
Investigations upon the Improvement of Dermatophyte Identification Using an Online Mass Spectrometry Application

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
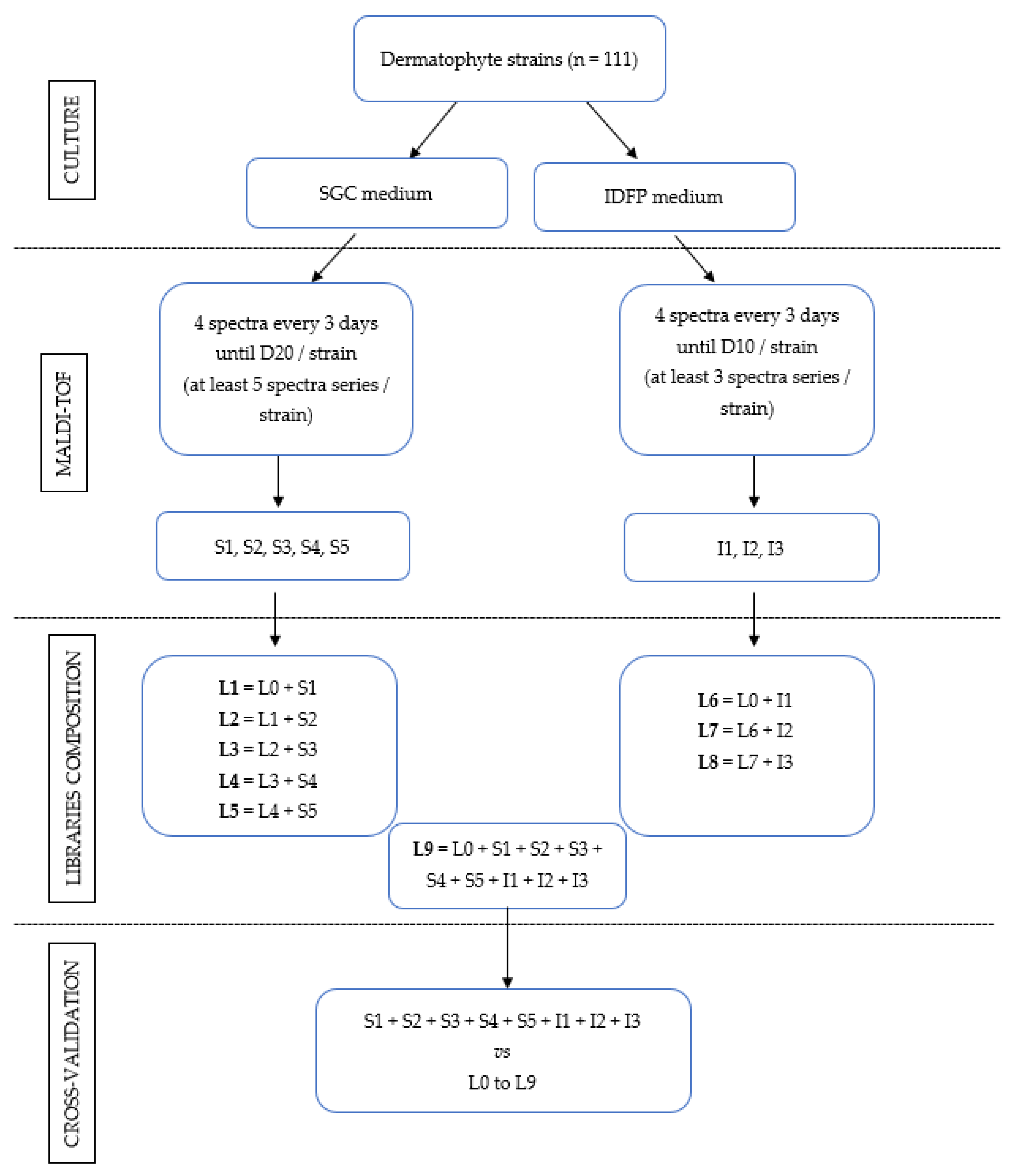
2.1. Strains and Culture
2.2. Sequencing
2.3. MALDI-TOF Mass Spectrometry
2.4. Libraries of Spectra
2.5. Cross-Validation
2.6. Statistical Analysis
3. Results
3.1. Overall Results
3.2. Results by Species and Species Complexes
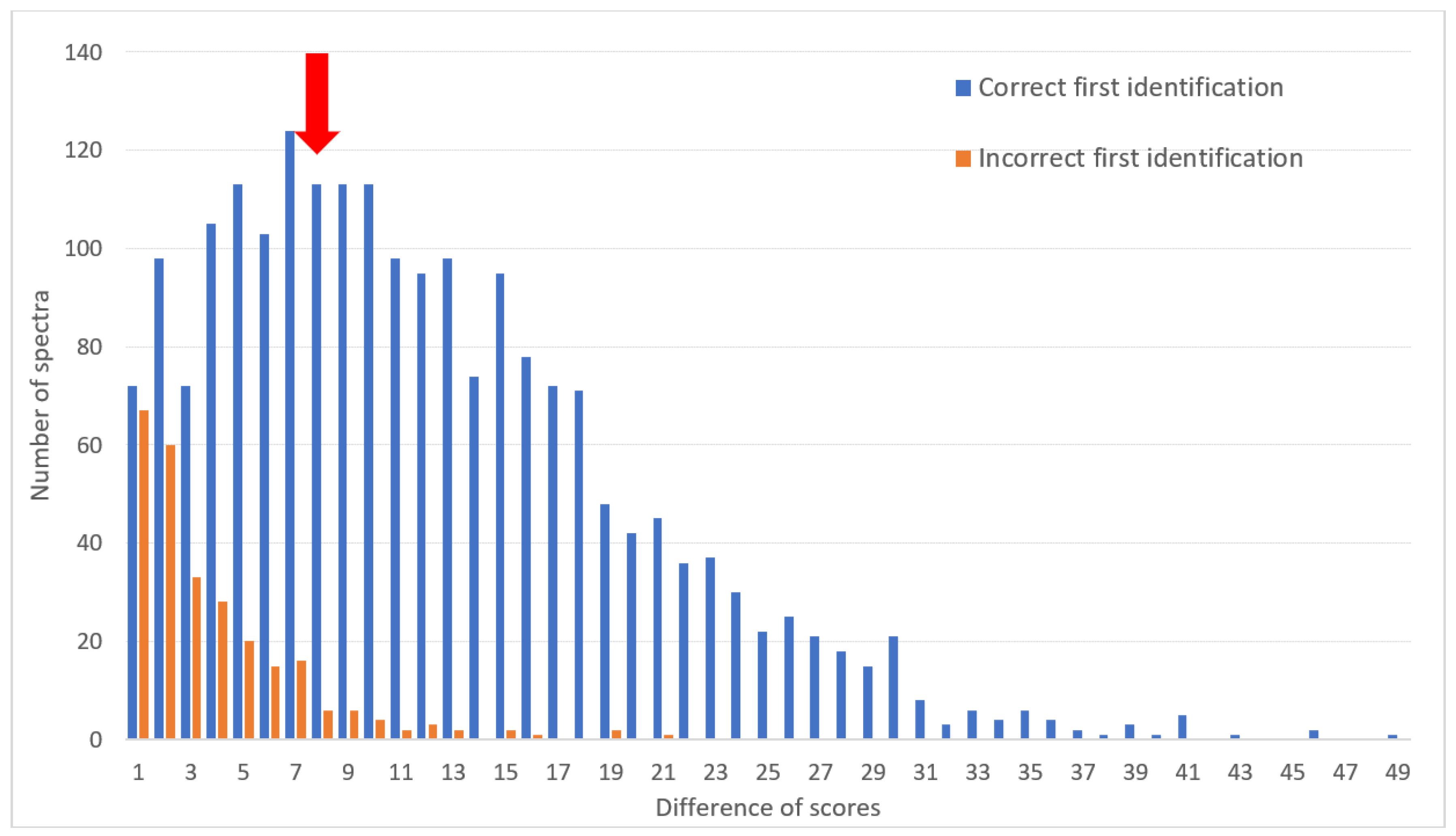
3.3. Distinction of Correct and Incorrect Identifications
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Hoog, G.S.; Dukik, K.; Monod, M.; Packeu, A.; Stubbe, D.; Hendrickx, M.; Kupsch, C.; Stielow, J.B.; Freeke, J.; Göker, M.; et al. Toward a novel multilocus phylogenetic taxonomy for the dermatophytes. Mycopathologia 2017, 182, 5–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubka, V.; Peano, A.; Cmokova, A.; Guillot, J. Common and emerging dermatophytoses in animals: Well-known and new threats. In Emerging and Epizootic Fungal Infections in Animals; Seyedmousavi, S., de Hoog, G.S., Guillot, J., Verweij, P.E., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 31–79. ISBN 978-3-319-72091-3. [Google Scholar]
- Erhard, M.; Hipler, U.-C.; Burmester, A.; Brakhage, A.A.; Wöstemeyer, J. Identification of dermatophyte species causing onychomycosis and tinea pedis by MALDI-TOF mass spectrometry. Exp. Dermatol. 2008, 17, 356–361. [Google Scholar] [CrossRef]
- L’Ollivier, C.; Ranque, S. MALDI-TOF-based dermatophyte identification. Mycopathologia 2017, 182, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Theel, E.S.; Hall, L.; Mandrekar, J.; Wengenack, N.L. Dermatophyte identification using matrix-assisted laser desorption ionization-time of flight mass spectrometry. J. Clin. Microbiol. 2011, 49, 4067–4071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Respinis, S.; Monnin, V.; Girard, V.; Welker, M.; Arsac, M.; Cellière, B.; Durand, G.; Bosshard, P.P.; Farina, C.; Passera, M.; et al. Matrix-assisted laser desorption ionization-time of flight (MALDI-TOF) mass spectrometry using the vitek MS system for rapid and accurate identification of dermatophytes on solid cultures. J. Clin. Microbiol. 2014, 52, 4286–4292. [Google Scholar] [CrossRef] [Green Version]
