
Multiple F-Box Proteins Collectively Regulate Cell Development and Pathogenesis in the Human Pathogen Cryptococcus neoformans



Abstract
:1. Introduction
2. Results
2.1. Identification of F-Box Proteins in C. neoformans
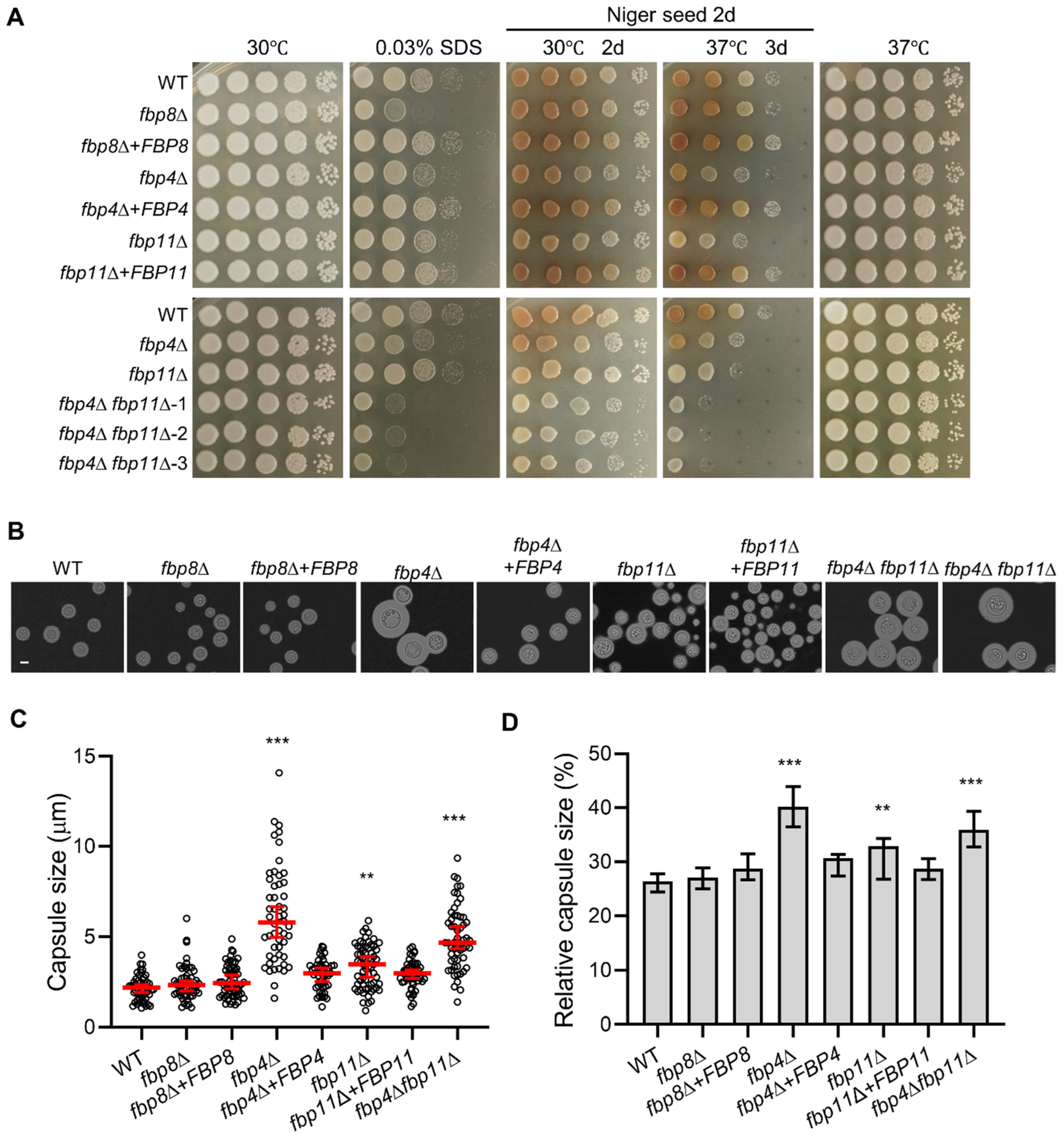
2.2. Phenotypes of F-Box Protein Encoding Genes Deletion Mutants under In Vitro Conditions
2.3. F-Box Proteins Are Important for Cell Membrane Integrity and Virulence Factor Production
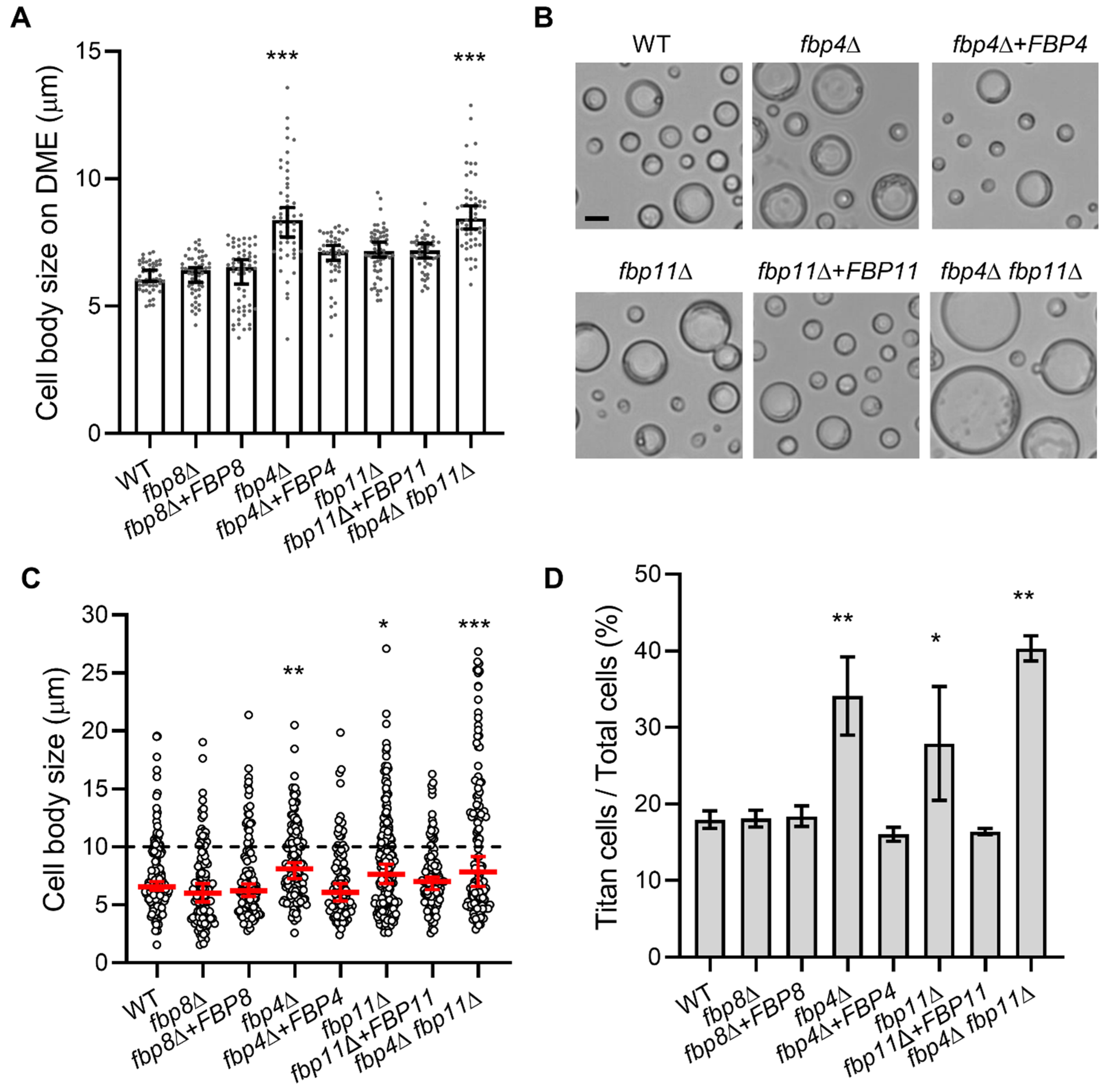
2.4. Fbp4 and Fbp11 Negatively Regulate Cell Size
2.5. Fbp8 and Fbp4 Are Important for Fluconazole Resistance
2.6. Fbp4 Is Essential for Sexual Reproduction
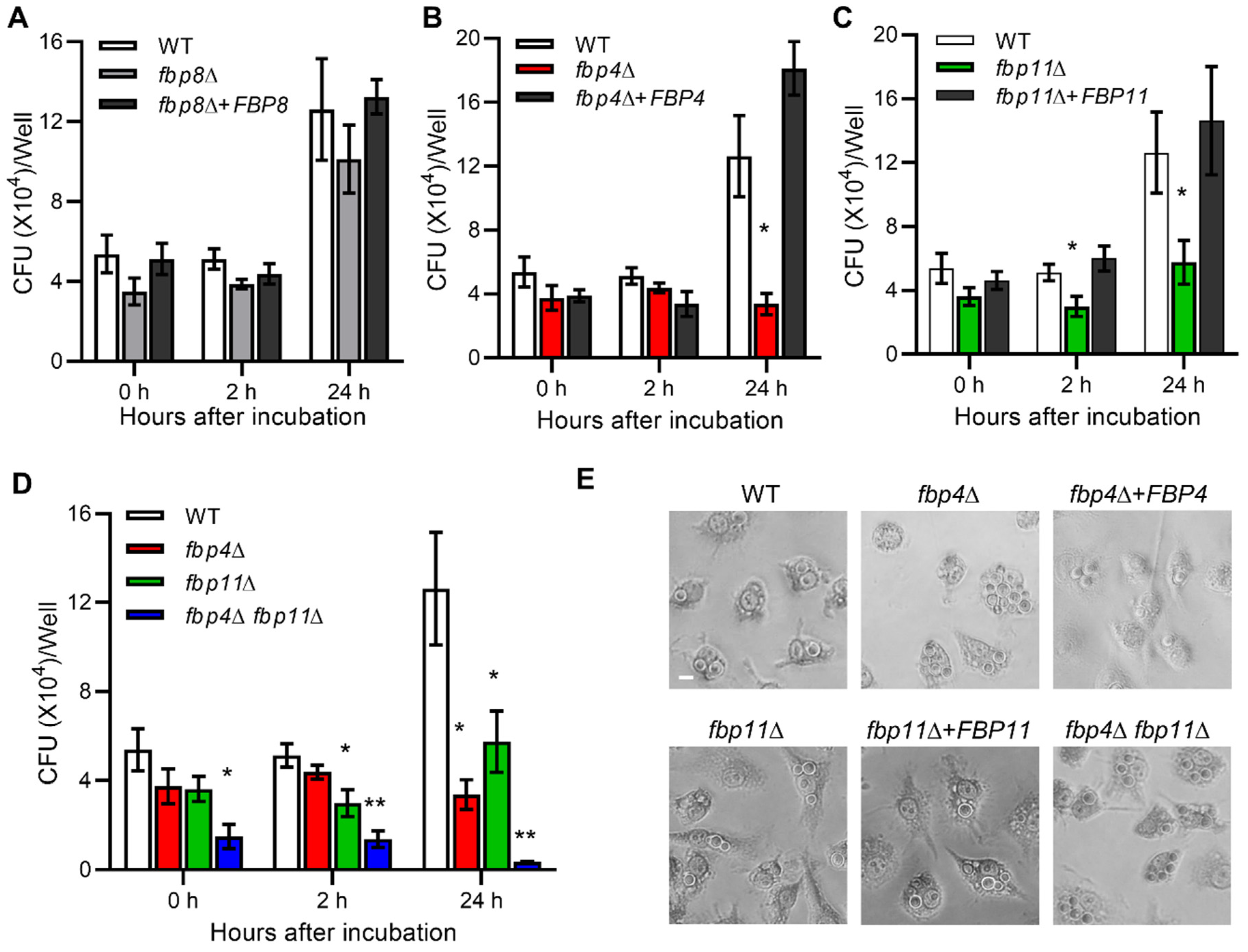
2.7. F-Box Proteins Are Involved in Cryptococcus-Macrophage Interaction
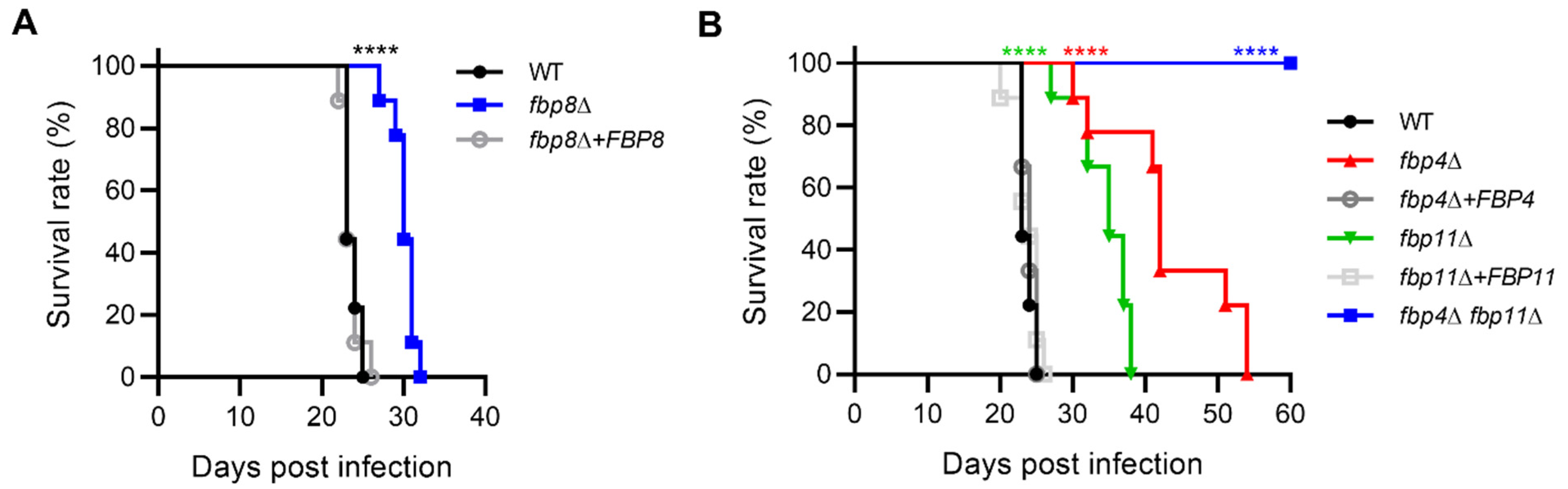
2.8. Roles of F-Box Proteins in Fungal Virulence
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and multi-national prevalence of fungal diseases-estimate precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G. Cryptococcal infections in non-HIV-infected patients. Trans. Am. Clin. Climatol. Assoc. 2013, 124, 61–79. [Google Scholar] [PubMed]
- Beardsley, J.; Sorrell, T.C.; Chen, S.C. Central nervous system cryptococcal infections in Non-HIV infected patients. J. Fungi 2019, 5, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D.R. Global burden of disease of HIV-associated cryptococcal meningitis: An updated analysis. Lancet Infect. Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef] [Green Version]
- Velagapudi, R.; Hsueh, Y.P.; Geunes-Boyer, S.; Wright, J.R.; Heitman, J. Spores as infectious propagules of Cryptococcus neoformans. Infect. Immun. 2009, 77, 4345–4355. [Google Scholar] [CrossRef] [Green Version]
- Kronstad, J.; Jung, W.H.; Hu, G. Beyond the big three: Systematic analysis of virulence factors in Cryptococcus neoformans. Cell Host. Microbe 2008, 4, 308–310. [Google Scholar] [CrossRef] [Green Version]
- Zaragoza, O. Basic principles of the virulence of Cryptococcus. Virulence 2019, 10, 490–501. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, K.; Heitman, J. Sex and virulence of human pathogenic fungi. Adv. Genet. 2007, 57, 143–173. [Google Scholar]
- Idnurm, A.; Bahn, Y.S.; Nielsen, K.; Lin, X.; Fraser, J.A.; Heitman, J. Deciphering the model pathogenic fungus Cryptococcus neoformans. Nat. Rev. Microbiol. 2005, 3, 753–764. [Google Scholar] [CrossRef]
- Altamirano, S.; Jackson, K.M.; Nielsen, K. The interplay of phenotype and genotype in Cryptococcus neoformans disease. Biosci. Rep. 2020, 40, BSR20190337. [Google Scholar] [CrossRef]
