
The Small Ras Superfamily GTPase Rho4 of the Maize Anthracnose Fungus Colletotrichum graminicola Is Required for β-1,3-glucan Synthesis, Cell Wall Integrity, and Full Virulence
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains, Culture Conditions, Hyphopodium Differentiation, and Virulence Assays
2.2. Genome Mining, Phylogeny, and Identification of Conserved Domains
2.3. Bioinformatics
2.4. Targeted Deletion of C. graminicola RHO4
2.5. DNA Isolation and Genomic Southern Blot Analyses
2.6. Microscopy and Ratio Imaging
2.7. Statistical Analyses
2.8. Protein Accession Numbers
3. Results
3.1. The Genome of C. graminicola Harbors a Single-Copy Gene Encoding a Rho4-like Protein
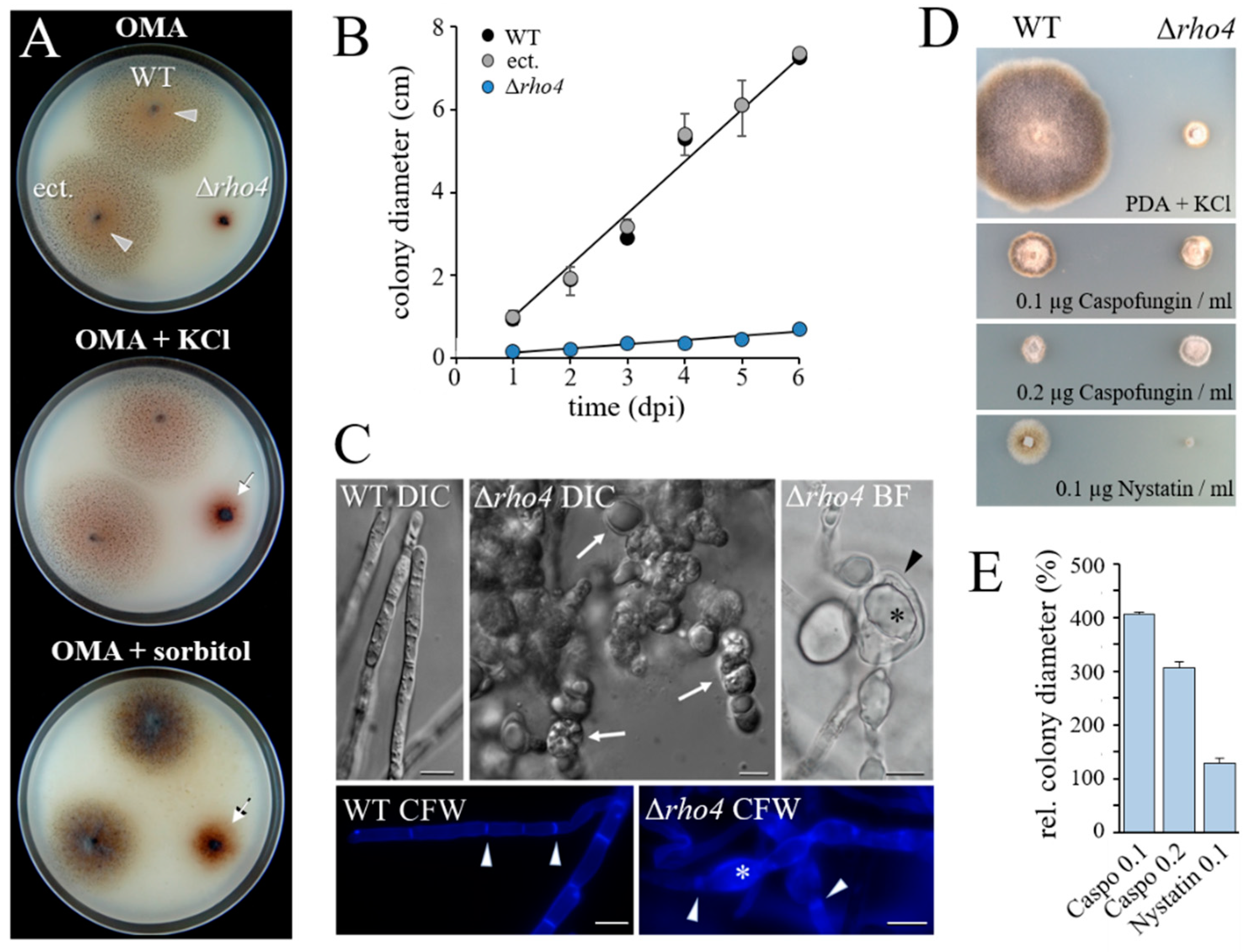
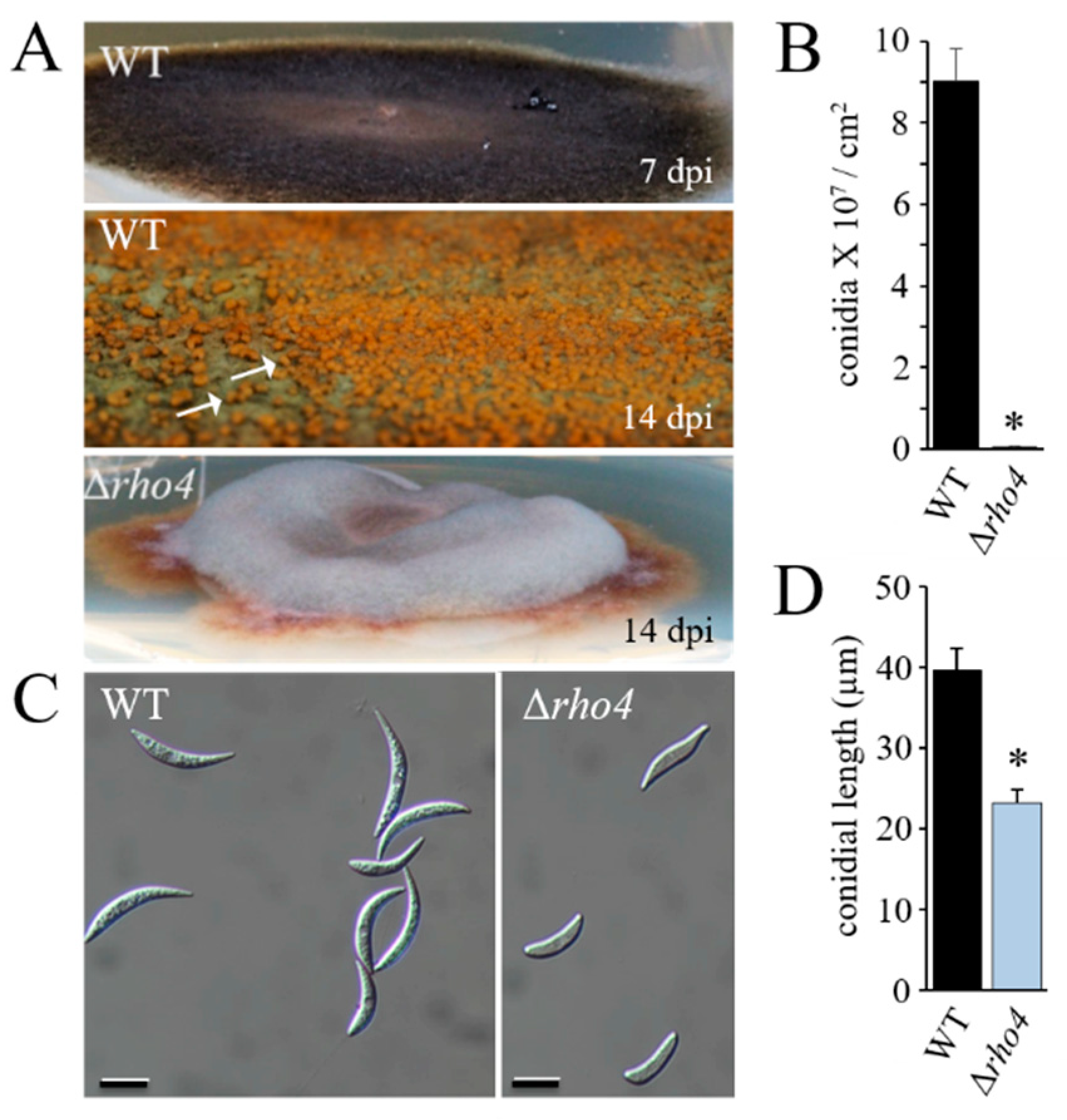
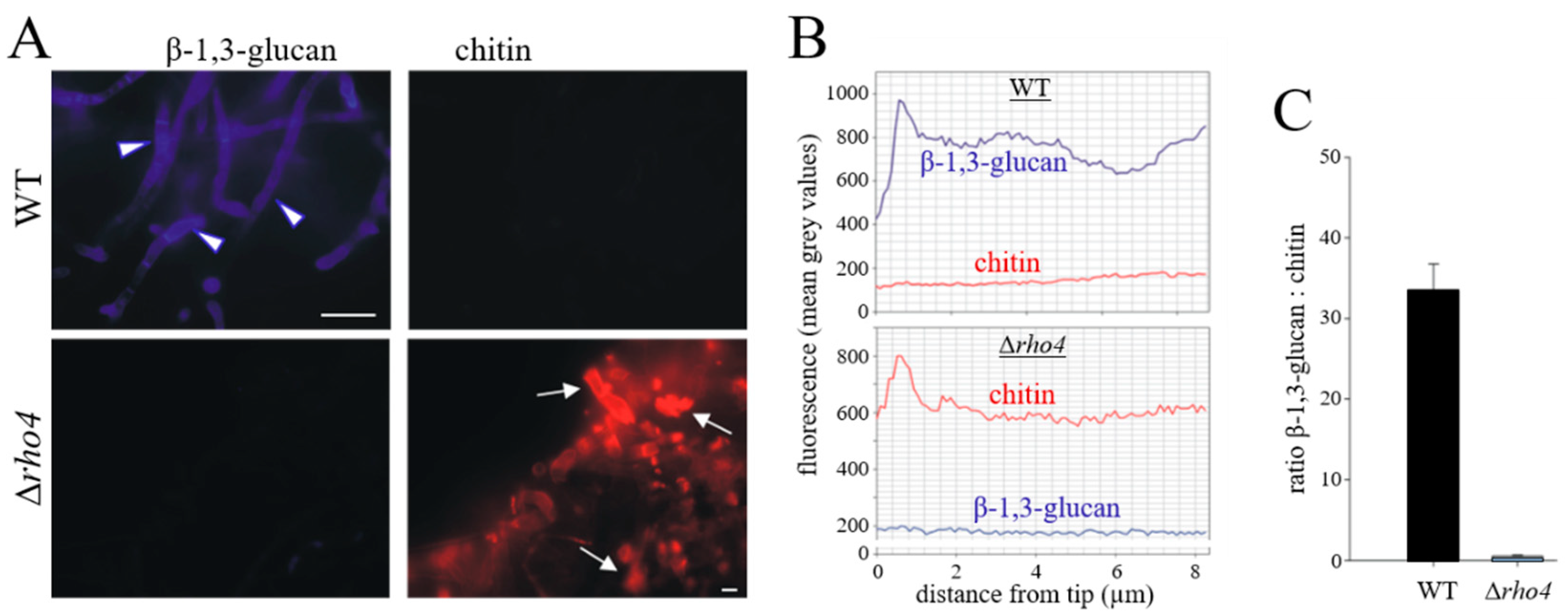
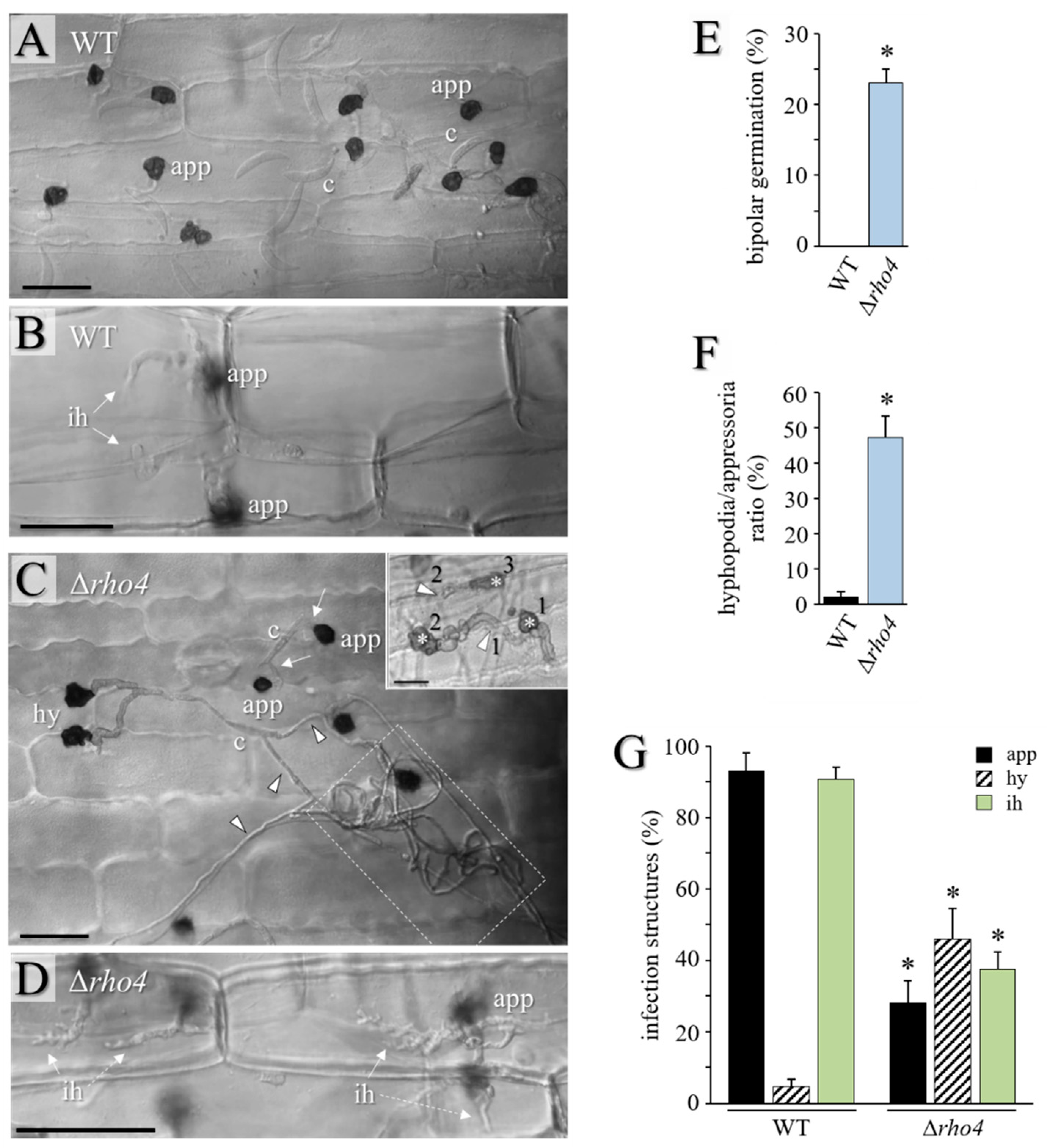
3.2. RHO4-Deficient Mutants Exhibit Phenotypic Similarities with GLS1-RNAi Strains of C. graminicola
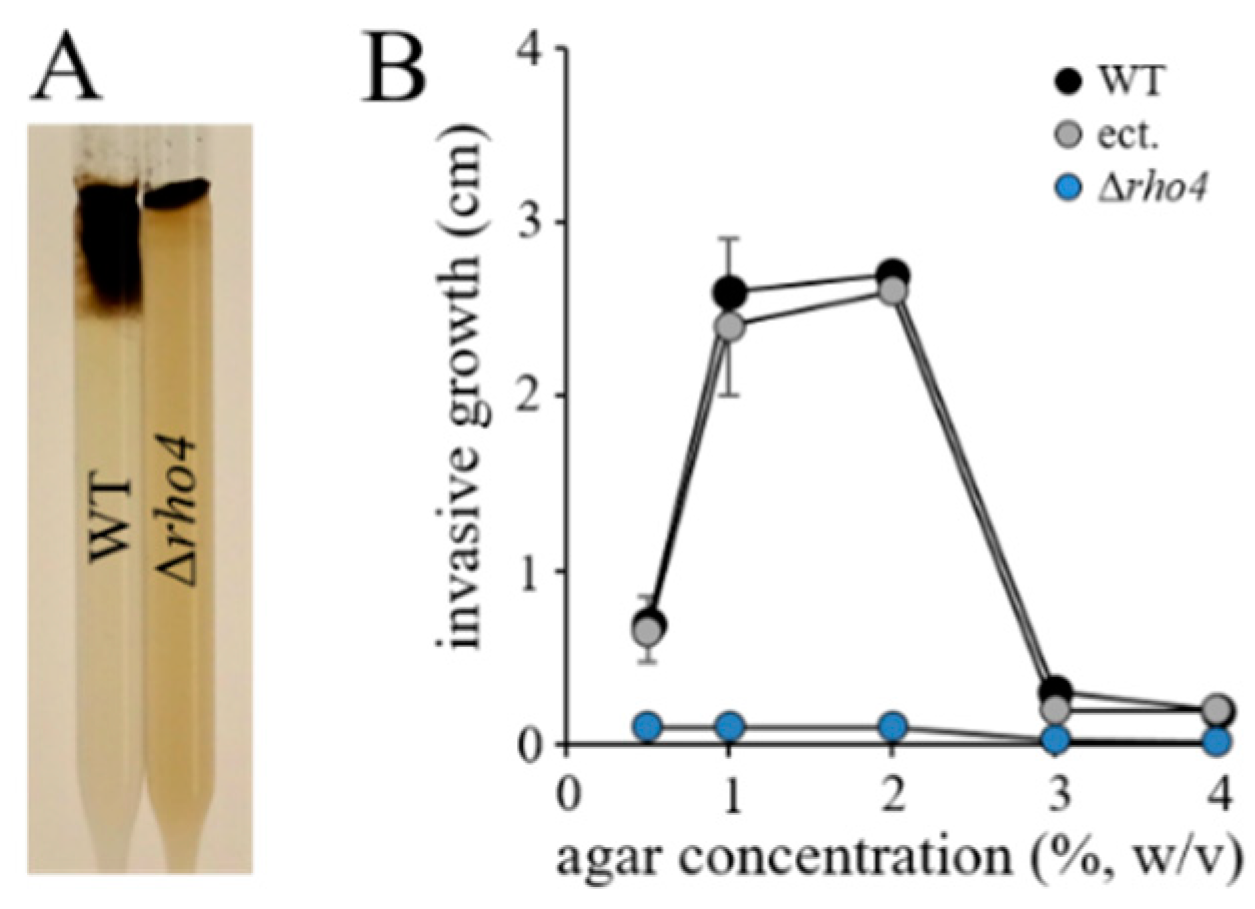
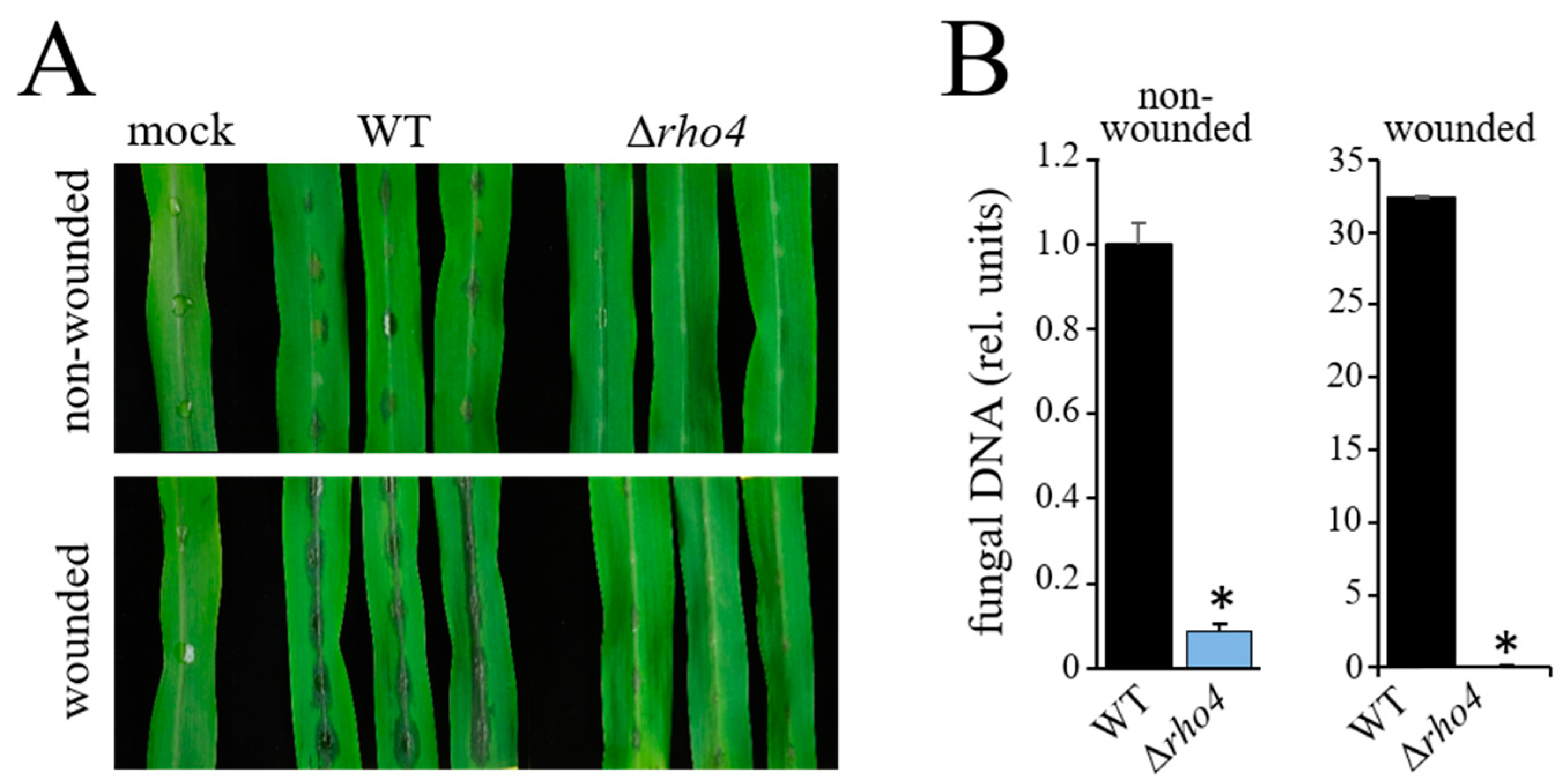
3.3. RHO4 Is Required for Invasive Growth, Infection Structure Differentiation, and Full Virulence
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cosio, E.G.; Feger, M.; Miller, C.J.; Antelo, L.; Ebel, J. High affinity binding of fungal β-glucan elicitors to cell membranes of species of the plant family Fabaceae. Planta 1996, 200, 92–99. [Google Scholar] [CrossRef]
- Mithöfer, A.; Fliegmann, J.; Neuhaus-Uri, G.; Schwarz, H.; Ebel, J. The hepta-β-glucoside elicitor-binding proteins from legumes represent a putative receptor family. Biol. Chem. 2000, 381, 705–713. [Google Scholar] [CrossRef]
- Böhm, H.; Albert, I.; Fan, L.; Reinhard, A.; Nürnberger, T. Immune receptor complexes at the plant cell surface. Curr. Opin. Plant Biol. 2014, 20, 47–54. [Google Scholar] [CrossRef]
- Fesel, P.H.; Zuccaro, A. β-glucan: Crucial component of the fungal cell wall and elusive MAMP in plants. Fungal Genet. Biol. 2016, 90, 53–60. [Google Scholar] [CrossRef]