- L’Ollivier, C.; Cassagne, C.; Normand, A.-C.; Bouchara, J.-P.; Contet-Audonneau, N.; Hendrickx, M.; Fourquet, P.; Coulibaly, O.; Piarroux, R.; Ranque, S. A MALDI-TOF MS procedure for clinical dermatophyte species identification in the routine laboratory. Med. Mycol. 2013, 51, 713–720. [Google Scholar] [CrossRef] [Green Version]
- Alshawa, K.; Beretti, J.-L.; Lacroix, C.; Feuilhade, M.; Dauphin, B.; Quesne, G.; Hassouni, N.; Nassif, X.; Bougnoux, M.-E. Successful identification of clinical dermatophyte and neoscytalidium species by matrix-assisted laser desorption ionization-time of flight mass spectrometry. J. Clin. Microbiol. 2012, 50, 2277–2281. [Google Scholar] [CrossRef] [Green Version]
- Hedayati, M.T.; Ansari, S.; Ahmadi, B.; Taghizadeh Armaki, M.; Shokohi, T.; Abastabar, M.; Er, H.; Özhak, B.; Öğünç, D.; Ilkit, M.; et al. Identification of clinical dermatophyte isolates obtained from Iran by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Curr. Med. Mycol. 2019, 5, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Packeu, A.; Hendrickx, M.; Beguin, H.; Martiny, D.; Vandenberg, O.; Detandt, M. Identification of the Trichophyton Mentagrophytes complex species using MALDI-TOF mass spectrometry. Med. Mycol. 2013, 51, 580–585. [Google Scholar] [CrossRef] [Green Version]
- Gräser, Y.; Kuijpers, A.F.; Presber, W.; de Hoog, G.S. Molecular taxonomy of the trichophyton rubrum complex. J. Clin. Microbiol. 2000, 38, 3329–3336. [Google Scholar] [CrossRef] [Green Version]
- Gräser, Y.; Kuijpers, A.F.; Presber, W.; De Hoog, G.S. Molecular taxonomy of Trichophyton Mentagrophytes and T. Tonsurans. Med. Mycol. 1999, 37, 315–330. [Google Scholar] [CrossRef]
- Beguin, H.; Pyck, N.; Hendrickx, M.; Planard, C.; Stubbe, D.; Detandt, M. The Taxonomic status of Trichophyton Quinckeanum and T. Interdigitale revisited: A multigene phylogenetic approach. Med. Mycol. 2012, 50, 871–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodgyer, A. The curious adventures of Trichophyton Equinum in the realm of molecular biology: A modern fairy tale. Med. Mycol. 