- Glickman, M.H.; Ciechanover, A. The ubiquitin-proteasome proteolytic pathway: Destruction for the sake of construction. Physiol. Rev. 2002, 82, 373–428. [Google Scholar] [CrossRef] [PubMed]
- Finley, D.; Ulrich, H.D.; Sommer, T.; Kaiser, P. The ubiquitin-proteasome system of Saccharomyces cerevisiae. Genetics 2012, 192, 319–360. [Google Scholar] [PubMed] [Green Version]
- Liu, T.B.; Wang, Y.; Stukes, S.; Chen, Q.; Casadevall, A.; Xue, C. The F-Box protein Fbp1 regulates sexual reproduction and virulence in Cryptococcus neoformans. Eukaryot. Cell 2011, 10, 791–802. [Google Scholar] [CrossRef] [Green Version]
- Cao, C.; Xue, C. More than tust cleaning: Ubiquitin-mediated proteolysis in fungal pathogenesis. Front. Cell. Infect. Microbiol. 2021, 11, 774613. [Google Scholar] [CrossRef]
- Atir-Lande, A.; Gildor, T.; Kornitzer, D. Role for the SCFCDC4 ubiquitin ligase in Candida albicans morphogenesis. Mol. Biol. Cell 2005, 16, 2772–2785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shieh, J.C.; White, A.; Cheng, Y.C.; Rosamond, J. Identification and functional characterization of Candida albicans CDC4. J. Biomed. Sci. 2005, 12, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Butler, D.K.; All, O.; Goffena, J.; Loveless, T.; Wilson, T.; Toenjes, K.A. The GRR1 gene of Candida albicans is involved in the negative control of pseudohyphal morphogenesis. Fungal Genet. Biol. 2006, 43, 573–582. [Google Scholar] [CrossRef]
- Li, W.J.; Wang, Y.M.; Zheng, X.D.; Shi, Q.M.; Zhang, T.T.; Bai, C.; Li, D.; Sang, J.L.; Wang, Y. The F-box protein Grr1 regulates the stability of Ccn1, Cln3 and Hof1 and cell morphogenesis in Candida albicans. Mol. Microbiol. 2006, 62, 212–226. [Google Scholar] [CrossRef]
- Jonkers, W.; van Kan, J.; Tijm, P.; Lee, Y.-W.; Tudzynski, P.; Rep, M.; Michielse, C.B. The FRP1 F-box gene has different functions in sexuality, pathogenicity and metabolism in three fungal pathogens. Mol. Plant Pathol. 2011, 12, 548–563. [Google Scholar] [CrossRef]
- Leach, M.D.; Stead, D.A.; Argo, E.; MacCallum, D.M.; Brown, A.J. Molecular and proteomic analyses highlight the importance of ubiquitination for the stress resistance, metabolic adaptation, morphogenetic regulation and virulence of Candida albicans. Mol. Microbiol. 2011, 79, 1574–1593. [Google Scholar] [CrossRef] [Green Version]
- Skowyra, D.; Craig, K.L.; Tyers, M.; Elledge, S.J.; Harper, J.W. F-box proteins are receptors that recruit phosphorylated substrates to the SCF ubiquitin-ligase complex. Cell 1997, 91, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Fan, C.L.; Han, L.T.; Guo, Y.B.; Liu, T.B. Role of F-box protein Cdc4 in fungal virulence and sexual reproduction of Cryptococcus neoformans. Front. Cell. Infect. Microbiol. 2021, 11, 806465. [Google Scholar] [CrossRef] [PubMed]
