa-Ard, J.J.; Wingfield, M.J.; Carnegie, A.J.; Hernández-Restrepo, M.; Lombard, L.; Roux, J.; Barreto, R.W.; Baseia, I.G.; Cano-Lira, J.F.; et al. Fungal Planet description sheets: 785–867. Persoonia 2018, 41, 238–417. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.Z.; Yu, Z.D.; Ruan, Y.M.; Wang, L. Three new species of Talaromyces sect. Talaromyces discovered from soil in China. Sci. Rep. 2018, 8, 4932. [Google Scholar]
- Su, L.; Niu, Y.C. Multilocus phylogenetic analysis of Talaromyces species isolated from curcurbit plants in China and description of two new species, T. curcurbitiradicus and T. endophyticus. Mycologia 2018, 110, 375–386. [Google Scholar] [CrossRef]
- Varriale, S.; Houbraken, J.; Granchi, Z.; Pepe, O.; Cerullo, G.; Ventorino, V.; Chin-A-Woeng, T.; Meijer, M.; Riley, R.; Grigoriev, I.V.; et al. Talaromyces borbonicus sp. nov., a novel fungus from biodegraded Arundo donax with potential abilities in lignocellulose conversion. Mycologia 2018, 110, 316–324. [Google Scholar] [CrossRef] [Green Version]
- Rajeshkumar, K.C.; Yilmaz, N.; Marathe, S.D.; Seifert, K.A. Morphology and multigene phylogeny of Talaromyces amyrossmaniae, a new synnematous species belonging to the section Trachyspermi from India. Mycokeys 2019, 45, 41–56. [Google Scholar] [CrossRef]
- Peterson, S.W.; Jurjevic, Z. The Talaromyces pinophilus species complex. Fungal Biol. 2019, 123, 745–762. [Google Scholar] [CrossRef]
- Doilom, M.; Guo, J.W.; Phookamsak, R.; Mortimer, P.E.; Karunarathna, S.C.; Dong, W.; Liao, C.F.; Yan, K.; Pem, D.; Suwannarach, N.; et al. Screening of phosphate-solubilizing fungi from air and soil in Yunnan, China: Four novel species in Aspergillus, Gongronella, Penicillium, and Talaromyces. Front. Microbiol. 2020, 11, 585215. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Andrade, E.; Stchigel, A.M.; Terrab, A.; Guarro, J.; Cano-Lira, J.F. Diversity of xerotolerant and xerophilic fungi in honey. IMA Fungus 2019, 10, 20. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Cowan, D.A.; Maggs-Kölling, G.; Yilmaz, N.; Larsson, E.; Angelini, C.; Brandrud, T.E.; Dearnaley, J.D.; Dima, B.; Dovana, F.; et al. Fungal Planet description sheets: 1112–1181. Persoonia 2020, 45, 251–409. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.D.; Chen, A.J.; Houbraken, J.; Frisvad, J.C.; Wu, W.P.; Wei, H.L.; Zhou, Y.G.; Jiang, X.Z.; Samson, R.A. New section and species in Talaromyces. Mycokeys 2020, 68, 75–113. [Google Scholar] [CrossRef]
- Houbraken, J.; Kocsubé, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Sang, H.; An, T.; Kim, C.S.; Shin, G.; Sung, G.; Yu, S.H. Two novel Talaromyces species isolated from medicinal crops in Korea. J. Microbiol. 2013, 51, 704–708. [Google Scholar] [CrossRef]
- You, Y.H.; Aktaruzzaman, M.; Heo, I.; Park, J.M.; Hong, J.W.; Hong, S.B. Talaromyces halophytorum sp. nov. isolated from roots of Limonium tetragonum in Korea. Mycobiology 2020, 48, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Reyes, I.; Bernier, L.; Simard, R.R. Characteristics of phosphate solubilization by an isolate of a tropical Penicillium rugulosum and two UV-induced mutants. FEMS Microbiol. Ecol. 1999, 28, 291–295. [Google Scholar] [CrossRef]
