
Geographical Distribution and Genetic Diversity of the Banana Fusarium Wilt Fungus in Laos and Vietnam

 , ,
, ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
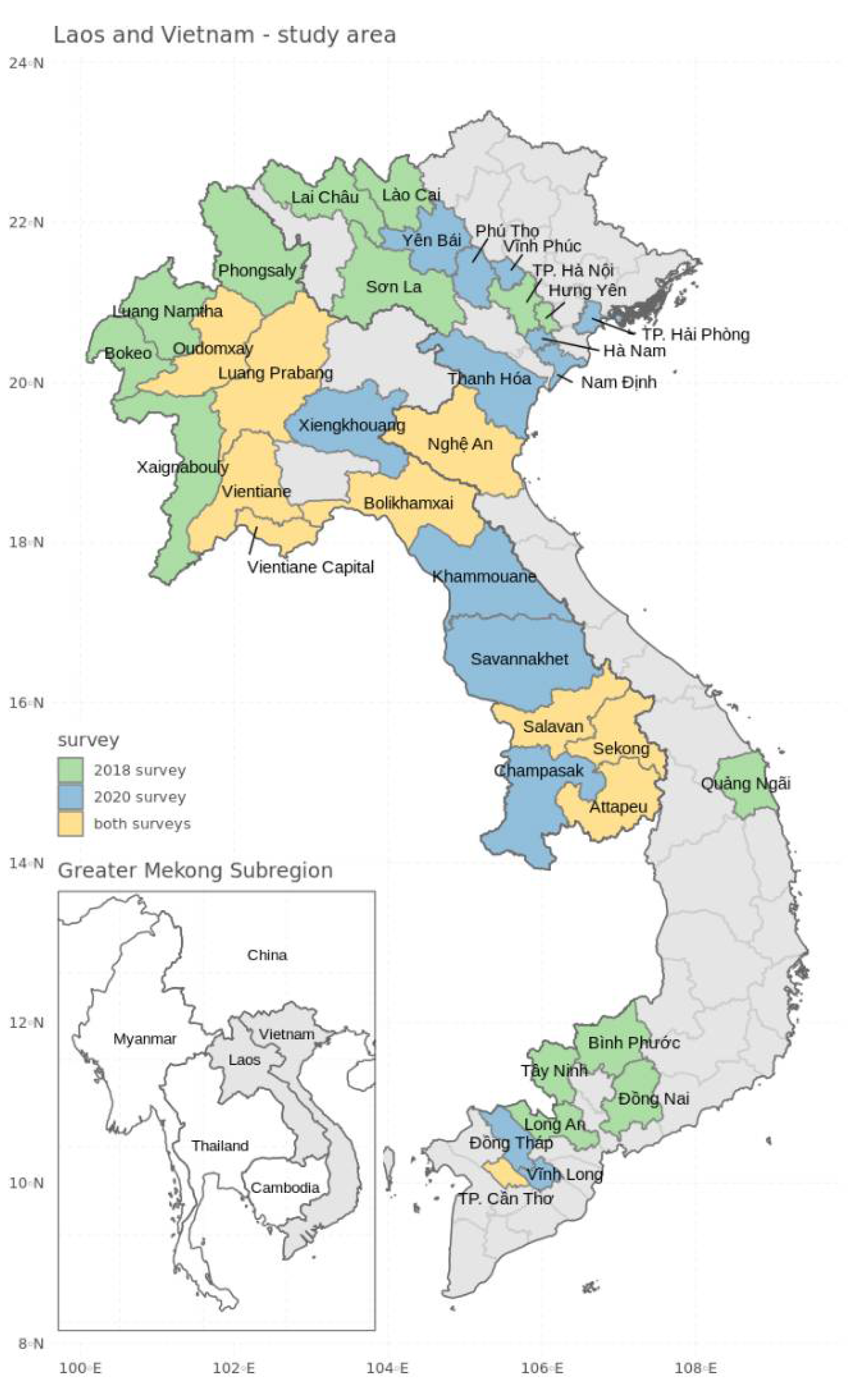
2.1. Study Area and Field Surveys
2.2. 2018 Survey: Sample Processing and Detection of Foc
2.3. 2020 Survey: Sample Processing and Detection of Foc
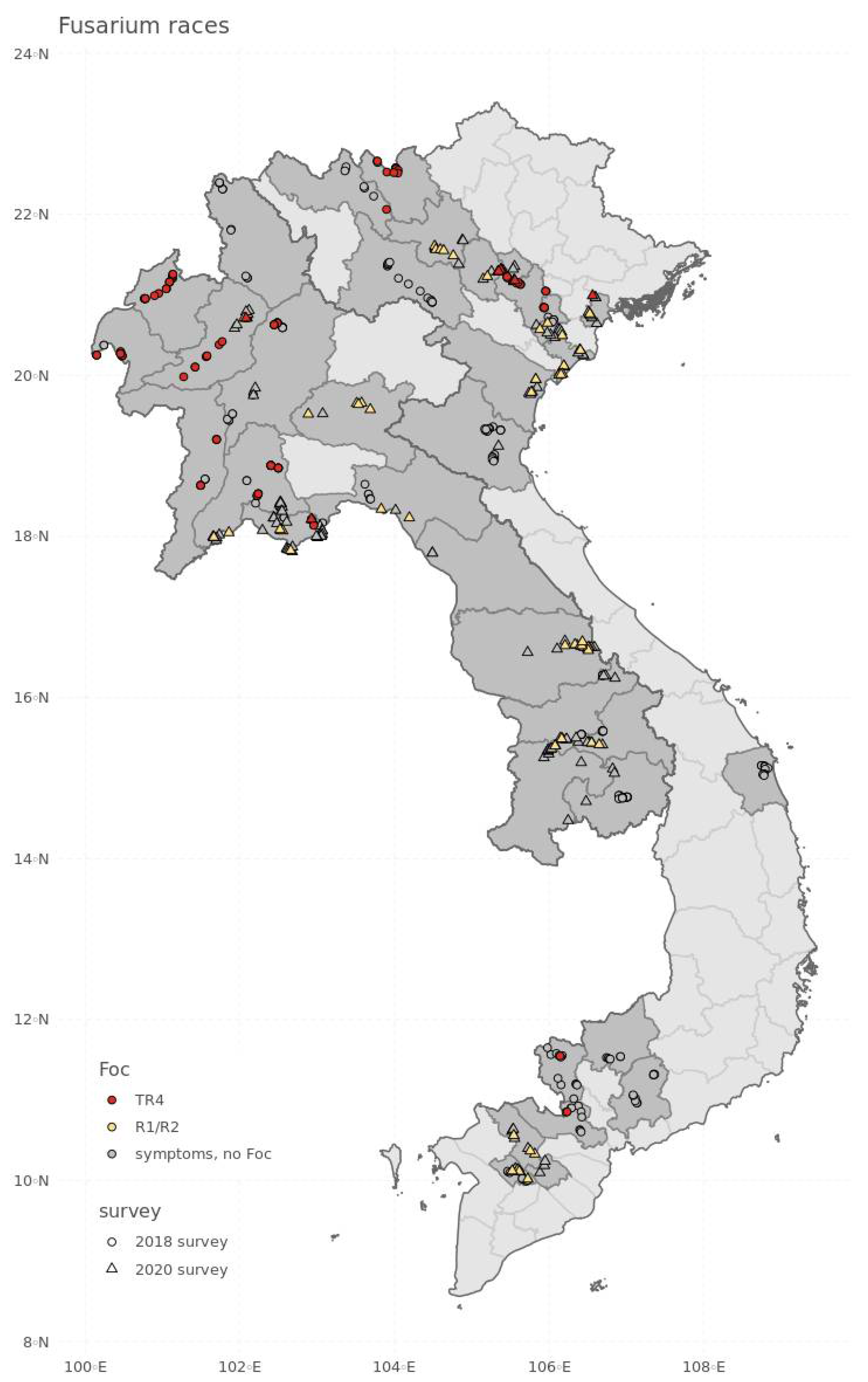
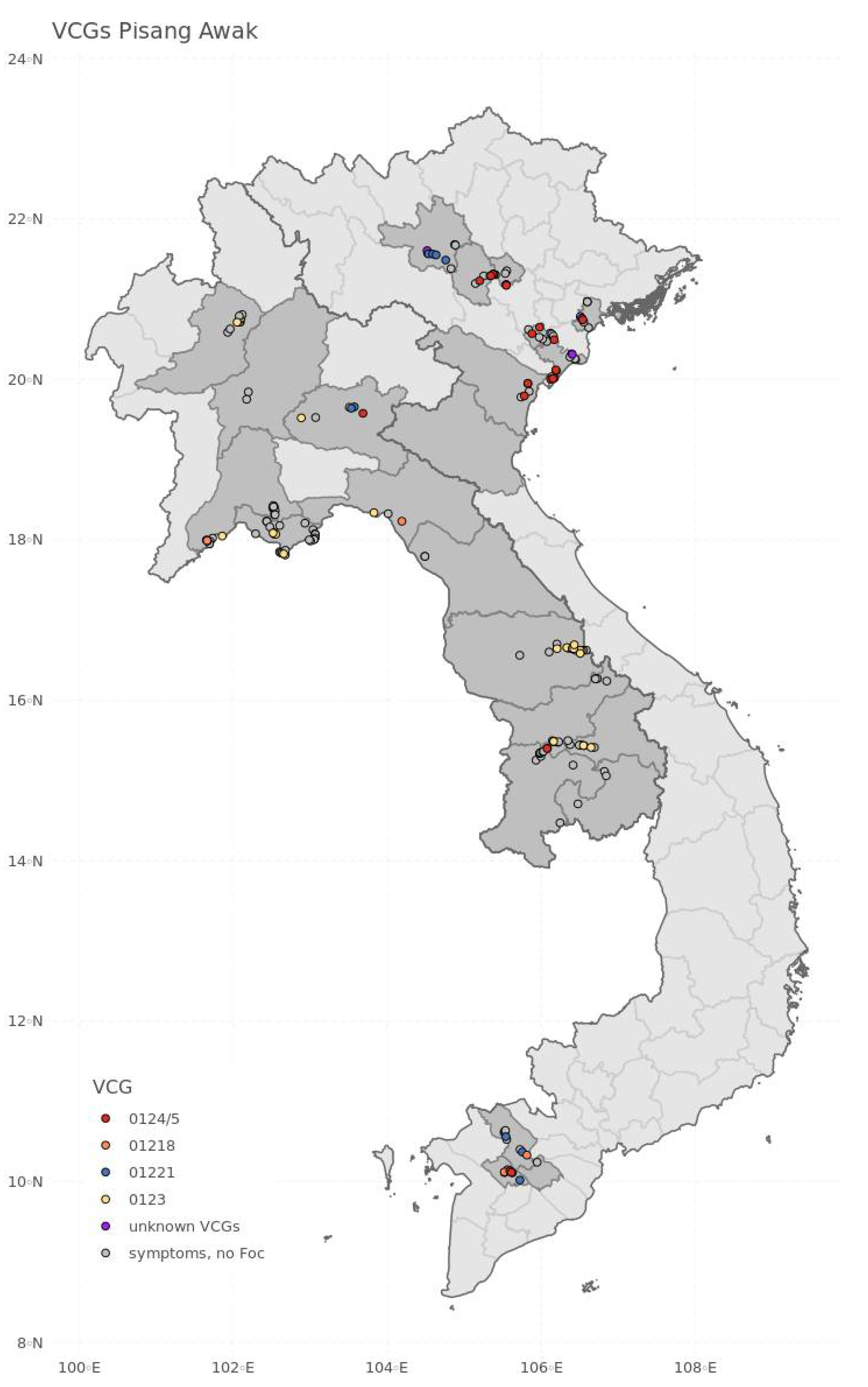
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stover, R.H.; Simmonds, N.W. Bananas, 3rd ed.; Longman: London, UK, 1987. [Google Scholar]
- Li, C.Y.; Mostert, G.; Zuo, C.W.; Beukes, I.; Yang, Q.S.; Sheng, O.; Kuang, R.B.; Wei, Y.R.; Hu, C.H.; Rose, L.; et al. Diversity and distribution of the banana wilt pathogen Fusarium oxysporum f. sp. cubense in China. Fungal Genom. Biol. 2013, 3, 111. [Google Scholar]
- Ploetz, R.C. Management of Fusarium wilt of banana: A review with special reference to 105 tropical race 4. Crop Prot. 2015, 73, 7–15. [Google Scholar] [CrossRef]
- Bai, T.T.; Xu, S.; Rupp, F.; Fan, H.; Yin, K.; Guo, Z.; Zhang, L.; Yang, B.; Huang, Y.; Li, Y.; et al. Temporal variations of Fusarium oxysporum f. sp. cubense tropical race 4 population in a heavily infected banana field in Southwest China. Acta Agric. Scand. Sect. B Soil Plant Sci. 2019, 69, 641–648. [Google Scholar] [CrossRef]
- Stover, R.H. Fusarial Wilt (Panama Disease) of Banana and Other Musa Species; Commonwealth Mycological Institute: Kew, UK, 1962; p. 117. [Google Scholar]
- Bai, T.T.; Qin, M.; Li, X.D.; Fan, H.C.; Xu, S.T.; Zeng, L.; Zheng, S.J. An additional threat to ‘Cavendish’ banana growers and traders: The infection of banana peduncles by Fusarium oxysporum f. sp. cubense Tropical Race 4 (Foc TR4). Plant Health Prog. 2020, 21, 312–316. [Google Scholar] [CrossRef]
- Pittaway, P.A.; Nasir, N.; Pegg, K.G. Soil receptivity and host–pathogen dynamics in soils naturally infested with Fusarium oxysporum f. sp. cubense, the cause of Panama disease in bananas. Aust. J. Agric. Res. 1999, 50, 623–628. [Google Scholar] [CrossRef]