2004, 42, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Su, H.; Packeu, A.; Ahmed, S.A.; Al-Hatmi, A.M.S.; Blechert, O.; İlkit, M.; Hagen, F.; Gräser, Y.; Liu, W.; Deng, S.; et al. Species distinction in the Trichophyton Rubrum complex. J. Clin. Microbiol. 2019, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Packeu, A.; Stubbe, D.; Roesems, S.; Goens, K.; Van Rooij, P.; de Hoog, S.; Hendrickx, M. Lineages within the Trichophyton Rubrum complex. Mycopathologia 2020, 185, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Kandemir, H.; Dukik, K.; Hagen, F.; Ilkit, M.; Gräser, Y.; de Hoog, G.S. Polyphasic discrimination of Trichophyton Tonsurans and T. Equinum from humans and horses. Mycopathologia 2020, 185, 113–122. [Google Scholar] [CrossRef]
- Taghipour, S.; Pchelin, I.M.; Zarei Mahmoudabadi, A.; Ansari, S.; Katiraee, F.; Rafiei, A.; Shokohi, T.; Abastabar, M.; Taraskina, A.E.; Kermani, F.; et al. Trichophyton Mentagrophytes and T Interdigitale genotypes are associated with particular geographic areas and clinical manifestations. Mycoses 2019, 62, 1084–1091. [Google Scholar] [CrossRef]
- Gräser, Y.; Kuijpers, A.F.; El Fari, M.; Presber, W.; de Hoog, G.S. Molecular and conventional taxonomy of the microsporum Canis complex. Med. Mycol. 2000, 38, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Rezaei-Matehkolaei, A.; Makimura, K.; de Hoog, G.S.; Shidfar, M.R.; Satoh, K.; Najafzadeh, M.J.; Mirhendi, H. Multilocus differentiation of the related dermatophytes Microsporum Canis, Microsporum Ferrugineum and Microsporum Audouinii. J. Med. Microbiol. 2012, 61, 57–63. [Google Scholar] [CrossRef]
- Normand, A.C.; Becker, P.; Gabriel, F.; Cassagne, C.; Accoceberry, I.; Gari-Toussaint, M.; Hasseine, L.; De Geyter, D.; Pierard, D.; Surmont, I.; et al. Validation of a new web application for identification of fungi by use of matrix-assisted laser desorption ionization–time of flight mass spectrometry. J. Clin. Microbiol. 2017, 55, 2661–2670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, P.; Normand, A.-C.; Vanantwerpen, G.; Vanrobaeys, M.; Haesendonck, R.; Vercammen, F.; Stubbe, D.; Piarroux, R.; Hendrickx, M. Identification of fungal isolates by MALDI-TOF mass spectrometry in veterinary practice: Validation of a web application. J. Vet. Diagn. Invest. 2019, 31, 471–474. [Google Scholar] [CrossRef] [PubMed]