- Oliveira-Garcia, E.; Deising, H.B. Infection structure-specific expression of β-1,3-glucan synthase is essential for pathogenicity of Colletotrichum graminicola and evasion of β-glucan-triggered immunity. Plant Cell 2013, 25, 2356–2378. [Google Scholar] [CrossRef]
- de Oliveira Silva, A.; Aliyeva-Schnorr, L.; Wirsel, S.G.R.; Deising, H.B. Fungal pathogenesis-related cell wall biogenesis, with emphasis on the maize anthracnose fungus Colletotrichum graminicola. Plants 2022, 11, 849. [Google Scholar] [CrossRef] [PubMed]
- Mendgen, K.; Deising, H. Infection structures of fungal plant pathogens—A cytological and physiological evaluation. New Phytol. 1993, 124, 193–213. [Google Scholar] [CrossRef]
- Mendgen, K.; Hahn, M.; Deising, H. Morphogenesis and mechanisms of penetration by plant pathogenic fungi. Annu. Rev. Phytopathol. 1996, 34, 367–386. [Google Scholar] [CrossRef]
- Frank, B. Ueber einige neue und weniger bekannte Pflanzenkrankheiten. Ber. Deutsch. Bot. Ges. 1883, 1, 29–34. [Google Scholar]
- Deising, H.B.; Werner, S.; Wernitz, M. The role of fungal appressoria in plant infection. Microbes Infect. 2000, 2, 1631–1641. [Google Scholar] [CrossRef]
- Wilson, R.A.; Talbot, N.J. Under pressure: Investigating the biology of plant infection by Magnaporthe oryzae. Nat. Rev. Microbiol. 2009, 7, 185–195. [Google Scholar] [CrossRef]
- Chethana, K.W.T.; Jayawardena, R.S.; Chen, Y.-J.; Konta, S.; Tibpromma, S.; Abeywickrama, P.D.; Gomdola, D.; Balasuriya, A.; Xu, J.; Lumyong, S.; et al. Diversity and function of appressoria. Pathogens 2021, 10, 746. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.J.; Ferrari, M.A.; Roach, D.H.; Money, N.P. Penetration of hard substances by a fungus employing enormous turgor pressures. Proc. Natl. Acad. Sci. USA 1991, 88, 11281–11284. [Google Scholar] [CrossRef] [PubMed]
- Bastmeyer, M.; Deising, H.B.; Bechinger, C. Force exertion in fungal infection. Ann. Rev. Biophys. Biomol. Struct. 2002, 31, 321–341. [Google Scholar] [CrossRef] [PubMed]