- Hennessy, C.; Walduck, G.; Daly, A.; Padovan, A. Weed hosts of Fusarium oxysporum f. sp. cubense tropical race 4 in northern Australia. Australas. Plant Pathol. 2005, 34, 115–117. [Google Scholar] [CrossRef]
- Dita, M.A.; Barquero, M.; Heck, D.; Mizubuti, E.S.; Staver, C.P. Fusarium wilt of banana: Current knowledge and epidemiology on research needs toward sustainable disease management. Front. Plant Sci. 2018, 9, 1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dita, M.A.; Teixeira, L.A.J.; O’Neill, W.; Pattison, A.B.; Weinert, M.P.; Li, C.Y.; Zheng, S.J.; Staver, C.; Thangavelu, R.; Viljoen, A. Current state of Fusarium wilt of banana in the subtropics. Acta Hortic. 2020, 1272, 45–56. [Google Scholar] [CrossRef]
- Pegg, K.G.; Coates, L.M.; O’Neill, W.T.; Turner, D.W. The Epidemiology of Fusarium Wilt of Banana. Front. Plant Sci. 2019, 10, 1395. [Google Scholar] [CrossRef] [Green Version]
- Buddenhagen, I.W. Understanding strain diversity in Fusarium oxysporum f. sp. cubense and history of introduction of ‘tropical race 4′ to better manage banana production. Acta Hortic. 2009, 828, 193–204. [Google Scholar] [CrossRef]
- Siamak, S.B.; Zheng, S. Banana Fusarium wilt (Fusarium oxysporum f. sp. cubense) control and resistance, in the context of developing wilt-resistant bananas within sustainable production systems. Hortic. Plant J. 2018, 4, 208–218. [Google Scholar] [CrossRef]
- Ploetz, R.C.; Correll, J.C. Vegetative compatibility among races of Fusarium oxysporum f. sp. cubense. Plant Dis. 1988, 72, 325–328. [Google Scholar] [CrossRef]
- Bentley, S.; Dale, J.L. Genetic variation among a world-wide collection of isolates of Fusarium oxysporum f. sp. cubense analysed by RAPD-PCR fingerprinting. Mycol. Res. 1995, 99, 1378–1384. [Google Scholar] [CrossRef]
