
A Simple and Low-Cost Strategy to Improve Conidial Yield and Stress Resistance of Trichoderma guizhouense through Optimizing Illumination Conditions

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Light System
2.3. Quantification of Conidia
2.4. Assessment of Conidial Stress Resistance
2.5. RNA Isolation and Quantitative Real-Time PCR (qPCR)
2.6. Transcriptome Analysis
2.7. Statistical Analysis
3. Results
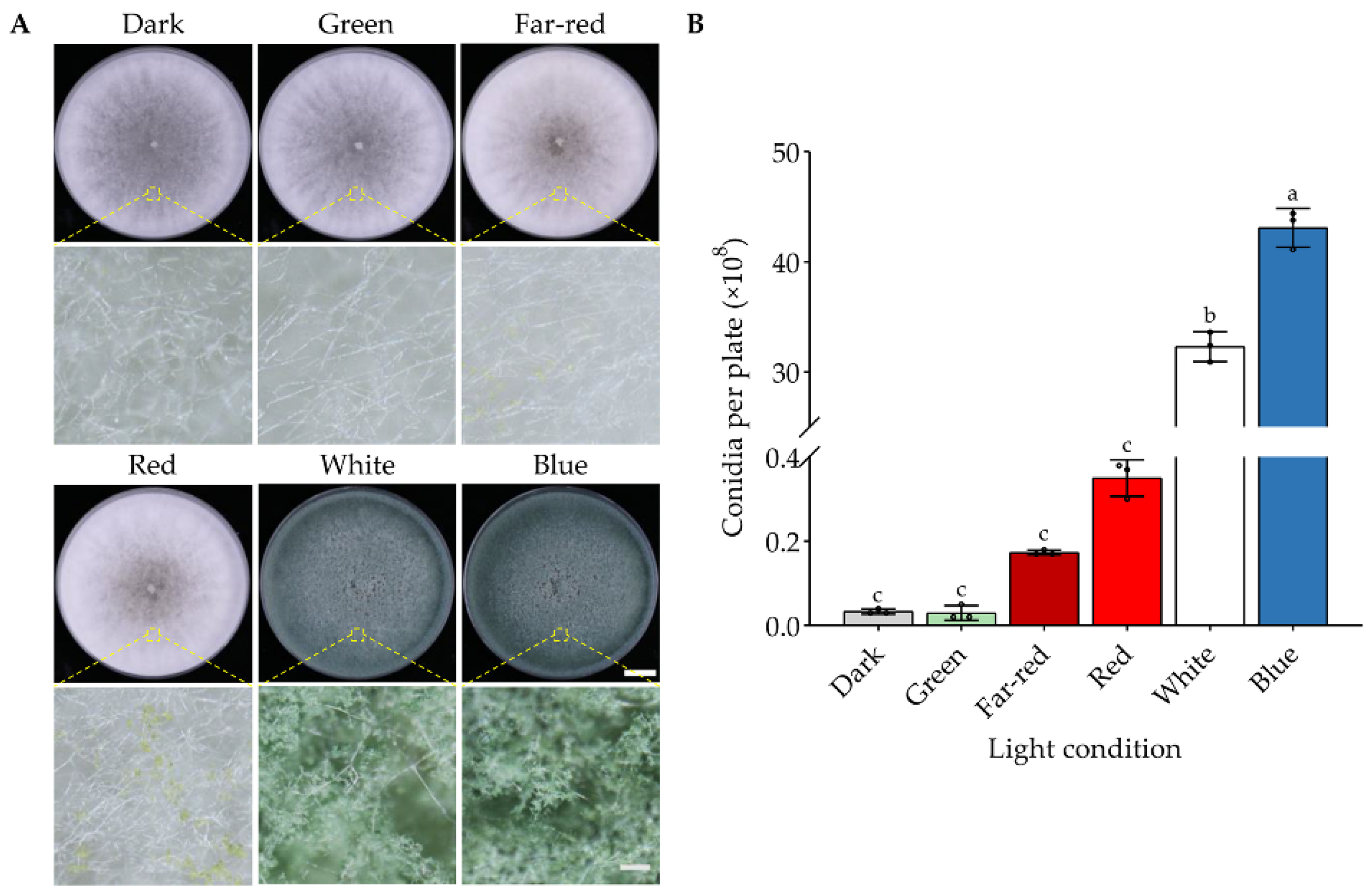
3.1. Specific Wavelengths of Light Activate Conidiation of T. guizhouense
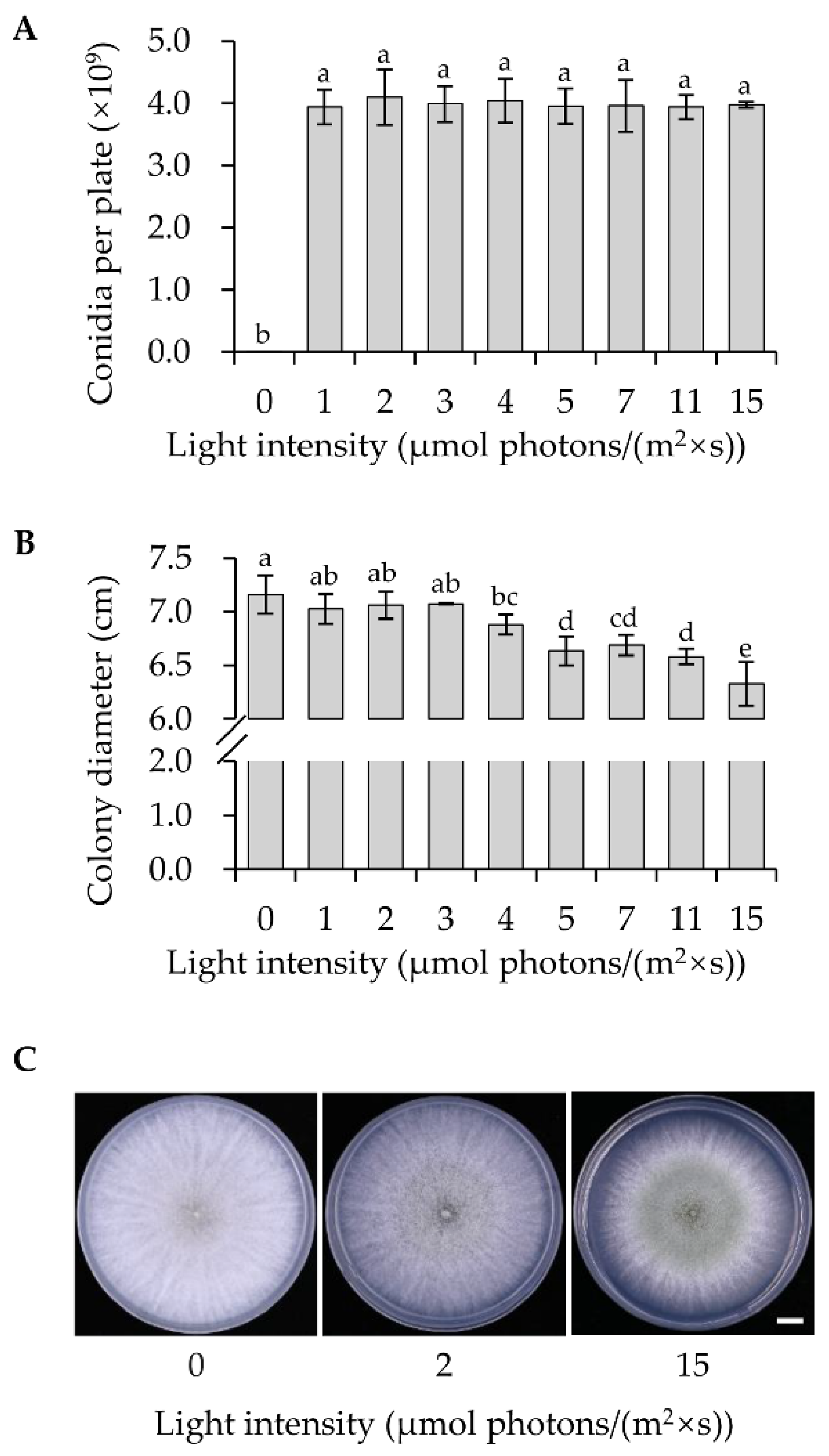
3.2. Low Intensity Blue Light Can Promote Conidial Yield Effectively
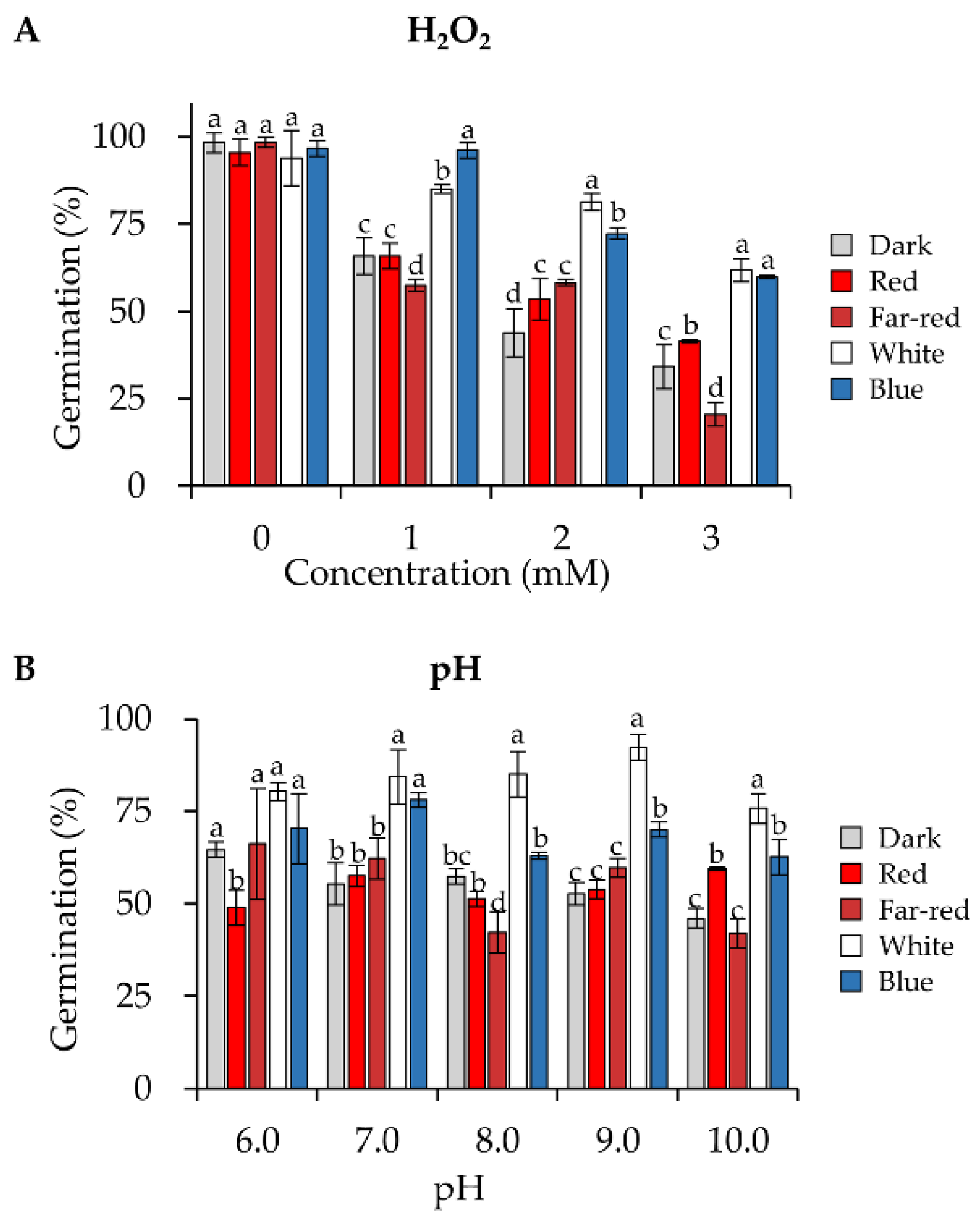
3.3. Blue and White Light Significantly Enhance the Conidial Resistance to Oxidative and pH Stresses
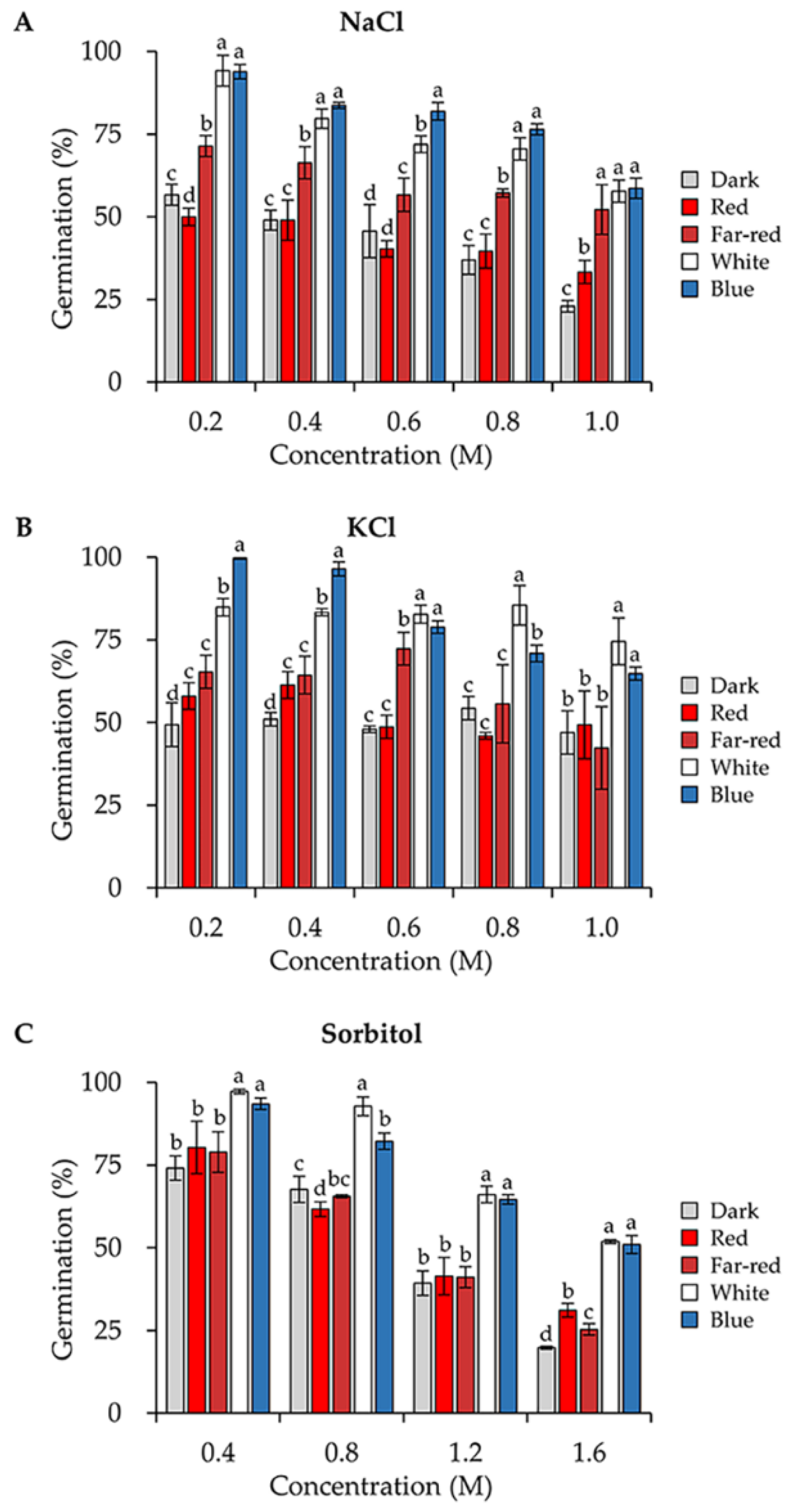
3.4. Blue and White Light Enhance the Conidial Stress Resistance to Several Osmotic Stresses
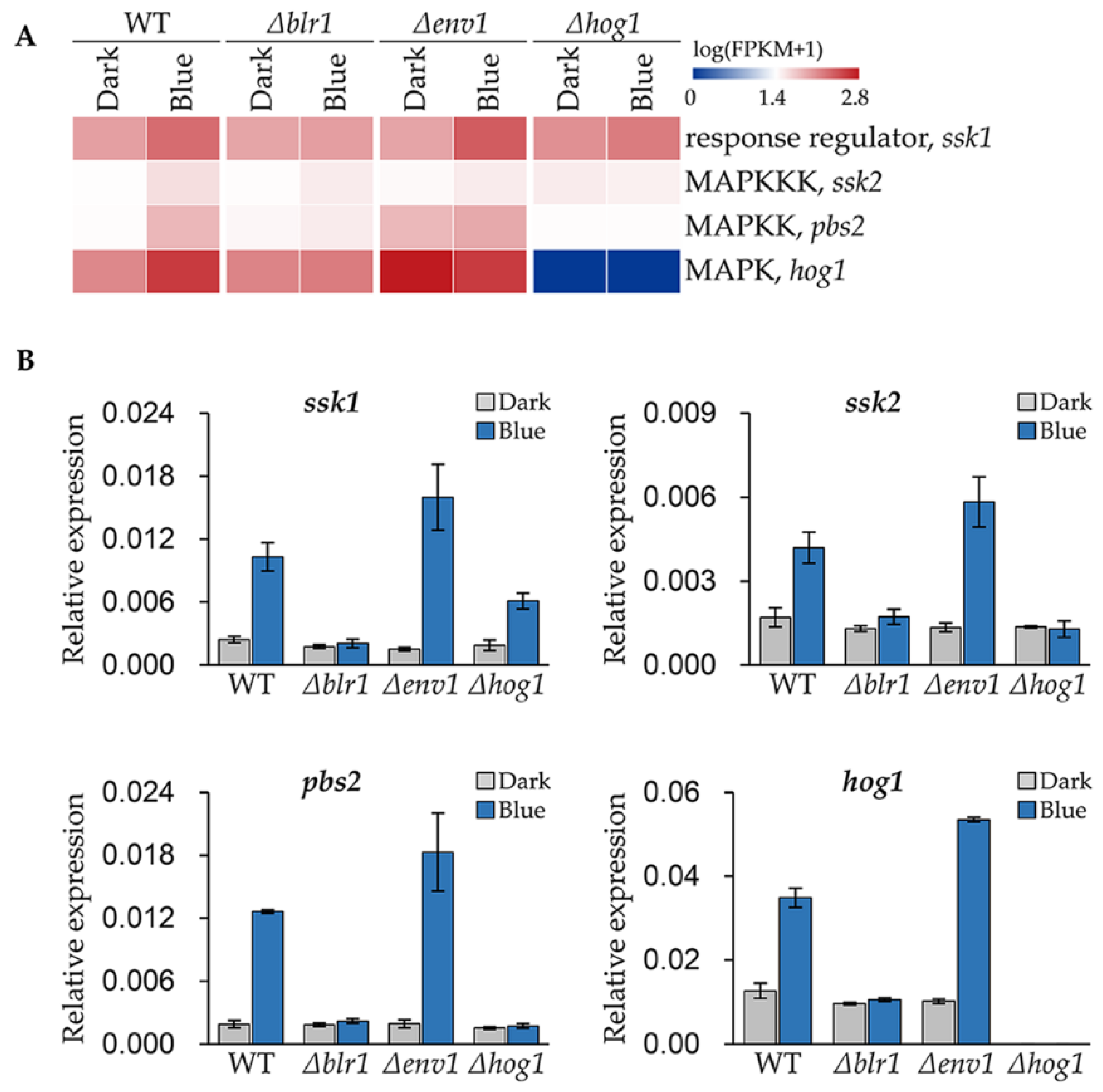
3.5. Blue Light Upregulates the Expression of the Genes Encoding HOG Pathway Components
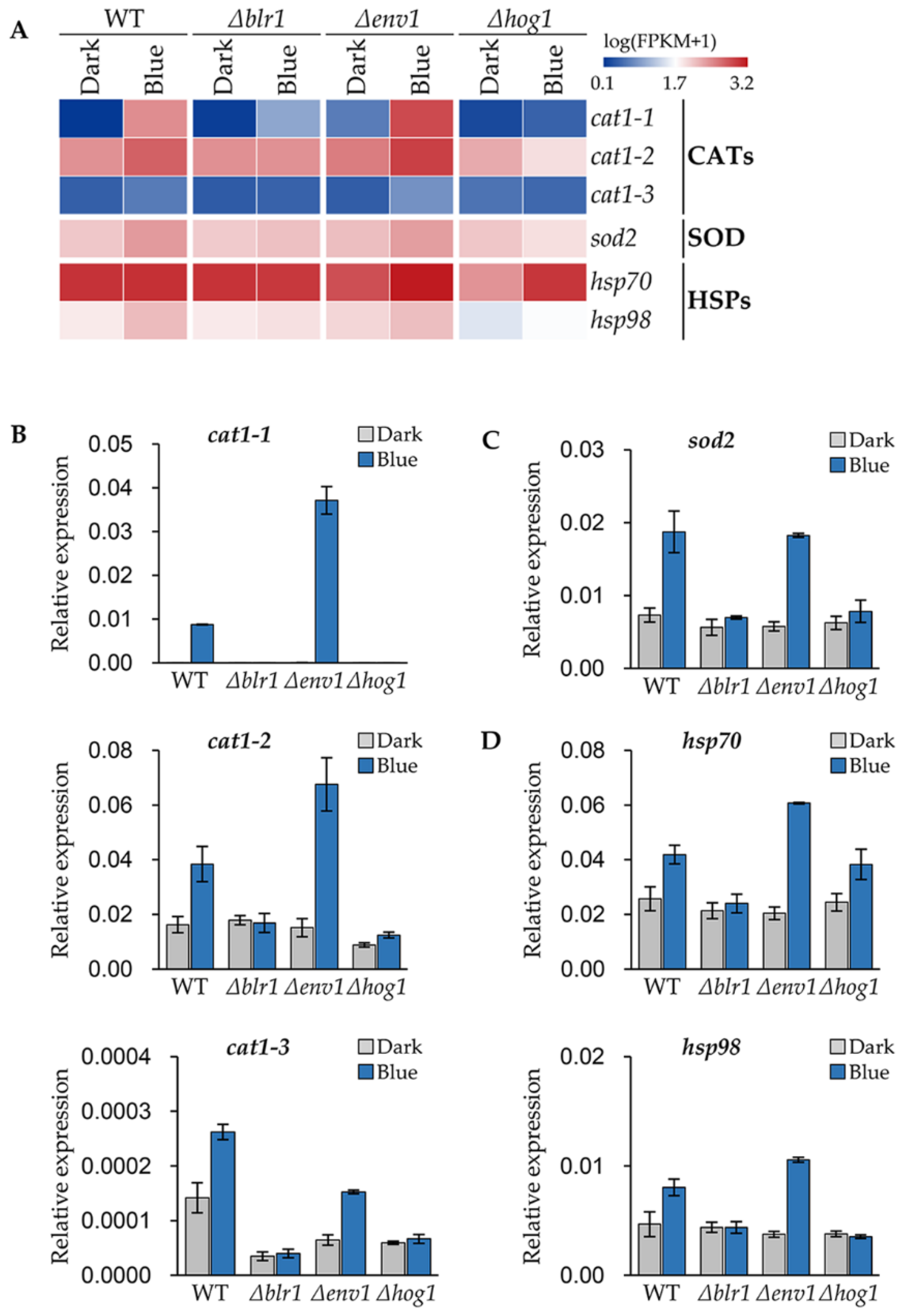
3.6. Blue Light Receptors Regulate the Stress Responsive Genes of T. guizhouense
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yu, Z.Z.; Fischer, R. Light sensing and responses in fungi. Nat. Rev. Microbiol. 2019, 17, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Blumenstein, A.; Vienken, K.; Tasler, R.; Purschwitz, J.; Veith, D.; Frankenberg-Dinkel, N.; Fischer, R. The Aspergillus nidulans phytochrome FphA represses sexual development in red light. Curr. Biol. 2005, 15, 1833–1838. [Google Scholar] [CrossRef]