- Duyvesteijn, R.G.; van Wijk, R.; Boer, Y.; Rep, M.; Cornelissen, B.J.; Haring, M.A. Frp1 is a Fusarium oxysporum F-box protein required for pathogenicity on tomato. Mol. Microbiol. 2005, 57, 1051–1063. [Google Scholar] [CrossRef] [PubMed]
- Johnk, B.; Bayram, O.; Abelmann, A.; Heinekamp, T.; Mattern, D.J.; Brakhage, A.A.; Jacobsen, I.D.; Valerius, O.; Braus, G.H. SCF ubiquitin ligase F-box protein Fbx15 controls nuclear co-repressor localization, stress response and virulence of the human pathogen Aspergillus fumigatus. PLoS Pathog. 2016, 12, e1005899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frawley, D.; Bayram, O. Identification of SkpA-CulA-F-box E3 ligase complexes in pathogenic Aspergilli. Fungal Genet. Biol. 2020, 140, 103396. [Google Scholar] [CrossRef]
- Shi, H.B.; Chen, N.; Zhu, X.M.; Liang, S.; Li, L.; Wang, J.Y.; Lu, J.P.; Lin, F.C.; Liu, X.H. F-box proteins MoFwd1, MoCdc4 and MoFbx15 regulate development and pathogenicity in the rice blast fungus Magnaporthe oryzae. Environ. Microbiol. 2019, 21, 3027–3045. [Google Scholar] [CrossRef]
- Liu, T.B.; Xue, C. Fbp1-mediated ubiquitin-proteasome pathway controls Cryptococcus neoformans virulence by regulating fungal intracellular growth in macrophages. Infect. Immun. 2014, 82, 557–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masso-Silva, J.; Espinosa, V.; Liu, T.B.; Wang, Y.; Xue, C.; Rivera, A. The F-Box protein Fbp1 shapes the immunogenic potential of Cryptococcus neoformans. Mbio 2018, 9, e01828-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, K.; Masso-Silva, J.A.; Rivera, A.; Xue, C. A heat-killed Cryptococcus mutant strain induces host protection against multiple invasive mycoses in a murine vaccine model. Mbio 2019, 10, e02145-19. [Google Scholar] [CrossRef] [Green Version]
- Chun, C.D.; Madhani, H.D. Applying genetics and molecular biology to the study of the human pathogen Cryptococcus neoformans. Methods Enzym. 2010, 470, 797–831. [Google Scholar]
- Thomas, D.; Kuras, L.; Barbey, R.; Cherest, H.; Blaiseau, P.L.; Surdin-Kerjan, Y. Met30p, a yeast transcriptional inhibitor that responds to S-adenosylmethionine, is an essential protein with WD40 repeats. Mol. Cell. Biol. 1995, 15, 6526–6534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, R.B.; Woodword, A. Isolation and characterization of a pleiotropic glucose repression resistant mutant of Saccharomyces cerevisiae. Mol. Gen Genet. 