- Ploetz, R.C.; Peggy, K.G. Fusarium wilt. In Diseases of Banana, Abaca and Enset; Jones, J.R., Ed.; CABI Publishing: Wallingford, UK, 2000; pp. 143–159. [Google Scholar]
- Viljoen, A. The status of Fusarium wilt of banana in South Africa. S. Afr. J. Sci. 2002, 98, 341–344. [Google Scholar]
- Ploetz, R.C. Fusarium wilt of banana is caused by several pathogens referred to as Fusarium oxysporum f. sp. cubense. Phytopathology 2006, 96, 653–656. [Google Scholar] [CrossRef] [Green Version]
- Aguayo, J.; Mostert, D.; Fourrier-Jeandel, C.; Cerg-Wendling, I.; Hostachy, B.; Viljoen, A.; Ioos, R. Development of a hydrolysis probe-based real-time assay for the detection of tropical strains of Fusarium oxysporum f. sp. cubense race 4. PLoS ONE 2017, 12, e0171767. [Google Scholar] [CrossRef]
- Czislowski, E.; Fraser-Smith, S.; Zander, M.; O’Neill, W.T.; Meldrum, R.A.; Tran-Nguyen, L.T.; Batley, J.; Aitken, E.A. Investigation of the diversity of effector genes in the banana pathogen, Fusarium oxysporum f. sp. cubense, reveals evidence of horizontal gene transfer. Mol. Plant Pathol. 2018, 19, 1155–1171. [Google Scholar] [CrossRef] [Green Version]
- FAOSTAT. FAO Statistical Database. 2020. Available online: http://www.fao.org/ (accessed on 23 October 2021).
- Viljoen, A.; Ma, L.-J.; Molina, A.B. Fusarium wilt (Panama disease) and monoculture in banana production: Resurgence of a century-old disease. In Emerging Plant Diseases and Global Food Security; Ristaino, J.B., Records, A., Eds.; American Phytopathological Society: St. Paul, MN, USA, 2020; pp. 159–184. [Google Scholar]
- Zheng, S.-J.; García-Bastidas, F.A.; Li, X.; Zeng, L.; Bai, T.; Xu, S.; Yin, K.; Li, H.; Fu, G.; Yu, Y.; et al. New geographical insights of the latest expansion of Fusarium oxysporum f. sp. cubense tropical race 4 into the greater Mekong subregion. Front. Plant Sci. 2018, 9, 457. [Google Scholar] [CrossRef] [Green Version]