- Bechinger, C.; Giebel, K.-F.; Schnell, M.; Leiderer, P.; Deising, H.B.; Bastmeyer, M. Optical measurements of invasive forces exerted by appressoria of a plant pathogenic fungus. Science 1999, 285, 1896–1899. [Google Scholar] [CrossRef] [PubMed]
- Oliveira-Garcia, E.; Deising, H.B. Attenuation of PAMP-triggered immunity in maize requires down-regulation of the key β-1,6-glucan synthesis genes KRE5 and KRE6 in biotrophic hyphae of Colletotrichum graminicola. Plant J. 2016, 87, 355–375. [Google Scholar] [CrossRef] [PubMed]
- Bücherl, C.A.; Jarsch, I.K.; Schudoma, C.; Segonzac, C.; Mbengue, M.; Robatzek, S.; MacLean, D.; Ott, T.; Cyril Zipfel, C. Plant immune and growth receptors share common signalling components but localise to distinct plasma membrane nanodomains. eLife 2017, 6, e25114. [Google Scholar] [CrossRef]
- Gulli, M.-P.; Peter, M. Temporal and spatial regulation of Rho-type guanine-nucleotide exchange factors: The yeast perspective. Genes Devel. 2001, 15, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Dautt-Castro, M.; Rosendo-Vargas, M.; Casas-Flores, S. The small GTPases in fungal signaling conservation and function. Cells 2021, 10, 1039. [Google Scholar] [CrossRef] [PubMed]
- Cabib, E.; Drgonová, J.; Drgon, T. Role of small G proteins in yeast cell polarization and wall biosynthesis. Annu. Rev. Biochem. 1998, 67, 307–333. [Google Scholar] [CrossRef] [PubMed]
- Levin, D.E. Regulation of cell wall biogenesis in Saccharomyces cerevisiae: The cell wall integrity signaling pathway. Genetics 2011, 189, 1145–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, P.; Rincón, S.A. Rho GTPases: Regulation of cell polarity and growth in yeasts. Biochem. J. 2010, 426, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Qadota, H.; Python, C.P.; Inoue, S.B.; Arisawa, M.; Anraku, Y.; Zhang, Y.; Watanabe, T.; Levin, D.E.; Ohya, Y. Identification of a yeast Rho1p GTPase as a regulatory subunit of 1,3-β-glucan synthase. Science 1996, 272, 279–281. [Google Scholar] [CrossRef] [PubMed]
- Beauvais, A.; Bruneau, J.M.; Mol, P.C.; Buitrago, M.J.; Legrand, R.; Latgé, J.P. Glucan synthase complex of Aspergillus fumigatus. J. Bacteriol. 2001, 183, 2273–2279. [Google Scholar] [CrossRef]