- Mooney, J.L.; Yager, L.N. Light is required for conidiation in Aspergillus nidulans. Genes Dev. 1990, 4, 1473–1482. [Google Scholar] [CrossRef] [Green Version]
- Purschwitz, J.; Mueller, S.; Kastner, C.; Schoser, M.; Haas, H.; Espeso, E.A.; Atoui, A.; Calvo, A.M.; Fischer, R. Functional and physical interaction of blue- and red-light sensors in Aspergillus nidulans. Curr. Biol. 2008, 18, 255–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasgupta, A.; Fuller, K.K.; Dunlap, J.C.; Loros, J.J. Seeing the world differently: Variability in the photosensory mechanisms of two model fungi. Environ. Microbiol. 2016, 18, 5–20. [Google Scholar] [CrossRef] [Green Version]
- Schmoll, M.; Esquivel-Naranjo, E.U.; Herrera-Estrella, A. Trichoderma in the light of day—Physiology and development. Fungal Genet. Biol. 2010, 47, 909–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igbalajobi, O.; Yu, Z.Z.; Fischer, R. Red- and blue-light sensing in the plant pathogen Alternaria alternata depends on phytochrome and the white-collar protein LreA. Mbio 2019, 10, e00371-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanz, C.; Rodriguez-Romero, J.; Idnurm, A.; Christie, J.M.; Heitman, J.; Corrochano, L.M.; Eslava, A.P. Phycomyces MADB interacts with MADA to form the primary photoreceptor complex for fungal phototropism. Proc. Natl. Acad. Sci. USA 