- García-Bastidas, F.; Ordóñez, N.; Konkol, J.; Al-Qasim, M.; Naser, Z.; Abdelwali, M.; Salem, N.; Waalwijk, C.; Ploetz, R.C.; Kema, G.H.J. First report of Fusarium oxysporum f. sp. cubense 86 tropical race 4 associated with Panama disease of banana outside Southeast Asia. Plant Dis. 2014, 98, 694. [Google Scholar] [CrossRef]
- García-Bastidas, F.A.; Van der Veen, A.; Nakasato-Tagami, G.; Meijer, H.J.G.; Arango-Isaza, R.E.; Kema, G.H.J. An Improved Phenotyping Protocol for Panama Disease in Banana. Front. Plant Sci. 2019, 10, 1006. [Google Scholar] [CrossRef] [Green Version]
- Maymon, M.; Shpatz, U.; Harel, Y.M.; Levy, E.; Elkind, E.; Teverovsky, E.; Gofman, R.; Haberman, A.; Zemirski, R.; Nadav, E.; et al. First report of Fusarium oxysporum f. sp. cubense tropical race 4 causing Fusarium wilt of Cavendish bananas in Israel. Plant Dis. 2018, 102, 2407–2410. [Google Scholar] [CrossRef]
- Maymon, M.; Sela, N.; Shpatz, U.; Galpaz, N.; Freeman, S. The origin and current situation of Fusarium oxysporum f. sp. cubense tropical race 4 in Israel and the Middle east. Sci. Rep. 2020, 10, 1590. [Google Scholar] [CrossRef] [PubMed]
- Aguayo, J.; Cerf-Wendling, I.; Folscher, A.B.; Fourrier-Jeandel, C.; Ioos, R.; Mathews, M.C.; Mostert, D.; Renault, C.; Wilson, V.; Viljoen, A. First report of Fusarium oxysporum f. sp. cubense tropical race 4 (TR4) causing banana wilt in the Island of Mayotte. Plant Dis. 2021, 105, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Viljoen, A.; Mostert, D.; Chiconela, T.; Beukes, I.; Fraser, C.; Dwyer, J.; Murray, H.; Amisse, J.; Mutabuana, E.L.; Tazan, G.; et al. Occurrence and spread of the banana fungus Fusarium oxyporum f. sp. cubense TR4 in Mozambique. S. Afr. J. Sci. 2020, 116, 1–11. [Google Scholar] [CrossRef]
- García-Bastidas, F.A.; Quintero-Vargas, J.C.; Ayala-Vasquez, M.; Schermer, T.; Seidl, M.F.; Santos-Paiva, M.; Noguera, A.M.; Aguilera-Galvez, C.; Wittenberg, A.; Hofstede, R.; et al. First report of Fusarium Wilt Tropical race 4 in Cavendish Bananas Caused by Fusarium odoratissimum in Colombia. Plant Dis. 2020, 104, 994. [Google Scholar] [CrossRef]