- Zhang, X.; Jia, X.; Tian, S.; Zhang, C.; Lu, Z.; Chen, Y.; Chen, F.; Li, Z.; Su, X.; Han, X.; et al. Role of the small GTPase Rho1 in cell wall integrity, stress response, and pathogenesis of Aspergillus fumigatus. Fungal Genet. Biol. 2018, 120, 30–41. [Google Scholar] [CrossRef]
- Smith, S.E.; Csank, C.; Reyes, G.; Ghannoum, M.A.; Berlin, V. Candida albicans RHO1 is required for cell viability in vitro and in vivo. FEMS Yeast Res. 2002, 2, 103–111. [Google Scholar] [CrossRef]
- Wendland, J.; Philippsen, P. Cell polarity and hyphal morphogenesis are controlled by multiple rho-protein modules in the filamentous ascomycete Ashbya gossypii. Genetics 2001, 157, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, Y.; Wang, J.; Zhai, Z.; Zhang, L.; Zheng, W.; Zheng, W.; Yu, W.; Zhou, J.; Lu, G.; et al. Functional characterization of Rho family small GTPases in Fusarium graminearum. Fungal Genet. Biol. 2013, 61, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Werner, S.; Sugui, J.A.; Steinberg, G.; Deising, H.B. A chitin synthase with a myosin-like motor domain is essential for hyphal growth, appressorium differentiation and pathogenicity of the maize anthracnose fungus Colletotrichum graminicola. Mol. Plant-Microbe Interact. 2007, 20, 1555–1567. [Google Scholar] [CrossRef]
- Weihmann, F.; Eisermann, I.; Becher, R.; Krijger, J.-J.; Hübner, K.; Deising, H.B.; Wirsel, S.G.R. Correspondence between symptom development of Colletotrichum graminicola and fungal biomass, quantified by a newly developed qPCR assay, depends on the maize variety. BMC Microbiol. 2016, 16, 94. [Google Scholar] [CrossRef]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F.; et al. MycoCosm portal: Gearing up for 1000 fungal genomes. Nucleic Acids Res. 2014, 42, D699–D704. [Google Scholar] [CrossRef] [PubMed]