- Narikawa, T.; Shinoyama, H.; Fujii, T. A β-rutinosidase from Penicillum rugulosum IFO 7242 that is a peculiar flavonoid glycosidase. Biosci. Biotechnol. Biochem. 2000, 64, 1317–1319. [Google Scholar] [CrossRef]
- Pol, D.; Laxman, R.S.; Rao, M. Purification and biochemical characterization of endoglucanase from Penicillium pinophilum MS 20. Indian J. Biochem. Biophys. 2012, 49, 189–194. [Google Scholar] [PubMed]
- Maeda, R.N.; Barcelos, C.A.; Anna, L.M.M.S. Cellulase production by Penicillium funiculosum and its application in the hydrolysis of sugar cane bagasse for second generation ethanol production by fed batch operation. J. Biotechnol. 2013, 163, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, N.; Houbraken, J.; Hoekstra, E.S.; Frisvad, J.C.; Visagie, C.M.; Samson, R.A. Delimitation and characterisation of Talaromyces purpurogenus and related species. Persoonia 2012, 29, 39–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frisvad, J.C.; Yilmaz, N.; Thrane, U.; Rasmussen, K.B.; Houbraken, J.; Samson, R.A. Talaromyces atroroseus, a new species efficiently producing industrially relevant red pigments. PLoS ONE 2013, 8, e84102. [Google Scholar] [CrossRef] [Green Version]
- Kakvan, N.; Heydari, A.; Zamanizadeh, H.R.; Rezaee, S.; Naraghi, L. Development of new bioformulations using Trichoderma and Talaromyces fungal antagonists for biological control of sugar beet damping-off disease. Crop Prot 2013, 53, 80–84. [Google Scholar] [CrossRef]
- Marois, J.J.; Fravel, D.R.; Papavizas, G.C. Ability of Talaromyces flavus to occupy the rhizosphere. Soil Biol. Biochem. 1984, 16, 387–390. [Google Scholar] [CrossRef]
- Fravel, D.R.; Davis, J.R.; Sorenson, L.H. Effect of Talaromyces flavus and metham on verticillium wilt incidence and potato yield 1984–1985. Biol. Cult. Tests 1986, 1, 17. [Google Scholar]
- McLaren, D.L.; Huang, H.C.; Kozub, G.C.; Rimmer, S.R. Biological control of sclerotinia wilt of sunflower with Talaromyces flavus and Coniothyrium minitans. Plant Dis. 1994, 78, 231–235. [Google Scholar] [CrossRef]
- Naraghi, L.; Heydari, A.; Rezaee, S.; Razavi, M.; Jahanifar, H. Study on antagonistic effects of Talaromyces flavus on Verticillium albo-atrum, the causal agent of potato wilt disease. Crop Prot. 2010, 29, 658–662. [Google Scholar] [CrossRef]
- Pretsch, A.; Nag, M.; Schwendinger, K.; Kreiseder, B.; Wiederstein, M.; Pretsch, D.; Genov, M.; Hollaus, R.; Zinssmeister, D.; Debbab, A.; et al. Antimicrobial and anti-inflammatory activities of endophytic fungi Talaromyces wortmannii extracts against acne-inducing bacteria. PLoS ONE 2014, 9, e97929. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.L.; Ribas, J.L.; Gibson, D.W.; Connor, D.H. Infections caused by Penicillium marneffei in China and Southeast Asia. Review of eighteen cases and report of four more Chinese cases. Rev. Infec. Dis. 1988, 10, 640–652. [Google Scholar] [CrossRef]
- Hien, T.V.; Loc, P.P.; Hoa, N.T.T. First case of disseminated Penicilliosis marneffei infection among patients with acquired immunodeficiency syndrome in Vietnam. Clin. Infect. Dis. 2001, 32, 78–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolk, A.C.; Samson, R.A. Studies on Talaromyces and related genera II. The genus Talaromyces. Stud. Mycol. 1972, 2, 1221. [Google Scholar]