- SENASA. Confirma Brote de Fusarium Raza 4 Tropical en Piura. Available online: https://www.gob.pe/institucion/senasa/noticias/429832-senasa-confirma-brote-de-fusarium-raza-4-tropical-en-piura (accessed on 15 April 2021).
- Chittarath, K.; Mostert, D.; Crew, K.S.; Viljoen, A.; Kong, G.; Molina, A.B.; Thomas, J.E. First report of Fusarium oxysporum f. sp. cubense Tropical Race 4 (VCG 01213/16) associated with Cavendish bananas in Laos. Plant Dis. 2018, 102, 449. [Google Scholar] [CrossRef]
- Hung, T.N.; Hung, N.Q.; Mostert, D.; Viljoen, A.; Chao, C.P.; Molina, A.B. First report of Fusarium wilt on Cavendish bananas, caused by Fusarium oxysporum f. sp. cubense Tropical Race 4 (VCG 01213/16) in Vietnam. Plant Dis. 2017, 102, 448. [Google Scholar] [CrossRef]
- International Plant Protection Convection (IPPC). Detection of Fusarium oxysporum f. sp. cubense Tropical Race 4 in Thailand; Plant Quarantine Research Group; Plant Protection Research and Development Office; Department of Agriculture: Bangkok, Thailand, 2019.
- Sisanonh, S. Agricultural Statistics Year Book 2019; Ministry of Agriculture and Forestry; Department of Planning and Finance: Vientiane, Laos, 2020; p. 121.
- Viljoen, A.; Mahuku, G.S.; Massawe, C.; Tendo Ssali, R.; Kimunye, J.; Mostert, G.; Ndayihanzamaso, P.; Coyne, D. Banana Diseases and Pests: Field Guide for Diagnostics and Data Collection; IITA: Ibadan, Nigeria, 2017; p. 73. [Google Scholar]
- Hartung, C.; Lerer, A.; Anokwa, Y.; Tseng, C.; Brunette, W.; Borriello, G. Open data kit: Tools to build information services for developing regions. In Proceedings of the 4th ACM/IEEE International Conference on Information and Communication Technologies and Development–ICTD, London, UK, 13–15 December 2010. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 12 August 2021).