1984, 193, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R.M.; Correll, C.C.; Kaplan, K.B.; Deshaies, R.J. A complex of Cdc4p, Skp1p, and Cdc53p/cullin catalyzes ubiquitination of the phosphorylated CDK inhibitor Sic1p. Cell 1997, 91, 221–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiederkehr, A.; Avaro, S.; Prescianotto-Baschong, C.; Haguenauer-Tsapis, R.; Riezman, H. The F-box protein Rcy1p is involved in endocytic membrane traffic and recycling out of an early endosome in Saccharomyces cerevisiae. J. Cell Biol. 2000, 149, 397–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samanta, M.P.; Liang, S. Predicting protein functions from redundancies in large-scale protein interaction networks. Proc. Natl. Acad. Sci. USA 2003, 100, 12579–12583. [Google Scholar] [CrossRef] [Green Version]
- Escusa, S.; Camblong, J.; Galan, J.M.; Pinson, B.; Daignan-Fornier, B. Proteasome- and SCF-dependent degradation of yeast adenine deaminase upon transition from proliferation to quiescence requires a new F-box protein named Saf1p. Mol. Microbiol. 2006, 60, 1014–1025. [Google Scholar] [CrossRef]
- Kozel, T.R.; Gotschlich, E.C. The capsule of Cryptococcus neoformans passively inhibits phagocytosis of the yeast by macrophages. J. Immunol. 1982, 129, 1675–1680. [Google Scholar]
- Wang, Y.; Aisen, P.; Casadevall, A. Cryptococcus neoformans melanin and virulence: Mechanism of action. Infect. Immun. 1995, 63, 3131–3136. [Google Scholar] [CrossRef] [Green Version]
- Okagaki, L.H.; Strain, A.K.; Nielsen, J.N.; Charlier, C.; Baltes, N.J.; Chretien, F.; Heitman, J.; Dromer, F.; Nielsen, K. Cryptococcal cell morphology affects host cell interactions and pathogenicity. PLoS Pathog. 2010, 6, e1000953. [Google Scholar] [CrossRef]
- Zaragoza, O.; Garcia-Rodas, R.; Nosanchuk, J.D.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L.; Casadevall, A. Fungal cell gigantism during mammalian infection. PLoS Pathog. 2010, 6, e1000945. [Google Scholar] [CrossRef]
- Garcia-Rodas, R.; de Oliveira, H.C.; Trevijano-Contador, N.; Zaragoza, O. Cryptococcal titan cells: When yeast cells are all grown up. Curr. Top Microbiol. Immunol. 2019, 422, 101–120. [Google Scholar] [PubMed]
- Cao, C.; Wang, K.; Wang, Y.; Liu, T.B.; Rivera, A.; Xue, C. Ubiquitin proteolysis of a CDK-related kinase regulates titan cell formation and virulence in the fungal pathogen Cryptococcus neoformans. Nat. Commun. 2022, 13, 6397. [Google Scholar] [CrossRef] [PubMed]