- Goh, T.K.; Hyde, K.D. Biodiversity of freshwater fungi. J. Ind. Microbiol. 1996, 17, 328–345. [Google Scholar] [CrossRef]
- Hawksworth, D.L. The fungal dimension of biodiversity: Magnitude, significance, and conservation. Mycol. Res. 1991, 95, 641–655. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Lücking, R. Fungal diversity revisited 2.2 to 3.8 million species. Microbiol Spectr. 2017, 5, FUNK-0052-2016. [Google Scholar] [CrossRef]
- Shearer, C.A.; Descals, E.; Kohlmeyer, B.; Kohlmeyer, J.; Marvanová, L.; Padgett, D.; Porter, D.; Raja, H.A.; Schmit, J.P.; Thorton, H.A.; et al. Fungal biodiversity in aquatic habitats. Biodivers. Conserv. 2017, 16, 49–67. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Hyde, K.D.; Pang, K.L. Freshwater Fungi and Fungal-Like Organisms; De Gruyter: Boston, MA, USA, 2014. [Google Scholar]
- Nguyen, T.T.T.; Paul, N.C.; Lee, H.B. Characterization of Paecilomyces variotii and Talaromyces amestolkiae in Korea based on the morphological characteristics and multigene phylogenetic analyses. Mycobiology 2016, 44, 248–259. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Cambridge, MA, USA, 1990; pp. 315–322. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 6, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Peterson, S.W.; Vega, F.; Posada, F.; Nagai, C. Penicillium coffeae, a new endophytic species isolated from a coffee plant and its phylogenetic relationship to P. fellutanum, P. thiersii and P. brocae based on parsimony analysis of multilocus DNA sequences. Mycologia 2005, 97, 659–666. [Google Scholar] [CrossRef]
- Hong, S.B.; Cho, H.S.; Shin, H.D.; Frisvad, J.C.; Samson, R.A. Novel Neosartorya species isolated from soil in Korea. Int. J. Syst. Evol. Microbiol. 2006, 56, 477–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Houbraken, J.; Samson, R.A. Phylogeny of Penicillium and the segregation of Trichocomaceae into three families. Stud. Mycol. 2011, 70, 1–51. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. TrimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glez-Peña, D.; Gómez-Blanco, D.; Reboiro-Jato, M.; Fdez-Riverola, F.; Posada, D. ALTER: Program–oriented format conversion of DNA and protein alignments. Nucleic Acids. Res. 2010, 38, 14–18. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree, Version 1.3. 1. Computer Program Distributed by the Author. 2009. Available online: http://www.treebioedacuk/software/fgtree (accessed on 4 January 2011).
- Frisvad, J.C.; Thrane, U. Standardized high performance liquid chromatography of 182 mycotoxins and other fungal metabolites based on alkylphenone indices and UV VIS spectra (diode array detection). J. Chromatogr. 1987, 404, 195–214. [Google Scholar] [CrossRef]
- Nielsen, K.F.; Månsson, M.; Rank, C.; Frisvad, J.C.; Larsen, T.O. Dereplication of microbial natural products by LC-DAD-TOFMS. J. Nat. Prod. 2011, 74, 2338–2348. [Google Scholar] [CrossRef]
- Houbraken, J.; Wang, L.; Lee, H.B.; Frisvad, J.C. New sections in Penicillium containing novel species producing patulin, pyripyropens or other bioactive compounds. Persoonia 2016, 36, 299–314. [Google Scholar] [CrossRef] [Green Version]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Bolchacova, E.; Voigt, K.; Crous, P.W.; et al. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, E.; Fujimoto, H.; Baba, M.; Yamazaki, M. 4 new chlorinated azaphilones, helicusins A-D, closely related to 7-epi-sclerotiorin, from an ascomycetous fungus, Talaromcyes helicus. Chem. Pharm. Bull. 1995, 43, 1307–1310. [Google Scholar] [CrossRef] [Green Version]