- Dita, M.A.; Waalwijk, C.; Buddenhagen, I.W.; Souza, M.T.; Kema, G.H.J. A molecular diagnostic for tropical race 4 of the banana Fusarium wilt pathogen. Plant Pathol. 2010, 59, 348–357. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- González-Mendoza, D.; Argumedo-Delira, R.; Morales-Trejo, A.; Pulido-Herrera, A.; Cervantes-Díaz, L.; Grimaldo-Juarez, O.; Alarcón, A. A rapid method for isolation of total DNA from pathogenic filamentous plant fungi. Genet. Mol. Res. 2010, 9, 162–166. [Google Scholar] [CrossRef]
- Matthews, M.C.; Mostert, D.; Ndayihanzamaso, P.; Rose, L.J.; Viljoen, A. Quantitative detection of economically important Fusarium oxysporum f. sp. cubense strains in Africa in 56 plants, soil and water. PLoS ONE 2020, 15, e0236110. [Google Scholar] [CrossRef] [PubMed]
- Fourie, G.; Steenkamp, E.T.; Gordon, T.R.; Viljoen, A. Evolutionary relationships among the Fusarium oxysporum f. sp. cubense vegetative compatibility groups. Appl. Environ. Microbiol. 2009, 75, 4770–4781. [Google Scholar] [CrossRef] [Green Version]
- Ndayihanzamaso, P.; Karangwa, P.; Mostert, D.; Mahuku, G.; Blomme, G.; Beed, F.; Swennen, R.; Viljoen, A. The development of a multiplex PCR assay for the detection of Fusarium oxysporum f. sp. cubense lineage VI strains in East and Central Africa. Eur. J. Plant. Pathol. 2020, 158, 495–509. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing Professional: Ames, IA, USA, 2006; p. 388. [Google Scholar]
- Kang, T. Laos Sees 76% Increase in Banana Export Value in 2019. Available online: https://laotiantimes.com/2020/02/28/laos-sees-76-increase-in-banana-export-value-in-2019/ (accessed on 14 August 2020).
- Chittarath, K. Situation of Distribution and Management of Foc TR4 in Laos; Plant Protection Center, DOA, MAF: Vientiane, Laos, 2020; pp. 2–4. [Google Scholar]
- Xu, L.B.; Huang, B.Z.; Wei, Y.R. Production and banana R&D in China. In Advancing Banana and Plantain R&D in Asia and the Pacific, Proceedings of the First BAPNET Steering Committee Meeting Held in Los Baños, Laguna, Philippines, 1–10 October 2002; Molina, A.B., Eusebio, J.E., Roa, V.N., Van den Bergh, I., Maghuyop, M.A.G., Eds.; INIBAP: Montpellier, France, 2003; Volume 11, pp. 77–80. [Google Scholar]
- Su, H.J.; Hwang, S.C.; Ko, W.H. Fusarial wilt of ‘Cavendish’ Bananas in Taiwan. Plant Dis. 1986, 70, 814–818. [Google Scholar] [CrossRef]
- Mostert, D.; Molina, A.B.; Daniells, J.; Fourie, G.; Hermanto, C.; Chao, C.-P.; Fabregar, E.; Sinohin, V.G.; Masdek, N.; Thangavelu, R.; et al. The distribution and host range of Fusarium oxysporum f. sp. cubense vegetative compatibility groups in Asia. PLoS ONE 2017, 12, e0181630. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.R. Handbook of Diseases of Banana, Abaca and Enset; CABI: Wallingford, UK, 2018; p. 632. [Google Scholar]
- Nel, B.; Steinberg, C.; Labuschagne, N.; Viljoen, A. Isolation and characterization of non-pathogenic Fusarium oxysporum isolates from the rhizosphere of healthy banana plants. Plant Pathol. 2006, 55, 207–216. [Google Scholar] [CrossRef]
- Kangire, A.; Rutherford, M. Wilt-like Disorder of Bananas in Uganda. Musa Disease Factsheet No. 10; INIBAP: Montpellier, France, 2001; p. 4. [Google Scholar]