- Hommel, B.; Mukaremera, L.; Cordero, R.J.B.; Coelho, C.; Desjardins, C.A.; Sturny-Leclere, A.; Janbon, G.; Perfect, J.R.; Fraser, J.A.; Casadevall, A.; et al. Titan cells formation in Cryptococcus neoformans is finely tuned by environmental conditions and modulated by positive and negative genetic regulators. PLoS Pathog. 2018, 14, e1006982. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.; Casadevall, A. Phagosome extrusion and host-cell survival after Cryptococcus neoformans phagocytosis by macrophages. Curr. Biol. 2006, 16, 2161–2165. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Croudace, J.E.; Lammas, D.A.; May, R.C. Expulsion of live pathogenic yeast by macrophages. Curr. Biol. 2006, 16, 2156–2160. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.S. Mechanisms of microbial traversal of the blood-brain barrier. Nat. Rev. Microbiol. 2008, 6, 625–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.B.; Xue, C. The ubiquitin-proteasome system and F-box proteins in pathogenic fungi. Mycobiology 2011, 39, 243–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenbroucke, K.; Robbens, S.; Vandepoele, K.; Inze, D.; Van de Peer, Y.; Van Breusegem, F. Hydrogen peroxide-induced gene expression across kingdoms: A comparative analysis. Mol. Biol. Evol. 2008, 25, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Ko, Y.J.; Yu, Y.M.; Kim, G.B.; Lee, G.W.; Maeng, P.J.; Kim, S.; Floyd, A.; Heitman, J.; Bahn, Y.S. Remodeling of global transcription patterns of Cryptococcus neoformans genes mediated by the stress-activated HOG signaling pathways. Eukaryot Cell 2009, 8, 1197–1217. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Watt, R.; Piper, P.W. Polyubiquitin gene expression contributes to oxidative stress resistance in respiratory yeast (Saccharomyces cerevisiae). Mol. Gen. Genet. 1994, 243, 358–362. [Google Scholar] [CrossRef]
- Fang, W.; Price, M.S.; Toffaletti, D.L.; Tenor, J.; Betancourt-Quiroz, M.; Price, J.L.; Pan, W.H.; Liao, W.Q.; Perfect, J.R. Pleiotropic effects of deubiquitinating enzyme Ubp5 on growth and pathogenesis of Cryptococcus neoformans. PLoS ONE 2012, 7, e38326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanamatsu, H.; Fujimura-Kamada, K.; Yamamoto, T.; Furuta, N.; Tanaka, K. Interaction of the phospholipid flippase Drs2p with the F-box protein Rcy1p plays an important role in early endosome to trans-Golgi network vesicle transport in yeast. J. Biochem. 2014, 155, 51–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spampinato, C.; Leonardi, D. Candida infections, causes, targets, and resistance mechanisms: Traditional and alternative antifungal agents. Biomed. Res. Int. 2013, 2013, 204237. [Google Scholar] [CrossRef]