- Seifert, K.A.; Hoekstra, E.S.; Frisvad, J.C.; Louis-Seize, G. Penicillium cecidicola, a new species on cynipid insect galls on Quercus pacifica in the western United States. Stud. Mycol. 2004, 50, 517–523. [Google Scholar]
- Visagie, C.M.; Roets, F.; Jacobs, K. A new species of Penicillium, P. ramulosum sp. nov., from the natural environment. Mycologia 2009, 101, 888–895. [Google Scholar] [CrossRef]
- Van der Walt, L.; Spotts, R.A.; Visagie, C.M.; Jacobs, K.; Smit, F.J.; McLeod, A. Penicillium species associated with preharvest wet core rot in South Africa and their pathogenicity on apple. Plant Dis. 2010, 94, 666–675. [Google Scholar] [CrossRef] [Green Version]
- Visagie, C.M.; Jacobs, K. Three new additions to the genus Talaromyces isolated from Atlantis sandveld fynbos soils. Persoonia 2012, 28, 14–24. [Google Scholar] [CrossRef]
- Heo, I.; Hong, K.; Yang, H.; Lee, H.B.; Choi, Y.-J.; Hong, S.-B. Diversity of Aspergillus, Penicillium, and Talaromyces species isolated from freshwater environments in Korea. Mycobiology 2019, 47, 12–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pangging, M.; Nguyen, T.T.T.; Lee, H.B. New records of four species belonging to Eurotiales from soil and freshwater in Korea. Mycobiology 2019, 47, 154–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visagie, C.M.; Houbraken, J. Updating the taxonomy of Aspergillus in South Africa. Stud. Mycol. 2020, 95, 253–292. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon Name | Strain No. | GenBank Accession No. | References | |||
|---|---|---|---|---|---|---|
| ITS | BenA | CaM | RPB2 | |||
| T. aerugineus | CBS 350.66 T | AY753346 | KJ865736 | KJ885285 | JN121502 | [3] |
| T. apiculatus | CBS 312.59 T | JN899375 | KF741916 | KF741950 | KM023287 | [3] |
| T. atricola | CBS 255.31 T | KF984859 | KF984566 | KF984719 | KF984948 | [3] |
| T. atroroseus | CBS 133442 T | KF114747 | KF114789 | KJ775418 | KM023288 | [3] |
| T. austrocalifornicus | CBS 644.95 T | JN899357 | KJ865732 | KJ885261 | MN969147 | [3,27] |
| T. bacillisporus | CBS 296.48 T | KM066182 | AY753368 | KJ885262 | JF417425 | [3] |
| T. bohemicus | CBS 545.86 T | JN899400 | KJ865719 | KJ885286 | JN121532 | [3] |
| T. boninensis | CBS 650.95 T | JN899356 | KJ865721 | KJ885263 | KM023276 | [3] |
| T. borbonicus | CBS 141340 T | MG827091 | MG855687 | MG855688 | MG855689 | [20] |
| T. brunneosporus | FMR 16566 T | LT962487 | LT962483 | LT962488 | LT962485 | [24] |
| T. cecidicola | CBS 101419 T | AY787844 | FJ753295 | KJ885287 | KM023309 | [3] |
| T. cinnabarinus | CBS 267.72 T | JN899376 | AY753377 | KJ885256 | JN121477 | [3] |
| T. cinnabarinus | CBS 357.72 | – | KM066134 | – | – | [3] |
| T. chlamydosporus | CBS 140635 T | KU866648 | KU866836 | KU866732 | KU866992 | [5] |
| T. chlorolomus | DAOM 241016 T | FJ160273 | GU385736 | KJ885265 | KM023304 | [3,27] |
| T. chlorolomus | DTO 180-F4 | – | FJ753294 | – | – | [3] |
| T. chlorolomus | DTO 182-A5 | – | JX091597 | – | – | [3] |
