
Taxonomy and Phylogeny of Novel and Extant Taxa in Pleosporales Associated with Mangifera indica from Yunnan, China (Series I)



 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analyses
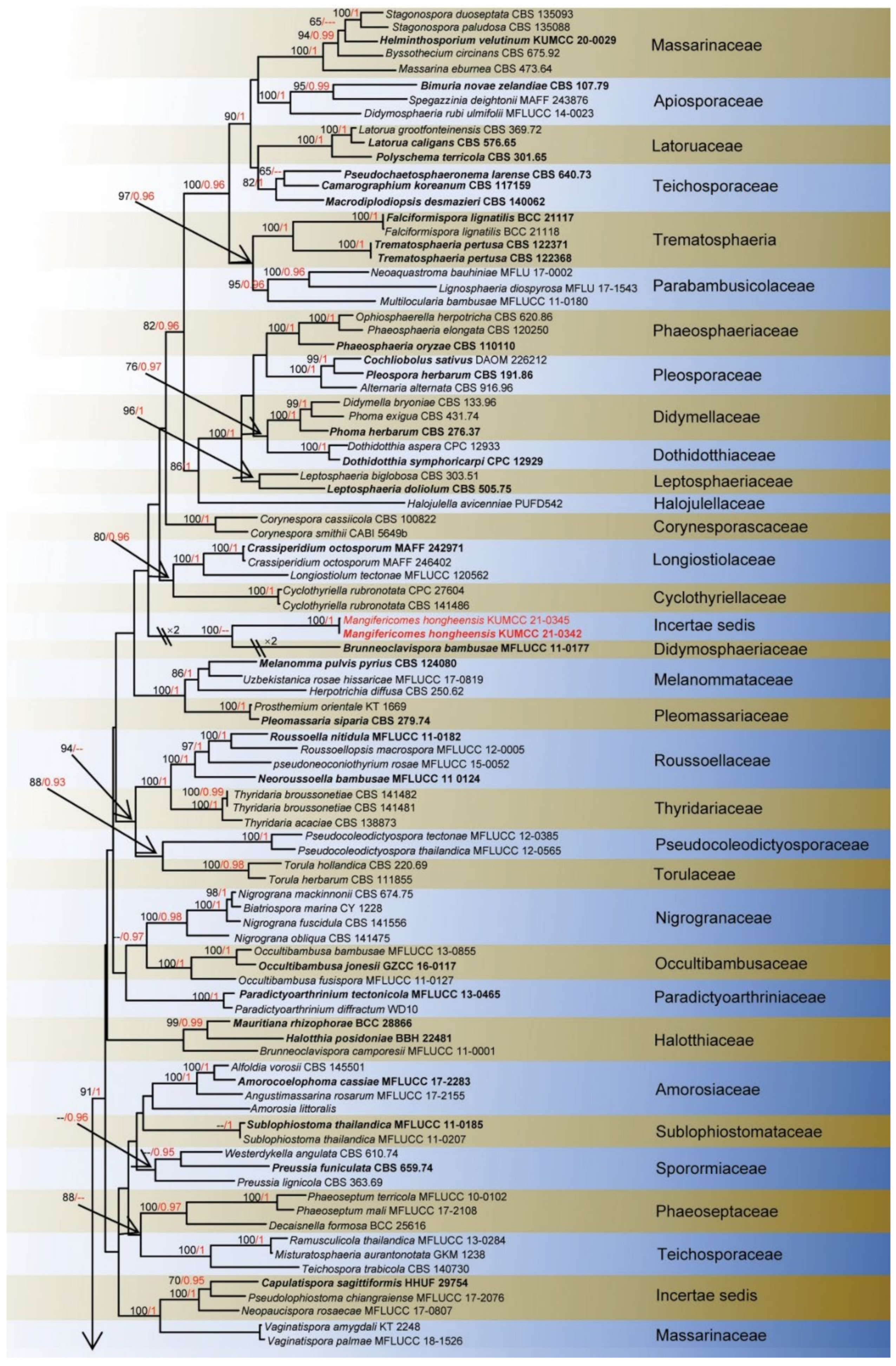
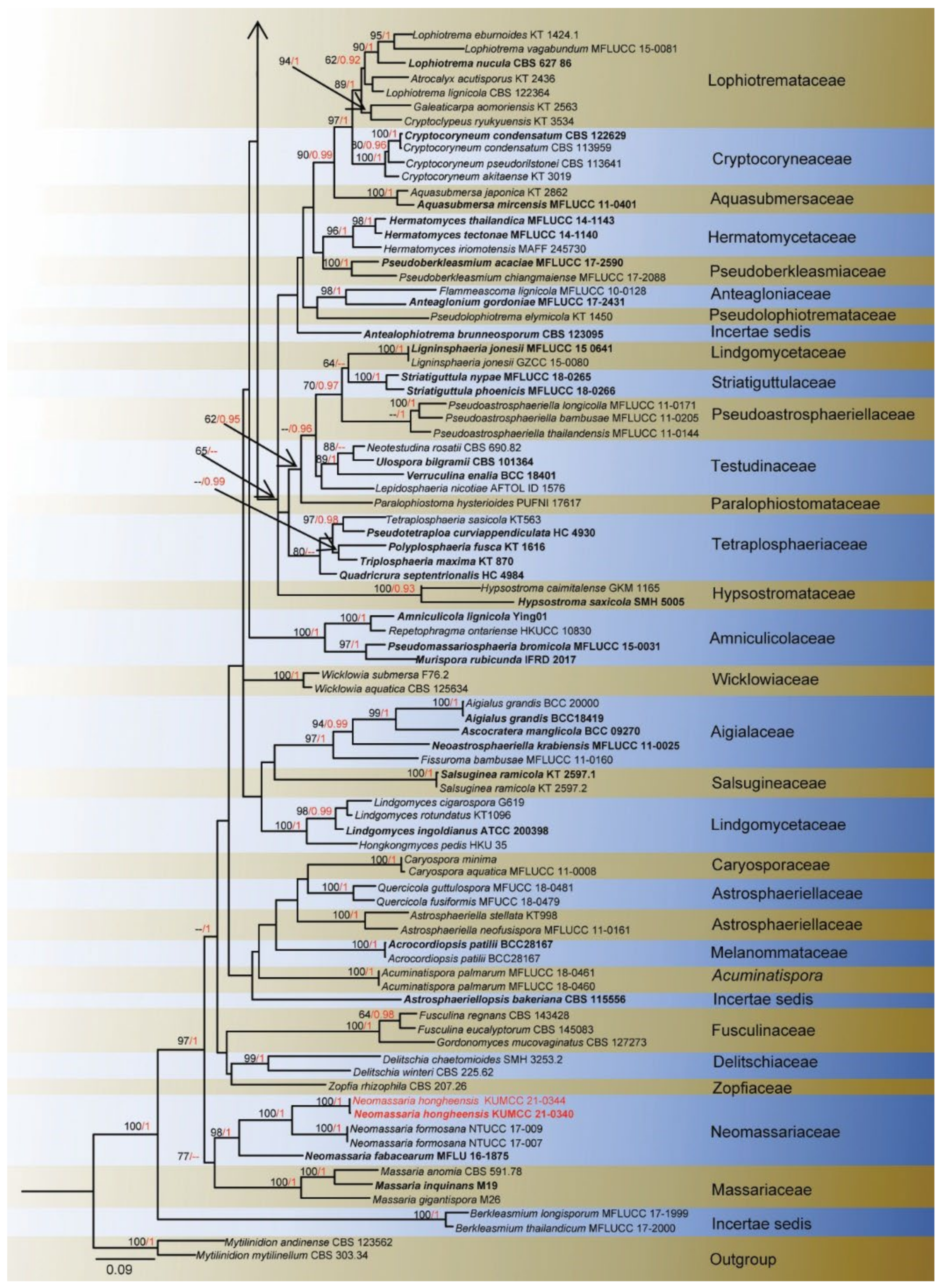
3. Results
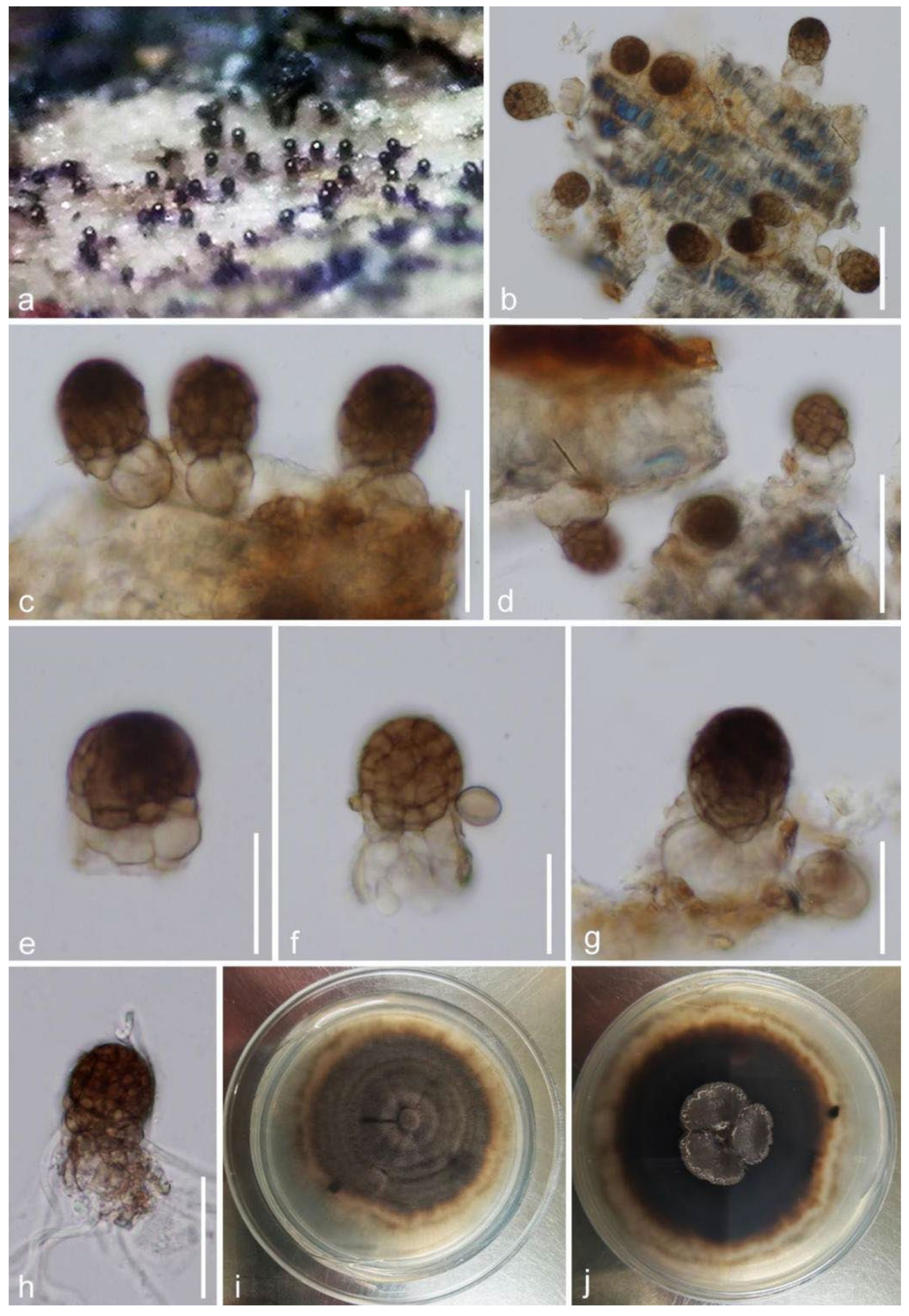
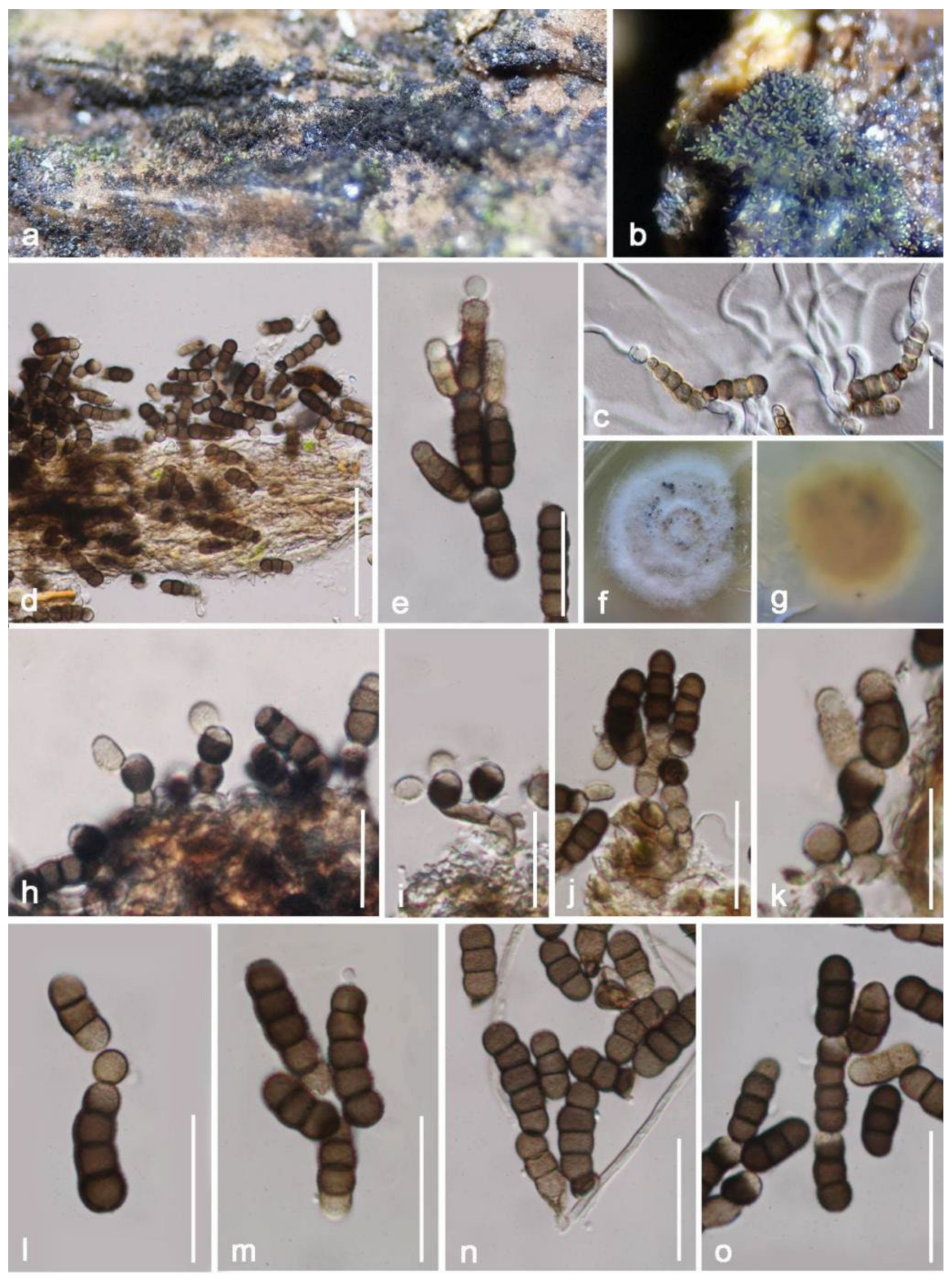
Taxonomy and Phylogenetic Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shah, K.A.; Patel, M.B.; Patel, R.J.; Parmar, P.K. Mangifera indica (mango). Pharmacogn. Rev. 2010, 4, 42–48. [Google Scholar] [CrossRef]
- Solís-Fuentes, J.A.; del Carmen Durán-de-Bazúa, M. Mango (Mangifera indica L.) seed and its fats. In Nuts and Seeds in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2011; pp. 741–748. [Google Scholar] [CrossRef]
- Muchiri, D.R.; Mahungu, S.M.; Gituanja, S.N. Studies on mango (Mangifera indica, L.) kernel fat of some Kenyan varieties in Meru. J. Am. Oil Chem. Soc. 2012, 89, 1567–1575. [Google Scholar] [CrossRef]
- Jahurul, M.H.A.; Zaidul, I.S.M.; Ghafoor, K.; Al-Juhaimi, F.Y.; Nyam, K.L.; Norulaini, N.A.N.; Sahena, F.; Omar, A.M. Mango (Mangifera indica L.) by-products and their valuable components: A review. Food Chem. 2015, 183, 173–180. [Google Scholar] [CrossRef]