- Stenmark, H.; Olkkonen, V.M. The Rab GTPase family. Genome Biol. 2001, 2, reviews3007.1. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Luo, A.; Qiao, H.; Zhang, Y.; Shi, W.; Ho, S.Y.W.; Xu, W.; Zhang, A.; Zhu, C. Performance of criteria for selecting evolutionary models in phylogenetics: A comprehensive study based on simulated datasets. BMC Evol. Biol. 2010, 10, 242. [Google Scholar] [CrossRef]
- Yang, Z. PAML: A program package for phylogenetic analysis by maximum likelihood. Comp. Appl. BioSci. 1997, 13, 555–556. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview version 2: A multiple sequence alignment and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef]
- Malonek, S.; Rojas, M.C.; Hedden, P.; Gaskin, P.; Hopkins, P.; Tudzynski, B. The NADPH-cytochrome P450 reductase gene from Gibberella fujikuroi is essential for gibberellin biosynthesis. J. Biol. Chem. 2004, 279, 25075–25084. [Google Scholar] [CrossRef]
- Yu, J.H.; Hamari, Z.; Han, K.H.; Seo, J.A.; Reyes-Domínguez, Y.; Scazzocchio, C. Double-joint PCR: A PCR-based molecular tool for gene manipulations in filamentous fungi. Fungal Genet. Biol. 2004, 41, 973–981. [Google Scholar] [CrossRef]
- Döbbeling, U.; Böni, R.; Häffner, A.; Dummer, R.; Burg, G. Method for simultaneous RNA and DNA isolation from biopsy material, culture cells, plants and bacteria. Biotechniques 1997, 22, 88–90. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, R.J.; Thon, M.R.; Hacquard, S.; Amyotte, S.G.; Kleemann, J.; Torres, M.F.; Damm, U.; Buiate, E.A.; Epstein, L.; Alkan, N.; et al. Lifestyle transitions in plant pathogenic Colletotrichum fungi deciphered by genome and transcriptome analyses. Nat. Genet. 2012, 44, 1060–1065. [Google Scholar] [CrossRef]
- Oliveira-Garcia, E.; Deising, H.B. The glycosylphosphatidylinositol anchor biosynthesis genes GPI12, GAA1 and GPI8 are essential for cell wall integrity and pathogenicity of the maize anthracnose fungus Colletotrichum graminicola. Mol. Plant-Microbe Interact. 2016, 29, 889–901. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, S.P.; Patterson, T.F.; Lopez-Ribot, J.L. In vitro activity of caspofungin (MK-0991) against Candida albicans clinical isolates displaying different mechanisms of azole resistance. J. Clin. Microbiol. 2002, 40, 2228–2230. [Google Scholar] [CrossRef]