| T. cnidii | KACC 46617 T | KF183639 | KF183641 | KJ885266 | KM023299 | [3,28] |
| T. cinnabarinus | CBS 267.72 T | JN899376 | AY753377 | KJ885256 | JN121477 | [3] |
| T. cinnabarinus | CBS 357.72 | – | KM066134 | – | – | [3] |
| T. coalescens | CBS 103.83 T | JN899366 | JX091390 | KJ885267 | KM023277 | [3] |
| T. columbinus | NRRL 58811 T | KJ865739 | KF196843 | KJ885288 | KM023270 | [3] |
| T. dendriticus | CBS 660.80 T | JN899339 | JX091391 | KF741965 | KM023286 | [3] |
| T. dendriticus | DAOM 226674 | – | FJ753293 | – | – | [3] |
| T. dendriticus | DAOM 233861 | – | FJ753294 | – | – | [3] |
| T. derxii | CBS 412.89 T | JN899327 | JX494306 | KF741959 | KM023282 | [3,27] |
| T. diversiformis | CBS 141931 T | KX961215 | KX961216 | KX961259 | KX961274 | [11] |
| T. diversus | CBS 320.48 T | KJ865740 | KJ865723 | KJ885268 | KM023285 | [3] |
| T. duclauxii | CBS 322.48 T | JN899342 | JX091384 | KF741955 | JN121491 | [3] |
| T. emodensis | CBS 100536 T | JN899337 | KJ865724 | KJ885269 | JF417445 | [27] |
| T. erythromellis | CBS 644.80 T | JN899383 | HQ156945 | KJ885270 | KM023290 | [3] |
| T. euchlorocarpius | DTO 176-I3 T | AB176617 | KJ865733 | KJ885271 | KM023303 | [3] |
| T. flavus | CBS 310.38 T | JN899360 | JX494302 | KF741949 | JF417426 | [3] |
| T. fusiformis | CBS 140637 T | KU866656 | KU866843 | KU866740 | KU867000 | [5] |
| T. georgiensis | DI16-145 T | LT558967 | LT559084 | – | LT795606 | [12] |
| T. gwangjuensis | CNUFC WT19-1 T | MK766233 | MZ318448 | – | MK912174 | This study |
| T. gwangjuensis | CNUFC WT19-2 | MK766234 | MZ318449 | – | MK912175 | This study |
| T. helicus | CBS 335.48 T | JN899359 | KJ865725 | KJ885289 | KM023273 | [3] |
| T. helicus | CBS 134.67 | – | KM066133 | – | – | [3] |
| T. iowaense | NRRL 66822 T | MH281565 | MH282578 | MH282579 | MH282577 | [17] |
| T. islandicus | CBS 338.48 T | KF984885 | KF984655 | KF984780 | KF985018 | [3] |
| T. korena | CNUFC YJW2-13 T | MZ315100 | MZ318450 | MZ332529 | MZ332533 | This study |
| T. korena | CNUFC YJW2-14 | MZ315101 | MZ318451 | MZ332530 | MZ332534 | This study |
| T. mimosinus | CBS 659.80 T | JN899338 | KJ865726 | KJ885272 | MN969149 | [3,27] |
| T. minioluteus | CBS 642.68 T | JN899346 | MN969409 | KJ885273 | JF417443 | [3] |
| T. palmae | CBS 442.88 T | JN899396 | HQ156947 | KJ885291 | KM023300 | [3] |
| T. piceus | CBS 361.48 T | KF984792 | KF984668 | KF984680 | KF984899 | [3] |
| T. pigmentosus | CBS 142805 T | MF278330 | LT855562 | LT855565 | LT855568 | [15] |
| T. pittii | CBS 139.84 T | JN899325 | KJ865728 | KJ885275 | KM023297 | [3] |
| T. proteolyticus | CBS 303.67 T | JN899387 | KJ865729 | KJ885276 | KM023301 | [3] |
| T. pseudostromaticus | CBS 470.70 T | JN899371 | HQ156950 | KJ885277 | KM023298 | [3] |
| T. ptychoconidius | DAOM 241017 T | FJ160266 | GU385733 | JX140701 | KM023278 | [3,27] |
| T. ptychoconidius | DTO 180-E9 | – | GU385734 | – | – | [3] |
| T. ptychoconidius | DTO 180-F1 | – | GU385735 | – | – | [3] |
| T. purpureogenus | CBS 286.36 T | JN899372 | JX315639 | KF741947 | JX315709 | [3,27] |
| T. purpureus | CBS 475.71 T | JN899328 | GU385739 | KJ885292 | JN121522 | [3] |
| T. rademirici | CBS 140.84 T | JN899386 | KJ865734 | KM023302 | [3] | |