- Heireman, L.; Patteet, S.; Steyaert, S. Performance of the new ID-fungi plate using two types of reference libraries (Bruker and MSI) to identify fungi with the Bruker MALDI biotyper. Med. Mycol. 2020, 58, 946–957. [Google Scholar] [CrossRef]
- Sacheli, R.; Henri, A.-S.; Seidel, L.; Ernst, M.; Darfouf, R.; Adjetey, C.; Schyns, M.; Marechal, L.; Meex, C.; Arrese, J.; et al. Evaluation of the new Id-fungi plates from conidia for MALDI-TOF MS identification of filamentous fungi and comparison with conventional methods as identification tool for dermatophytes from nails, hair and skin samples. Mycoses 2020, 63, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Normand, A.-C.; Blaize, M.; Imbert, S.; Packeu, A.; Becker, P.; Fekkar, A.; Stubbe, D.; Piarroux, R. Identification of Molds with matrix-assisted laser desorption ionization-time of flight mass spectrometry: Performance of the newly developed MSI-2 Application in comparison with the Bruker filamentous fungi database and MSI-1. J. Clin. Microbiol. 2021, 59, e0129921. [Google Scholar] [CrossRef]
- Normand, A.C.; Packeu, A.; Cassagne, C.; Hendrickx, M.; Ranque, S.; Piarroux, R. Nucleotide sequence database comparison for routine dermatophyte identification by internal transcribed spacer 2 genetic region DNA barcoding. J. Clin. Microbiol. 2018, 56, e00046-18. [Google Scholar] [CrossRef] [Green Version]
- Summerbell, R.C.; Moore, M.K.; Starink-Willemse, M.; Van Iperen, A. ITS barcodes for trichophyton tonsurans and T. Equinum. Med. Mycol. 2007, 45, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Kupsch, C.; Czaika, V.-A.; Deutsch, C.; Gräser, Y. Trichophyton Mentagrophytes—A New genotype of zoophilic dermatophyte causes sexually transmitted infections. J. Dtsch. Dermatol. Ges. 2019, 17, 493–501. [Google Scholar] [CrossRef]