- Khosravi Rad, K.; Falahati, M.; Roudbary, M.; Farahyar, S.; Nami, S. Overexpression of MDR-1 and CDR-2 genes in fluconazole resistance of Candida albicans isolated from patients with vulvovaginal candidiasis. Curr. Med. Mycol. 2016, 2, 24–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sionov, E.; Chang, Y.C.; Kwon-Chung, K.J. Azole heteroresistance in Cryptococcus neoformans: Emergence of resistant clones with chromosomal disomy in the mouse brain during fluconazole treatment. Antimicrob. Agents Chemother. 2013, 57, 5127–5130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, M.; Sionov, E.; Khanal Lamichhane, A.; Kwon-Chung, K.J.; Chang, Y.C. Roles of three Cryptococcus neoformans and Cryptococcus gattii efflux pump-coding genes in response to drug treatment. Antimicrob. Agents Chemother. 2018, 62, e01751-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenoir, G.; Williamson, P.; Puts, C.F.; Holthuis, J.C. Cdc50p plays a vital role in the ATPase reaction cycle of the putative aminophospholipid transporter Drs2p. J. Biol. Chem. 2009, 284, 17956–17967. [Google Scholar] [CrossRef] [Green Version]
- Furuta, N.; Fujimura-Kamada, K.; Saito, K.; Yamamoto, T.; Tanaka, K. Endocytic recycling in yeast is regulated by putative phospholipid translocases and the Ypt31p/32p-Rcy1p pathway. Mol. Biol. Cell 2007, 18, 295–312. [Google Scholar] [CrossRef]
- Hankins, H.M.; Sere, Y.Y.; Diab, N.S.; Menon, A.K.; Graham, T.R. Phosphatidylserine translocation at the yeast trans-Golgi network regulates protein sorting into exocytic vesicles. Mol. Biol. Cell 2015, 26, 4674–4685. [Google Scholar] [CrossRef]
- Huang, W.; Liao, G.; Baker, G.M.; Wang, Y.; Lau, R.; Paderu, P.; Perlin, D.S.; Xue, C. Lipid flippase subunit Cdc50 mediates drug resistance and virulence in Cryptococcus neoformans. Mbio 2016, 7, e00478-16. [Google Scholar] [CrossRef] [Green Version]
- Love, G.L.; Boyd, G.D.; Greer, D.L. Large Cryptococcus neoformans isolated from brain abscess. J. Clin. Microbiol. 1985, 22, 1068–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okagaki, L.H.; Wang, Y.; Ballou, E.R.; O’Meara, T.R.; Bahn, Y.S.; Alspaugh, J.A.; Xue, C.; Nielsen, K. Cryptococcal titan cell formation is regulated by G-protein signaling in response to multiple stimuli. Eukaryot. Cell 2011, 10, 1306–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartwell, L.H.; Mortimer, R.K.; Culotti, J.; Culotti, M. Genetic control of the cell division cycle in yeast: V. Genetic analysis of cdc mutants. Genetics 1973, 74, 267–286. [Google Scholar] [CrossRef] [PubMed]
- Goh, P.Y.; Surana, U. Cdc4, a protein required for the onset of S phase, serves an essential function during G(2)/M transition in Saccharomyces cerevisiae. Mol. Cell. Biol. 1999, 19, 5512–5522. [Google Scholar] [CrossRef] [Green Version]