- DeCandolle, A.P. Origin of Cultivated Plants; The International Science Series; Harvard University: London, UK, 1884; pp. 1–462. [Google Scholar] [CrossRef]
- Douthett, B.D.G. The mango: Asia’s king of fruits. Ethnobot. Leafl. 2011, 2000, 4. [Google Scholar]
- Gao, A.; Chen, Y.; Luo, R.; Huang, J.; Zhao, Z.; Wang, W.; Wang, Y.; Dang, Z. Development status of Chinese mango industry in 2018. Adv. Agric. Hortic. Entomol. 2019, 1, 21–60. [Google Scholar]
- Calatrava-Requena, J. Mango: Economics and international trade. In Mango International Encyclopedia; Calatrava-Requena, J., Ed.; Royal Court Affairs: Muscat, Oman, 2014; pp. 1–45. [Google Scholar]
- Lauricella, M.; Emanuele, S.; Calvaruso, G.; Giuliano, M.; D’Anneo, A. Multifaceted health benefits of Mangifera indica L. (Mango): The inestimable value of orchards recently planted in Sicilian rural areas. Nutrients 2017, 9, 525. [Google Scholar] [CrossRef] [PubMed]
- Diedhiou, P.M.; Mbaye, N.; Drame, A.; Samb, P.I. Alteration of post harvest diseases of mango Mangifera indica through production practices and climatic factors. Afr. J. Biotechnol. 2007, 6, 1087–1094. [Google Scholar] [CrossRef] [Green Version]