- Nenoff, P.; Verma, S.B.; Vasani, R.; Burmester, A.; Hipler, U.-C.; Wittig, F.; Krüger, C.; Nenoff, K.; Wiegand, C.; Saraswat, A.; et al. The current Indian epidemic of superficial dermatophytosis due to trichophyton mentagrophytes—A molecular study. Mycoses 2019, 62, 336–356. [Google Scholar] [CrossRef] [PubMed]
- Kano, R.; Kimura, U.; Kakurai, M.; Hiruma, J.; Kamata, H.; Suga, Y.; Harada, K. Trichophyton Indotineae Sp. Nov.: A new highly terbinafine-resistant anthropophilic dermatophyte species. Mycopathologia 2020, 185, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Kong, X.; Ahmed, S.A.; Thakur, R.; Chowdhary, A.; Nenoff, P.; Uhrlass, S.; Verma, S.B.; Meis, J.F.; Kandemir, H.; et al. Taxonomy of the Trichophyton Mentagrophytes/T. Interdigitale Species Complex Harboring the Highly Virulent, Multiresistant genotype T. Indotineae. Mycopathologia 2021, 186, 315–326. [Google Scholar] [CrossRef]
- Seebacher, C.; Bouchara, J.-P.; Mignon, B. Updates on the epidemiology of dermatophyte infections. Mycopathologia 2008, 166, 335–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baert, F.; Lefevere, P.; D’hooge, E.; Stubbe, D.; Packeu, A. A Polyphasic approach to classification and identification of species within the trichophyton benhamiae complex. JoF 2021, 7, 602. [Google Scholar] [CrossRef] [PubMed]
- Bergmans, A.M.C.; van der Ent, M.; Klaassen, A.; Böhm, N.; Andriesse, G.I.; Wintermans, R.G.F. Evaluation of a single-tube real-time PCR for detection and identification of 11 dermatophyte species in clinical material. Clin. Microbiol. Infect. 2010, 16, 704–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Libraries | L0 | L1 | L2 | L3 | L4 | L5 | L6 | L7 | L8 | L9 |
|---|---|---|---|---|---|---|---|---|---|---|
| SGC Libraries | IDFP Libraries | |||||||||
| Composition of the libraries | MSI-2 references | L0 + S1 | L1 + S2 | L2 + S3 | L3 + S4 | L4 + S5 | L0 + I1 | L6 + I2 | L7 + I3 | L0 + S + I |
| Correct identification | 63.4 | 77.5 | 85.9 | 86.9 | 87.6 | 88.9 | 74.8 | 84.3 | 85.8 | 91.7 |
| Incorrect identification | 32.1 | 20.8 | 13.2 | 12.4 | 11.8 | 10.5 | 22.7 | 14.4 | 13.3 | 7.8 |
| No identification | 4.5 | 1.7 | 0.9 | 0.8 | 0.6 | 0.6 | 2.5 | 1.2 | 0.9 | 0.5 |
| Libraries | L0 | L9 |
|---|---|---|