| T. radicus | CBS 100489 T | KF984878 | KF984599 | KF984773 | KF985013 | [3] |
| T. ramulosus | DAOM 241660 T | EU795706 | FJ753290 | JX140711 | KM023281 | [3] |
| T. ramulosus | DTO 182-A6 | – | JX091631 | – | – | [3] |
| T. ramulosus | DTO 181-E3 | – | JX091626 | – | – | [3] |
| T. ramulosus | DTO 182-A3 | – | JX091630 | – | – | [3] |
| T. reverso-olivaceus | CBS 140672 T | KU866646 | KU866834 | KU866730 | KU866990 | [5] |
| T. rotundus | CBS 369.48 T | JN899353 | KJ865730 | KJ885278 | KM023275 | [3] |
| T. rugulosus | CBS 371.48 T | KF984834 | KF984575 | KF984702 | KF984925 | [3] |
| T. ryukyuensis | NHL 2917 T | AB176628 | – | – | – | [3] |
| T. stipitatus | CBS 375.48 T | JN899348 | KM111288 | KF741957 | KM023280 | [3] |
| T. subinflatus | CBS 652.95 T | JN899397 | MK450890 | KJ885280 | KM023308 | [3,27] |
| T. tabacinus | NRRL 66727 T | MG182613 | MG182627 | MG182606 | MG182620 | [17] |
| T. tardifaciens | CBS 250.94 T | JN899361 | KF984560 | KF984682 | KF984908 | [27] |
| T. teleomorpha | CNUFC YJW2-5 T | MZ315102 | MZ318452 | MZ332531 | MZ332535 | This study |
| T. teleomorpha | CNUFC YJW2-6 | MZ315103 | MZ318453 | MZ332532 | MZ332536 | This study |
| T. tenuis | CBS 141840 T | MN864275 | MN863344 | MN863321 | MN863333 | [26] |
| T. trachyspermus | CBS 373.48 T | JN899354 | KF114803 | KJ885281 | JF417432 | [3] |
| T. tratensis | CBS 133146 T | KF984891 | KF984559 | KF984690 | KF984911 | [3] |
| T. ucrainicus | CBS 162.67 T | JN899394 | KF114771 | KJ885282 | KM023289 | [3] |
| T. unicus | CBS 100535 T | JN899336 | KJ865735 | KJ885283 | MN969150 | [27] |
| T. varians | CBS 386.48 T | JN899368 | KJ865731 | KJ885284 | KM023274 | [3] |
| T. verruculosus | NRRL 1050 T | KF741994 | KF741928 | KF741944 | KM023306 | [27] |
| T. viridulus | CBS 252.87 T | JN899314 | JX091385 | KF741943 | JF417422 | [3] |
| Trichocoma paradoxa | CBS 788.83 T | JN899398 | KF984556 | KF984670 | JN121550 | [3] |
| Characteristics | CNUFC WT19-1 Isolated in This Study | Talaromyces rademiricia |
|---|---|---|
| Size after 7 days at 25 °C (diameter) | <1 mm on CYA | 5–6 mm on CYA |
| 3–5 mm on YES | 5–6 mm on YES | |
| 13–15 mm on MEA | 14–16 mm on MEA | |
| 6–7 mm on OA | 9–10 mm on OA | |
| No growth on CREA | No growth on CREA | |
| Size after 7 days at 37 °C on CYA (diameter) | No growth | 3 mm |
| Conidiophores | Biverticillate and monoverticillate, 39–174 × 1.5–3 µm | Biverticillate and monoverticillate; stipes smooth-walled, 25–95 × 1.5–2.5 μm; branches 10–15 μm |
| Metulae | Two to six, 6–10 × 1.5–2.5 µm | Two to five, divergent, 7–11 × 2–2.5 μm |
| Phialides | Acerose, three to eight per metula, 5.5–10 × 1.5–2 µm | Acerose, two to six per metula, 7.5–11.5 × 1.5–3 μm |
| Conidia | Globose, 1.5–2.0 µm, smooth-walled | Ellipsoidal, 2.5–4 × 1.5–2.5 μm, smooth |
| Ascomata | Absent | Absent |
| Characteristics | CNUFC YJW2-13 Isolated in This Study | Talaromyces boninensis a | Talaromyces reverso-olivaceus b |
|---|---|---|---|
| Size after 7 days at 25 °C (diameter) | 25–28 mm on CYA | 28 mm on CYA | 19–23 mm on CYA |
| 21–24 mm on YES | NI | 25–26 mm on YES | |
| 41–45 mm on MEA | 30 mm on MEA | 34–37 mm on MEA | |
| 36–39 mm on OA | 32 mm on OA | 33–36 mm on OA | |
| 15–18 mm CREA | NI | No growth on CREA | |