2009, 106, 7095–7100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.J.; Yu, Y.; Chen, J.; Yu, B.; Chen, T.P.; Ying, H.J.; Zhou, S.M.; Ouyang, P.K.; Liu, D.; Chen, Y. Light signaling regulates Aspergillus niger biofilm formation by affecting melanin and extracellular polysaccharide biosynthesis. Mbio 2021, 12, e03434-20. [Google Scholar] [CrossRef]
- Fuller, K.K.; Loros, J.J.; Dunlap, J.C. Fungal photobiology: Visible light as a signal for stress, space and time. Curr. Genet. 2015, 61, 275–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumacher, J.; Gorbushina, A.A. Light sensing in plant- and rock-associated black fungi. Fungal Biol. 2020, 124, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Corrochano, L.M. Light in the fungal world: From photoreception to gene transcription and beyond. Annu. Rev. Genet. 2019, 53, 149–170. [Google Scholar] [CrossRef] [PubMed]
- Bayram, O.; Biesemann, C.; Krappmann, S.; Galland, P.; Braus, G.H. More than a repair enzyme: Aspergillus nidulans photolyase-like CryA is a regulator of sexual development. Mol. Biol. Cell 2008, 19, 3254–3262. [Google Scholar] [CrossRef] [Green Version]
- Bieszke, J.A.; Braun, E.L.; Bean, L.E.; Kang, S.C.; Natvig, D.O.; Borkovich, K.A. The nop-1 gene of Neurospora crassa encodes a seven transmembrane helix retinal-binding protein homologous to archaeal rhodopsins. Proc. Natl. Acad. Sci. USA 1999, 96, 8034–8039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohrs, K.C.; Schumacher, J. The two cryptochrome/photolyase family proteins fulfill distinct roles in DNA photorepair and regulation of conidiation in the gray mold fungus Botrytis cinerea. Appl. Environ. Microbiol. 