- Sandoval-Sánchez, M.; Nieto-Ángel, D.; Sandoval-Islas, J.S.; Téliz-Ortiz, D.; Orozco-Santos, M.; Silva-Rojas, H. Fungi associated to stem-end rot and dieback of mango (Mangifera indica L.). Agrociencia 2013, 47, 61–73. [Google Scholar]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA, 2021. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 25 January 2022).
- Doilom, M.; Dissanayake, A.J.; Wanasinghe, D.N.; Boonmee, S.; Liu, J.K.; Bhat, D.J.; Taylor, J.E.; Bahkali, A.H.; Eric, H.; Mckenzie, C.; et al. Microfungi on Tectona grandis (teak) in Northern Thailand. Fungal Divers. 2017, 82, 107–182. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; Mckenzie, E.; Bhat, D.J.; Karunarathna, S.C. Fungal diversity notes 840–928: Micro-fungi associated with pandanaceae. Fungal Divers. 2018, 93, 1–160. [Google Scholar] [CrossRef]
- Raza, M.; Zhang, Z.F.; Hyde, K.D.; Diao, Y.Z.; Cai, L. Culturable plant pathogenic fungi associated with sugarcane in southern China. Fungal Divers. 2019, 99, 1–104. [Google Scholar] [CrossRef]