- Serfling, A.; Wohlrab, J.; Deising, H.B. Treatment of a clinically relevant plant pathogenic fungus with an agricultural azole causes cross-resistance to medical azoles and potentiates caspofungin efficacy. Antimicrob. Agents Chemother. 2007, 51, 3672–3676. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.A.; Gow, N.A.R.; Munro, C.A. Elevated chitin content reduces the susceptibility of Candida species to caspofungin. Antimicrob. Agents Chemother. 2013, 57, 146–154. [Google Scholar] [CrossRef]
- Walker, L.A.; Lee, K.K.; Munro, C.A.; Gow, N.A.R. Caspofungin treatment of Aspergillus fumigatus results in ChsG-dependent upregulation of chitin synthesis and the formation of chitin-rich microcolonies. Antimicrob. Agents Chemother. 2015, 59, 5932–5941. [Google Scholar] [CrossRef]
- Nezhad, A.S.; Geitmann, A. The cellular mechanics of an invasive lifestyle. J. Exp. Bot. Vol. 2013, 64, 4709–4728. [Google Scholar] [CrossRef]
- Groth, A.; Schunke, C.; Reschka, E.J.; Pöggeler, S.; Nordzieke, D.E. Tracking fungal growth: Establishment of Arp1 as a marker for polarity establishment and active hyphal growth in filamentous ascomycetes. J. Fungi 2021, 7, 580. [Google Scholar] [CrossRef]
- Arellano, M.; Duran, A.; Perez, R. Rho1 GTPase activates the (1,3)-β-D-glucan synthase and is involved in Schizosaccharomyces pombe morphogenesis. EMBO J. 1996, 15, 4584–4591. [Google Scholar] [CrossRef]
- Ozaki, K.; Tanaka, K.; Imamura, H.; Hihara, T.; Kameyama, T.; Nonaka, H.; Hirano, H.; Matsuura, Y.; Takai, Y. Rom1p and Rom2p are GDP/GTP exchange proteins (GEPs) for the Rho1p small GTP binding protein in Saccharomyces cerevisiae. EMBO J. 1996, 15, 2196–2207. [Google Scholar] [CrossRef]
- Richthammer, C.; Enseleit, M.; Sanchez-Leon, E.; März, S.; Heilig, Y.; Riquelme, M.; Seiler, S. RHO1 and RHO2 share partially overlapping functions in the regulation of cell wall integrity and hyphal polarity in Neurospora crassa. Mol. Microbiol. 2012, 85, 716–733. [Google Scholar] [CrossRef]