2017, 83, e00812-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Martinez, J.; Brunk, M.; Avalos, J.; Terpitz, U. The CarO rhodopsin of the fungus Fusarium fujikuroi is a light-driven proton pump that retards spore germination. Sci. Rep. 2015, 5, 7798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Wang, J.R.; Li, N.; Li, J.G.; Trail, F.; Dunlap, J.C.; Townsend, J.P. Light sensing by opsins and fungal ecology: Nop-1 modulates entry into sexual reproduction in response to environmental cues. Mol. Ecol. 2018, 27, 216–232. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, H.F.A.; Seleiman, M.F.; Al-Saif, A.M.; Alshiekheid, M.A.; Battaglia, M.L.; Taha, R.S. Biological control of celery powdery mildew disease caused by Erysiphe heraclei DC in vitro and in vivo conditions. Plants 2021, 10, 2342. [Google Scholar] [CrossRef] [PubMed]
- Druzhinina, I.S.; Seidl-Seiboth, V.; Herrera-Estrella, A.; Horwitz, B.A.; Kenerley, C.M.; Monte, E.; Mukherjee, P.K.; Zeilinger, S.; Grigoriev, I.V.; Kubicek, C.P. Trichoderma: The genomics of opportunistic success. Nat. Rev. Microbiol. 2011, 9, 749–759. [Google Scholar] [CrossRef]