- Maryani, N.; Sandoval-Denis, M.; Lombard, L.; Crous, P.W.; Kema, G.H.J. New endemic Fusarium species hitch-hiking with pathogenic Fusarium strains causing Panama disease in small-holder banana plots in Indonesia. Pers. Mol. Phylogeny Evol. Fungi 2019, 43, 48–69. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.C.; Ko, W.H. Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation. Plant Dis. 2004, 88, 580–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Country | Macro-Region | Region | Province | Districts | Villages | Fields | Fields TR4 Confirmed |
|---|---|---|---|---|---|---|---|
| Vietnam | Northern Vietnam | North West | Lào Cai | 3 | 4 | 10 | 8 |
| Lai Châu | 2 | 3 | 7 | 1 | |||
| Sơn La | 5 | 8 | 15 | 0 | |||
| Red River Delta | Hà Nội | 4 | 5 | 16 | 8 | ||
| Hưng Yên | 3 | 5 | 11 | 4 | |||
| Central Vietnam | North Central | * Nghệ An | 3 | 10 | 22 | 0 | |
| South Central | Quảng Ngãi | 3 | 7 | 14 | 0 | ||
| Southern Vietnam | South East | Bình Phước | 2 | 4 | 6 | 0 | |
| Đồng Nai | 3 | 4 | 10 | 0 | |||
| Tây Ninh | 5 | 8 | 12 | 1 | |||
| Mekong River Delta | Cần Thơ | 3 | 5 | 13 | 0 | ||
| Long An | 4 | 8 | 11 | 1 | |||
| Total | 40 | 71 | 147 | 23 | |||
| Laos | Northern Laos | North West | Bokeo | 2 | 6 | 14 | 10 |
| Luang Namtha | 2 | 4 | 17 | 17 | |||
| Luang Prabang | 3 | 6 | 20 | 6 | |||
| Oudomxay | 3 | 5 | 10 | 6 | |||
| Phongsaly | 3 | 4 | 10 | 0 | |||
| Xaignabouly | 2 | 2 | 10 | 7 | |||
| Central Laos | Central | Vientiane Capital | 3 | 5 | 10 | 2 | |
| Vientiane | 3 | 5 | 22 | 13 | |||
| Bolikhamxai | 1 | 3 | 6 | 0 | |||
| Southern Laos | South East | Attapeu | 2 | 3 | 17 | 0 | |
| Salavan | 2 | 2 | 11 | 0 | |||
| Sekong | 1 | 1 | 8 | 0 | |||
| Total | 27 | 46 | 155 | 61 |
| Country | Macro-Region | Region | Province | Districts | Villages | Fields | Fields TR4 Confirmed | Fields R1/R2 Confirmed |
|---|---|---|---|---|---|---|---|---|
| Vietnam | Northern Vietnam | North West | Yên Bái | 3 | 7 | 10 | 0 | 5 |
| North East | Phú Thọ | 4 | 7 | 11 | 3 | 5 | ||
| Red River Delta | Hà Nam | 5 | 9 | 10 | 0 | 3 | ||
| Hải Phòng | 2 | 7 | 10 | 1 | 2 | |||
| Nam Định | 3 | 9 | 11 | 0 | 5 | |||
| Vĩnh Phúc | 2 | 3 | 7 | 1 | 1 | |||
| Central Vietnam | North Central | * Nghệ An | 1 | 1 | 1 | 0 | 0 | |
| Thanh Hóa | 3 | 6 | 7 | 0 | 3 | |||
| Southern Vietnam | Mekong River Delta | Cần Thơ | 3 | 5 | 7 | 0 | 5 | |
| Đồng Tháp | 2 | 5 | 8 | 0 | 3 | |||
| Vĩnh Long | 2 | 3 | 3 | 0 | 0 | |||
| Total | 30 | 62 | 85 | 5 | 32 | |||
| Laos | Northern Laos | North West | Luang Prabang | 1 | 2 | 2 | 0 | 0 |
| Oudomxay | 2 | 8 | 11 | 1 | 1 | |||
| Xiangkhouang | 2 | 6 | 6 | 0 | 5 | |||
| Central Laos | Central | Vientiane Capital | 6 | 22 | 28 | 1 | 3 | |
| Vientiane | 3 | 12 | 15 | 0 | 2 | |||
| Bolikhamxai | 2 | 3 | 3 | 0 | 2 | |||
| Khammouane | 1 | 2 | 2 | 0 | 0 | |||
| Savannakhet | 3 | 16 | 19 | 0 | 9 | |||
| Southern Laos | South East | Attapeu | 2 | 3 | 4 | 0 | 0 | |
| Champasak | 1 | 6 | 10 | 0 | 1 | |||
| Salavan | 2 | 7 | 10 | 0 | 1 | |||
| Sekong | 2 | 9 | 10 | 0 | 2 | |||
| Total | 27 | 96 | 120 | 2 | 26 |
| Country | Region | Province | Banana Cultivar | Genome Group | Foc Race * | VCG | Fields |
|---|---|---|---|---|---|---|---|
| Vietnam | North West | Yên Bái | ‘Pisang Awak’ | ABB | R1 | 01221 | 4 |
| Yên Bái | ‘Pisang Awak’ | ABB | R1/R2 | Unknown VCG | 1 | ||
| North East | Phú Thọ | Cavendish | AAA | TR4 | 01213/16 | 3 | |
| Phú Thọ | ‘Pisang Awak’ | ABB | R1 | 01218 | 1 | ||
| Phú Thọ | ‘Pisang Awak’ | ABB | R1/R2 | 0124/5 | 3 | ||
| Phú Thọ | ‘Pisang Awak’ | ABB | R1/R2 | 0124 | 1 | ||