- Guest, G.M.; Lin, X.; Momany, M. Aspergillus nidulans RhoA is involved in polar growth, branching, and cell wall synthesis. Fungal Genet. Biol. 2004, 41, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.J.; Arentshorst, M.; Roos, E.D.; van den Hondel, C.A.M.J.J.; Meyer, V.; Ram, A.F.J. Functional characterization of Rho GTPases in Aspergillus niger uncovers conserved and diverged roles of Rho proteins within filamentous fungi. Mol. Microbiol. 2011, 79, 1151–1167. [Google Scholar] [CrossRef]
- Dichtl, K.; Helmschrott, C.; Dirr, F.; Wagener, J. Deciphering cell wall integrity signalling in Aspergillus fumigatus: Identification and functional characterization of cell wall stress sensors and relevant Rho GTPases. Mol. Microbiol. 2012, 83, 506–519. [Google Scholar] [CrossRef]
- Harris, S.D. Cdc42/Rho GTPases in fungi: Variations on a common theme. Mol. Microbiol. 2011, 79, 1123–1127. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.; Oliveira-Garcia, E.; Deising, H.B.; Peiter, E. A modular plasmid system for protein co-localization and bimolecular fluorescence complementation in filamentous fungi. Curr. Genet. 2014, 60, 343–350. [Google Scholar] [CrossRef]
- Rasmussen, C.G.; Glass, N.L. A Rho-type GTPase, rho-4, is required for septation in Neurospora crassa. Eukaryot. Cell 2005, 4, 1913–1925. [Google Scholar] [CrossRef]
- Si, H.; Justa-Schuch, D.; Seiler, S.; Harris, S.D. Regulation of septum formation by the Bud3-Rho4-GTPase module in Aspergillus nidulans. Genetics 2010, 185, 165–176. [Google Scholar] [CrossRef]
- Santos, B.; Gutiérrez, J.; Calonge, T.M.; Pérez, P. Novel Rho GTPase involved in cytokinesis and cell wall integrity in the fission yeast Schizosaccharomyces pombe. Eukaryot. Cell 2003, 2, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Chen, J.; Liu, W.; Zheng, S.; Zhou, J.; Lu, G.; Wang, Z. A Rho3 homolog is essential for appressorium development and pathogenicity of Magnaporthe grisea. Eukaryot. Cell 2007, 6, 2240–2250. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.S.; Zheng, W.; Zheng, S.Q.; Zhang, D.M.; Sang, W.J.; Chen, X.; Li, G.P.; Lu, G.D.; Wang, Z.H. Rac1 is required for pathogenicity and Chm1-dependent conidiogenesis in rice fungal pathogen Magnaporthe grisea. PLoS Pathog. 2008, 4, e1000202. [Google Scholar] [CrossRef]