- Pang, G.; Sun, T.T.; Yu, Z.Z.; Yuan, T.; Liu, W.; Zhu, H.; Gao, Q.; Yang, D.Q.; Kubicek, C.P.; Zhang, J.; et al. Azaphilones biosynthesis complements the defence mechanism of Trichoderma guizhouense against oxidative stress. Environ. Microbiol. 2020, 22, 4808–4824. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Miao, Y.Z.; Rahimi, M.J.; Zhu, H.; Steindorff, A.; Schiessler, S.; Cai, F.; Pang, G.; Chenthamara, K.; Xu, Y.; et al. Guttation capsules containing hydrogen peroxide: An evolutionarily conserved nadph oxidase gains a role in wars between related fungi. Environ. Microbiol. 2019, 21, 2644–2658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.H.; Miao, Y.Z.; Liu, Q.M.; Ma, L.; Guo, K.; Liu, D.Y.; Ran, W.; Shen, Q.R. TgSWO from Trichoderma guizhouense NJAU4742 promotes growth in cucumber plants by modifying the root morphology and the cell wall architecture. Microb. Cell Fact. 2019, 18, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, L.P.; Pedrini, N.; Braga, G.U.L.; Ferreira, P.C.; Pupin, B.; Araujo, C.A.S.; Corrochano, L.M.; Rangel, D.E.N. Outcome of blue, green, red, and white light on Metarhizium robertsii during mycelial growth on conidial stress tolerance and gene expression. Fungal Biol. 2020, 124, 263–272. [Google Scholar] [CrossRef]
- Li, Y.F.; Sun, T.T.; Guo, D.G.; Gao, J.; Zhang, J.A.; Cai, F.; Fischer, R.; Shen, Q.R.; Yu, Z.Z. Comprehensive analysis of the regulatory network of blue-light-regulated conidiation and hydrophobin production in Trichoderma guizhouense. Environ. Microbiol. 2021, 23, 6241–6256. [Google Scholar] [CrossRef]