| River Delta | Hà Nam | ‘Pisang Awak’ | ABB | R1/R2 | 0124/5 | 3 | |
| Hải Phòng | Cavendish | AAA | TR4 | 01213/16 | 1 | ||
| Hải Phòng | ‘Pisang Awak’ | ABB | R1/R2 | 0124/5 | 1 | ||
| Hải Phòng | ‘Pisang Awak’ | ABB | R1/R2 | Unknown VCG | 1 | ||
| Nam Định | ‘Pisang Awak’ | ABB | R1 | Unknown VCG | 1 | ||
| Nam Định | ‘Pisang Awak’ | ABB | R1/R2 | 0124/5 | 2 | ||
| Nam Định | ‘Pisang Awak’ | ABB | R1/R2 | 0124 | 2 | ||
| Vĩnh Phúc | Cavendish | AAA | TR4 | 01213/16 | 1 | ||
| Vĩnh Phúc | ‘Pisang Awak’ | ABB | R1/R2 | 0124 | 1 | ||
| North Central | Thanh Hóa | ‘Pisang Awak’ | ABB | R1/R2 | 0124/5 | 1 | |
| Thanh Hóa | ‘Pisang Awak’ | ABB | R1/R2 | 0125 | 1 | ||
| Thanh Hóa | ‘Pisang Mas’ | AA | R1/R2 | 0124 | 1 | ||
| Mekong River Delta | Cần Thơ | ‘Pisang Awak’ | ABB | R1 | 01218 | 2 | |
| Cần Thơ | ‘Pisang Awak’ | ABB | R1 | 01221 | 1 | ||
| Cần Thơ | ‘Pisang Awak’ | ABB | R1/R2 | 0124 | 2 | ||
| Cần Thơ | ‘Pisang Awak’ | ABB | R1/R2 | Unknown VCG | 1 | ||
| Đồng Tháp | ‘Pisang Awak’ | ABB | R1 | 01218 | 1 | ||
| Đồng Tháp | ‘Pisang Awak’ | ABB | R1 | 01221 | 2 | ||
| Laos | North West | Oudomxay | Cavendish | AAA | TR4 | 01213/16 | 1 |
| Oudomxay | ‘Pisang Awak’ | ABB | R1 | 0123 | 1 | ||
| Xiangkhouang | ‘Pisang Awak’ | ABB | R1 | 01221 | 2 | ||
| Xiangkhouang | ‘Pisang Awak’ | ABB | R1 | 0123 | 2 | ||
| Xiangkhouang | ‘Pisang Awak’ | ABB | R1/R2 | 0124 | 1 | ||
| Central | Vientiane Capital | Cavendish | AAA | TR4 | 01213/16 | 1 | |
| Vientiane Capital | ‘Pisang Awak’ | ABB | R1 | 0123 | 3 | ||
| Vientiane | ‘Pisang Awak’ | ABB | R1 | 01218 | 1 | ||
| Vientiane | ‘Pisang Awak’ | ABB | R1 | 0123 | 1 | ||
| Bolikhamxai | ‘Pisang Awak’ | ABB | R1 | 01218 | 1 | ||
| Bolikhamxai | ‘Pisang Awak’ | ABB | R1 | 0123 | 1 | ||
| Savannakhet | ‘Pisang Awak’ | ABB | R1 | 0123 | 9 | ||
| South East | Champasak | ‘Pisang Awak’ | ABB | R1/R2 | 0124 | 1 | |
| Salavan | ‘Pisang Awak’ | ABB | R1 | 0123 | 1 | ||
| Sekong | ‘Pisang Awak’ | ABB | R1 | 0123 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chittarath, K.; Nguyen, C.H.; Bailey, W.C.; Zheng, S.-J.; Mostert, D.; Viljoen, A.; Tazuba, A.F.; Ocimati, W.; Kearsley, E.; Chi, T.Y.; et al. Geographical Distribution and Genetic Diversity of the Banana Fusarium Wilt Fungus in Laos and Vietnam. J. Fungi 2022, 8, 46. https://doi.org/10.3390/jof8010046
Chittarath K, Nguyen CH, Bailey WC, Zheng S-J, Mostert D, Viljoen A, Tazuba AF, Ocimati W, Kearsley E, Chi TY, et al. Geographical Distribution and Genetic Diversity of the Banana Fusarium Wilt Fungus in Laos and Vietnam. Journal of Fungi. 2022; 8(1):46. https://doi.org/10.3390/jof8010046
Chicago/Turabian StyleChittarath, Khonesavanh, Chung Huy Nguyen, Wendy C. Bailey, Si-Jun Zheng, Diane Mostert, Altus Viljoen, Anthony Fredrick Tazuba, Walter Ocimati, Elizabeth Kearsley, Trần Yến Chi, and et al. 2022. "Geographical Distribution and Genetic Diversity of the Banana Fusarium Wilt Fungus in Laos and Vietnam" Journal of Fungi 8, no. 1: 46. https://doi.org/10.3390/jof8010046
APA StyleChittarath, K., Nguyen, C. H., Bailey, W. C., Zheng, S. -J., Mostert, D., Viljoen, A., Tazuba, A. F., Ocimati, W., Kearsley, E., Chi, T. Y., Tho, N. T., Hung, N. T., Dita, M., Shah, T., Karanja, M., Mahuku, G., & Blomme, G. (2022). Geographical Distribution and Genetic Diversity of the Banana Fusarium Wilt Fungus in Laos and Vietnam. Journal of Fungi, 8(1), 46. https://doi.org/10.3390/jof8010046