- Zheng, W.; Zhao, Z.Y.; Chen, J.S.; Liu, W.D.; Ke, H.Y.; Zhou, J.; Lu, G.D.; Darvill, A.G.; Albersheim, P.; Wu, S.C.; et al. A Cdc42 ortholog is required for penetration and virulence of Magnaporthe grisea. Fungal Genet. Biol. 2009, 46, 450–460. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira-Garcia, E.; Aliyeva-Schnorr, L.; De Oliveira Silva, A.; El Din Ghanem, S.; Thor, K.; Peiter, E.; Deising, H.B. The Small Ras Superfamily GTPase Rho4 of the Maize Anthracnose Fungus Colletotrichum graminicola Is Required for β-1,3-glucan Synthesis, Cell Wall Integrity, and Full Virulence. J. Fungi 2022, 8, 997. https://doi.org/10.3390/jof8100997
Oliveira-Garcia E, Aliyeva-Schnorr L, De Oliveira Silva A, El Din Ghanem S, Thor K, Peiter E, Deising HB. The Small Ras Superfamily GTPase Rho4 of the Maize Anthracnose Fungus Colletotrichum graminicola Is Required for β-1,3-glucan Synthesis, Cell Wall Integrity, and Full Virulence. Journal of Fungi. 2022; 8(10):997. https://doi.org/10.3390/jof8100997
Chicago/Turabian StyleOliveira-Garcia, Ely, Lala Aliyeva-Schnorr, Alan De Oliveira Silva, Seif El Din Ghanem, Kathrin Thor, Edgar Peiter, and Holger B. Deising. 2022. "The Small Ras Superfamily GTPase Rho4 of the Maize Anthracnose Fungus Colletotrichum graminicola Is Required for β-1,3-glucan Synthesis, Cell Wall Integrity, and Full Virulence" Journal of Fungi 8, no. 10: 997. https://doi.org/10.3390/jof8100997
APA StyleOliveira-Garcia, E., Aliyeva-Schnorr, L., De Oliveira Silva, A., El Din Ghanem, S., Thor, K., Peiter, E., & Deising, H. B. (2022). The Small Ras Superfamily GTPase Rho4 of the Maize Anthracnose Fungus Colletotrichum graminicola Is Required for β-1,3-glucan Synthesis, Cell Wall Integrity, and Full Virulence. Journal of Fungi, 8(10), 997. https://doi.org/10.3390/jof8100997