- Esquivel-Naranjo, E.U.; Garcia-Esquivel, M.; Medina-Castellanos, E.; Correa-Perez, V.A.; Parra-Arriaga, J.L.; Landeros-Jaime, F.; Cervantes-Chavez, J.A.; Herrera-Estrella, A. A Trichoderma atroviride stress-activated MAPK pathway integrates stress and light signals. Mol. Microbiol. 2016, 100, 860–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, K.; Hoshi, Y.; Maeda, T.; Nakajima, T.; Abe, K. Aspergillus nidulans HOG pathway is activated only by two-component signalling pathway in response to osmotic stress. Mol. Microbiol. 2005, 56, 1246–1261. [Google Scholar] [CrossRef]
- Lara-Rojas, F.; Sanchez, O.; Kawasaki, L.; Aguirre, J. Aspergillus nidulans transcription factor AtfA interacts with the MAPK SakA to regulate general stress responses, development and spore functions. Mol. Microbiol. 2011, 80, 436–454. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.Z.; Armant, O.; Fischer, R. Fungi use the SakA (HogA) pathway for phytochrome-dependent light signalling. Nat. Microbiol. 2016, 1, 16019. [Google Scholar] [CrossRef]
- Yu, Z.Z.; Ali, A.; Igbalajobi, O.A.; Streng, C.; Leister, K.; Krauss, N.; Lamparter, T.; Fischer, R. Two hybrid histidine kinases, TcsB and the phytochrome FphA, are involved in temperature sensing Aspergillus nidulans. Mol. Microbiol. 2019, 112, 1814–1830. [Google Scholar] [CrossRef]
- Yu, Z.Z.; Streng, C.; Seibeld, R.F.; Igbalajobi, O.A.; Leister, K.; Ingelfinger, J.; Fischer, R. Genome-wide analyses of light-regulated genes in Aspergillus nidulans reveal a complex interplay between different photoreceptors and novel photoreceptor functions. PLoS Genet. 2021, 17, e1009845. [Google Scholar] [CrossRef]