- Altamirano, S.; Li, Z.; Fu, M.S.; Ding, M.; Fulton, S.R.; Yoder, J.M.; Tran, V.; Nielsen, K. The cyclin Cln1 controls polyploid titan cell formation following a stress-Induced G2 arrest in Cryptococcus. Mbio 2021, 12, e0250921. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, Y.; Upadhyay, S.; Xue, C.; Lin, X. Activation of meiotic genes mediates ploidy reduction during Cryptococcal infection. Curr. Biol. 2020, 30, 1387–1396 e1385. [Google Scholar] [CrossRef]
- Lin, X. Cryptococcus neoformans: Morphogenesis, infection, and evolution. Infect. Genet. Evol. 2009, 9, 401–416. [Google Scholar] [CrossRef]
- Simchen, G.; Hirschberg, J. Effects of the mitotic cell-cycle mutation cdc4 on yeast meiosis. Genetics 1977, 86, 57–72. [Google Scholar] [CrossRef]
- Zhu, Z.; Bani Ismail, M.; Shinohara, M.; Shinohara, A. SCF(Cdc4) ubiquitin ligase regulates synaptonemal complex formation during meiosis. Life Sci. Alliance 2021, 4, e202000933. [Google Scholar] [CrossRef]
- Bahn, Y.S.; Hicks, J.K.; Giles, S.S.; Cox, G.M.; Heitman, J. Adenylyl cyclase-associated protein Aca1 regulates virulence and differentiation of Cryptococcus neoformans via the cyclic AMP-protein kinase A cascade. Eukaryot. Cell 2004, 3, 1476–1491. [Google Scholar] [CrossRef] [Green Version]
- Xue, C.; Tada, Y.; Dong, X.; Heitman, J. The human fungal pathogen Cryptococcus can complete its sexual cycle during a pathogenic association with plants. Cell Host Microbe 2007, 1, 263–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Lin, X. Multiple applications of a transient CRISPR-Cas9 coupled with electroporation (TRACE) system in the Cryptococcus neoformans species complex. Genetics 2018, 208, 1357–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, G.M.; Mukherjee, J.; Cole, G.T.; Casadevall, A.; Perfect, J.R. Urease as a virulence factor in experimental cryptococcosis. Infect. Immun. 2000, 68, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Perfect, J.R.; Schell, W.A.; Rinaldi, M.G. Uncommon invasive fungal pathogens in the acquired immunodeficiency syndrome. J. Med. Vet. Mycol. 1993, 31, 175–179. [Google Scholar] [CrossRef]
- Nielsen, K.; Cox, G.M.; Wang, P.; Toffaletti, D.L.; Perfect, J.R.; Heitman, J. Sexual cycle of Cryptococcus neoformans var. grubii and virulence of congenic a and alpha isolates. Infect. Immun. 2003, 71, 4831–4841. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID | Homolog in S. cerevisiae | Deletion Location | Function | References |
|---|---|---|---|---|---|
| Fbp1 | CNAG_05280 | Grr1 | 15C11 | F-box and leucine-rich repeat protein GRR1 | [13,32] |