- Mapook, A.; Hyde, K.D.; McKenzie, E.H.C.; Jones, E.B.G.; Bhat, D.J.; Jeewon, R.; Stadler, M.; Samarakoon, M.C.; Malaithong, M.; Tanunchai, B.; et al. Taxonomic and phylogenetic contributions to fungi associated with the invasive weed Chromolaena odorata (Siam weed). Fungal Divers. 2020, 101, 1–175. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; McKenzie, E.H.C.; Phillips, A.J.L.; Jones, E.B.G.; Bhat, D.J.; Stadler, M.; Bhunjun, C.S.; Wanasinghe, D.N.; Thongbai, B.; Camporesi, E.; et al. Microfungi associated with Clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 2020, 102, 1–203. [Google Scholar] [CrossRef]
- Hyde, K.D.; Lee, S.Y. Ecology of mangrove fungi and their role in nutrient cycling: What gaps occur in our knowledge? Hydrobiologia 1995, 295, 107–118. [Google Scholar] [CrossRef]
- Zhou, D.; Hyde, K.D. Host-specificity, hostexclusivity, and host-recurrence in saprobic fungi. Mycol. Res. 2001, 105, 1449–1457. [Google Scholar] [CrossRef]
- Roy, B.A. Patterns of association between crucifers and their flower-mimic pathogens: Host jumps are more common than coevolution or cospeciation. Evolution 2001, 55, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Senwanna, C.; Hongsanan, S.; Phookamsak, R.; Tibpromma, S.; Cheewangkoon, R.; Hyde, K.D. Muyocopron heveae sp. nov. and M. dipterocarpi appears to have host-jumped to rubber. Mycol Prog. 2019, 18, 741–752. [Google Scholar] [CrossRef]