- Dias, L.P.; Souza, R.K.F.; Pupin, B.; Rangel, D.E.N. Conidiation under illumination enhances conidial tolerance of insect-pathogenic fungi to environmental stresses. Fungal Biol. 2021, 125, 891–904. [Google Scholar] [CrossRef] [PubMed]
- Heller, J.; Tudzynski, P. Reactive oxygen species in phytopathogenic fungi: Signaling, development, and disease. Annu. Rev. Phytopathol. 2011, 49, 369–390. [Google Scholar] [CrossRef]
- Rangel, D.E.N. Stress induced cross-protection against environmental challenges on prokaryotic and eukaryotic microbes. World J. Microbiol. Biotechnol. 2011, 27, 1281–1296. [Google Scholar] [CrossRef]
- Quiroz, R.D.; Roussos, S.; Hernandez, D.; Rodriguez, R.; Castillo, F.; Aguilar, C.N. Challenges and opportunities of the bio-pesticides production by solid-state fermentation: Filamentous fungi as a model. Crit. Rev. Biotechnol. 2015, 35, 326–333. [Google Scholar] [CrossRef]
- Cai, F.; Gao, R.W.; Zhao, Z.; Ding, M.Y.; Jiang, S.Q.; Yagtu, C.; Zhu, H.; Zhang, J.; Ebner, T.; Mayrhofer-Reinhartshuber, M.; et al. Evolutionary compromises in fungal fitness: Hydrophobins can hinder the adverse dispersal of conidiospores and challenge their survival. ISME J. 2020, 14, 2610–2624. [Google Scholar] [CrossRef] [PubMed]