| Size after 7 days at 37 °C | 17–19 mm on CYA | NI | 18–20 mm on CYA |
| Conidiophores | Biverticillate, sometimes with additional branches, stipes smooth, 15–194 × 2–4 μm, branches 6–17 × 2–3 μm | Biverticillate; stipes finely rough, 25–260 × 2.5–4 μm | Biverticillate, sometimes with extra subterminal branches; stipes smooth, 50–100 × 2.5–4 μm, branches 12–15 × 2–3 μm |
| Metulae | Two to seven, 7.5–16 × 2–3 μm | Four to ten, 10–16(–20) × 2.5–3(–3.5) μm | Three to five, 10–13 × 3–4 μm |
| Phialides | Acerose, two to seven per metula, 5.5–15 × 2–3 μm | Acerose, two to six per metula, 10–15 × 2–3.5 μm | Acerose, three to five per metula, 10–12(–14) × 2.5–3 μm |
| Conidia | Ellipsoidal to fusiform, finely roughed, 2–3.5 × 1.5–2.5 μm | Ellipsoidal to fusiform, sometimes globose, smooth, 2–4 × 1.5–2.5 μm | Ellipsoidal to fusiform, finely roughed, 2.5–4.5 × 2.5–3 μm |
| Ascomata | Absent | Grayish green, globose to subglobose, 280–550 × 240–480 μm | Absent |
| Characteristics | CNUFC YJW2-5 Isolated in This Study | Talaromyces helicusa |
|---|---|---|
| Size after 7 days at 25 °C (diameter) | 26–29 mm on CYA | 13–23 mm on CYA |
| 29–33 mm on YES | 14–22 mm on YES | |
| 45–48 mm on MEA | 25–33 mm on MEA | |
| 32–34 mm on OA | 23–35 mm on OA | |
| 1–3 on CREA | No growth on CREA | |
| Size after 7 days at 37 °C (diameter) | 15–20 mm on CYA | 10–18 mm on CYA |
| Conidiophores | Not observed | Mono- to biverticillate, stipes smooth walled, 30–60(–80) × 2–2.5 μm |
| Metulae | Not observed | Two to five, 12–15 × 2–2.5 μm |
| Phialides | Not observed | Acerose, two to four per metula, 8.5–12(–16) × 2.5–3 μm |
| Conidia | Not observed | Globose to subglobose, smooth, 2.5–3.5(–4.5) × 2.2–3.5 μm |
| Ascomata | Creamish-white to yellow to reddish, globose to subglobose, 200–800 μm | Yellow, pastel yellow and creamish-white, globose to subglobose, 100–300 μm |
| Asci | Ellipsoidal, globose to subglobose, (5.5–)6.5–9 × (4.5–)6–7 μm | 6–9 × 4.5–6 μm |
| Ascospores | Ellipsoidal, smooth, 3–4 × 2–3 μm | Ellipsoidal, smooth (some with minute spines), 2.5–4 × 2–3 μm |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, T.T.T.; Frisvad, J.C.; Kirk, P.M.; Lim, H.J.; Lee, H.B. Discovery and Extrolite Production of Three New Species of Talaromyces Belonging to Sections Helici and Purpurei from Freshwater in Korea. J. Fungi 2021, 7, 722. https://doi.org/10.3390/jof7090722
Nguyen TTT, Frisvad JC, Kirk PM, Lim HJ, Lee HB. Discovery and Extrolite Production of Three New Species of Talaromyces Belonging to Sections Helici and Purpurei from Freshwater in Korea. Journal of Fungi. 2021; 7(9):722. https://doi.org/10.3390/jof7090722
Chicago/Turabian StyleNguyen, Thuong T. T., Jens Christian Frisvad, Paul M. Kirk, Hyo Jin Lim, and Hyang Burm Lee. 2021. "Discovery and Extrolite Production of Three New Species of Talaromyces Belonging to Sections Helici and Purpurei from Freshwater in Korea" Journal of Fungi 7, no. 9: 722. https://doi.org/10.3390/jof7090722
APA StyleNguyen, T. T. T., Frisvad, J. C., Kirk, P. M., Lim, H. J., & Lee, H. B. (2021). Discovery and Extrolite Production of Three New Species of Talaromyces Belonging to Sections Helici and Purpurei from Freshwater in Korea. Journal of Fungi, 7(9), 722. https://doi.org/10.3390/jof7090722