| Fbp2 | CNAG_00134 | - | 44B9 | hypothetical protein | |
| Fbp3 | CNAG_00416 | - | 21C12 | F-box/WD-repeat protein lin-23 | |
| Fbp4 | CNAG_00693 | Cdc4 | 34G11 | F-box and WD-40 domain-containing protein CDC4 | [22,33] |
| Fbp5 | CNAG_02349 | - | 16H8 | hypothetical protein | |
| Fbp6 | CNAG_03157 | - | KO | hypothetical protein | |
| Fbp7 | CNAG_03421 | - | KO | hypothetical protein | |
| Fbp8 | CNAG_04341 | Rcy1 | 31E5 | recyclin-1 | [34] |
| Fbp9 | CNAG_04462 | Hrt3 | 31F6 | F-box protein 9 | [35] |
| Fbp10 | CNAG_04606 | - | 31H11 | hypothetical protein | |
| Fbp11 | CNAG_05294 | Cdc4 | 33B3 | F-box and WD-40 domain-containing protein CDC4 | [22,33] |
| Fbp12 | CNAG_05450 | - | KO | hypothetical protein | |
| Fbp13 | CNAG_05454 | - | KO | hypothetical protein | |
| Fbp14 | CNAG_05773 | Met30 | - | F-box and WD-40 domain-containing protein MET30 | [31] |
| Fbp15 | CNAG_05874 | - | 37A5 | hypothetical protein | |
| Fbp16 | CNAG_06382 | - | 38C8 | beta-transducin repeat containing protein | |
| Fbp17 | CNAG_06722 | - | 43F9 | hypothetical protein | |
| Fbp18 | CNAG_07482 | Saf1 | 40C3 | SCF-associated factor 1 | [36] |
| Fbp19 | CNAG_07551 | - | KO | hypothetical protein | |
| Fbp20 | CNAG_07702 | - | 11F3 | hypothetical protein |
| Strains | MIC90 (µg/mL) | ||
|---|---|---|---|
| Fluconazole | Caspofungin | Amphotericin B | |
| WT | 8 | 16 | 1 |
| fbp8∆ | 2 | 16 | 1 |
| fbp8∆ + FBP8 | 8 | 16 | 1 |
| fbp4∆ | 2 | 16 | 1 |
| fbp4∆ + FBP4 | 8 | 16 | 1 |
| fbp11∆ | 8 | 16 | 1 |
| fbp11∆ + FBP11 | 8 | 16 | 1 |
| fbp4∆ fbp11∆ | 2 | 16 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, C.; Wang, Y.; Avina, S.L.; Walter, J.; Xue, C. Multiple F-Box Proteins Collectively Regulate Cell Development and Pathogenesis in the Human Pathogen Cryptococcus neoformans. J. Fungi 2022, 8, 1259. https://doi.org/10.3390/jof8121259
Cao C, Wang Y, Avina SL, Walter J, Xue C. Multiple F-Box Proteins Collectively Regulate Cell Development and Pathogenesis in the Human Pathogen Cryptococcus neoformans. Journal of Fungi. 2022; 8(12):1259. https://doi.org/10.3390/jof8121259
Chicago/Turabian StyleCao, Chengjun, Yina Wang, Samantha L. Avina, John Walter, and Chaoyang Xue. 2022. "Multiple F-Box Proteins Collectively Regulate Cell Development and Pathogenesis in the Human Pathogen Cryptococcus neoformans" Journal of Fungi 8, no. 12: 1259. https://doi.org/10.3390/jof8121259
APA StyleCao, C., Wang, Y., Avina, S. L., Walter, J., & Xue, C. (2022). Multiple F-Box Proteins Collectively Regulate Cell Development and Pathogenesis in the Human Pathogen Cryptococcus neoformans. Journal of Fungi, 8(12), 1259. https://doi.org/10.3390/jof8121259