- Tagua, V.G.; Pausch, M.; Eckel, M.; Gutierrez, G.; Miralles-Duran, A.; Sanz, C.; Eslava, A.P.; Pokorny, R.; Corrochano, L.M.; Batschauer, A. Fungal cryptochrome with DNA repair activity reveals an early stage in cryptochrome evolution. Proc. Natl. Acad. Sci. USA 2015, 112, 15130–15135. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Meng, X.; Guo, D.; Gao, J.; Huang, Q.; Zhang, J.; Fischer, R.; Shen, Q.; Yu, Z. A Simple and Low-Cost Strategy to Improve Conidial Yield and Stress Resistance of Trichoderma guizhouense through Optimizing Illumination Conditions. J. Fungi 2022, 8, 50. https://doi.org/10.3390/jof8010050
Li Y, Meng X, Guo D, Gao J, Huang Q, Zhang J, Fischer R, Shen Q, Yu Z. A Simple and Low-Cost Strategy to Improve Conidial Yield and Stress Resistance of Trichoderma guizhouense through Optimizing Illumination Conditions. Journal of Fungi. 2022; 8(1):50. https://doi.org/10.3390/jof8010050
Chicago/Turabian StyleLi, Yifan, Xiya Meng, Degang Guo, Jia Gao, Qiwei Huang, Jian Zhang, Reinhard Fischer, Qirong Shen, and Zhenzhong Yu. 2022. "A Simple and Low-Cost Strategy to Improve Conidial Yield and Stress Resistance of Trichoderma guizhouense through Optimizing Illumination Conditions" Journal of Fungi 8, no. 1: 50. https://doi.org/10.3390/jof8010050
APA StyleLi, Y., Meng, X., Guo, D., Gao, J., Huang, Q., Zhang, J., Fischer, R., Shen, Q., & Yu, Z. (2022). A Simple and Low-Cost Strategy to Improve Conidial Yield and Stress Resistance of Trichoderma guizhouense through Optimizing Illumination Conditions. Journal of Fungi, 8(1), 50. https://doi.org/10.3390/jof8010050