| Composition of the Libraries | MSI-2 References | L0 + S + I |
| Epidermophyton floccosum (n = 120) | 56.7 | 100 |
| M. audouinii (n = 360) | 57.2 | 96.9 |
| M. canis (n = 236) | 86.0 | 99.6 |
| N. fulva (n = 65) | 75.4 | 100 |
| N. gypsea (n = 198) | 100 | 100 |
| N. persicolor (n = 76) | 97.4 | 97.4 |
| T. benhamiae (n = 100) | 81.0 | 97.0 |
| T. equinum (n = 258) | 28.0 | 89.3 |
| T. erinacei (n = 168) | 96.9 | 100 |
| T. interdigitale (n = 439) | 63.6 | 81.8 |
| Trichophyton kuryangei (n = 96) | 62.5 | 77.1 |
| T. mentagrophytes (n = 352) | 71.6 | 88.1 |
| T. rubrum (n = 454) | 84.1 | 91.6 |
| T. soudanense (n = 348) | 2.6 | 94.3 |
| T. tonsurans (n = 442) | 60.2 | 91.2 |
| T. verrucosum (n = 48) | 0.0 | 50.0 |
| T. violaceum (n = 92) | 19.6 | 80.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jabet, A.; Normand, A.-C.; Moreno-Sabater, A.; Guillot, J.; Risco-Castillo, V.; Brun, S.; Demar, M.; Blaizot, R.; Nabet, C.; Packeu, A.; et al. Investigations upon the Improvement of Dermatophyte Identification Using an Online Mass Spectrometry Application. J. Fungi 2022, 8, 73. https://doi.org/10.3390/jof8010073
Jabet A, Normand A-C, Moreno-Sabater A, Guillot J, Risco-Castillo V, Brun S, Demar M, Blaizot R, Nabet C, Packeu A, et al. Investigations upon the Improvement of Dermatophyte Identification Using an Online Mass Spectrometry Application. Journal of Fungi. 2022; 8(1):73. https://doi.org/10.3390/jof8010073
Chicago/Turabian StyleJabet, Arnaud, Anne-Cécile Normand, Alicia Moreno-Sabater, Jacques Guillot, Veronica Risco-Castillo, Sophie Brun, Magalie Demar, Romain Blaizot, Cécile Nabet, Ann Packeu, and et al. 2022. "Investigations upon the Improvement of Dermatophyte Identification Using an Online Mass Spectrometry Application" Journal of Fungi 8, no. 1: 73. https://doi.org/10.3390/jof8010073
APA StyleJabet, A., Normand, A. -C., Moreno-Sabater, A., Guillot, J., Risco-Castillo, V., Brun, S., Demar, M., Blaizot, R., Nabet, C., Packeu, A., & Piarroux, R. (2022). Investigations upon the Improvement of Dermatophyte Identification Using an Online Mass Spectrometry Application. Journal of Fungi, 8(1), 73. https://doi.org/10.3390/jof8010073