- Luttrell, E.S. The ascostromatic ascomycetes. Mycologia 1995, 47, 511–532. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Lumbsch, H.T.; Liu, J.K.; Maharachchikumbura, S.S.; Ekanayaka, A.H.; Tian, Q.; Phookamsak, R. Outline of ascomycota: 2017. Fungal Divers. 2018, 88, 167–263. [Google Scholar] [CrossRef]
- Zhang, S.N.; Hyde, K.D.; Jones, E.B.G.; Cheewangkoon, R.; Liu, J.K. Acuminatispora palmarum gen. et. sp. nov. from mangrove habitats. Mycol. Prog. 2018, 17, 1173–1188. [Google Scholar] [CrossRef]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Lücking, R.; Pem, D.; Bhat, J.D.; et al. Refned families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
- Zhang, Y.; Schoch, C.L.; Fournier, J.; Crous, P.W.; De Gruyter, J.; Woudenberg, J.H.C.; Hirayama, K.; Tanaka, K.; Pointing, B.; Spatafora, J.W.; et al. Multi-locus phylogeny of Pleosporales: A taxonomic, ecological and evolutionary re-evaluation. Stud. Mycol. 2009, 64, 85–102. [Google Scholar] [CrossRef] [PubMed]
- De Gruyter, J.D.; Woudenberg, J.H.C.; Aveskamp, M.M.; Verkley, G.J.M.; Groenewald, J.Z.; Crous, P.W. Systematic reappraisal of species in Phoma section Paraphoma, Pyrenochaeta and Pleurophoma. Mycologia 2010, 102, 1066–1081. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Koko, T.W.; Hyde, K.D. Towards a monograph of Dothideomycetes: Studies on Diademaceae. Cryptogam. Mycol. 2011, 32, 115–126. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Voglmayr, H. Hidden diversity in Thyridaria and a new circumscription of the Thyridariaceae. Stud. Mycol. 2016, 85, 35–64. [Google Scholar] [CrossRef] [Green Version]
- Ariyawansa, H.A.; Jaklitsch, W.M.; Voglmayr, H. Additions to Taiwan fungal flora 1: Neomassariaceae fam. nov. Cryptogam. Mycol. 2018, 39, 359–372. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Fournier, J.; Voglmayr, H. Two unusual new species of Pleosporales: Anteaglonium rubescens and Atrocalyx asturiensis. Sydowia. 2018, 70, 129. [Google Scholar] [CrossRef] [PubMed]
- Kodsueb, R.; Jeewon, R.; Vijaykrishna, D.; Mckenzie, E.H.C.; Lumyong, P.; Lumyong, S.; Hyde, K.D. Systematic revision of Tubeufiaceae based on morphological and molecular data. Fungal Divers. 2006, 21, 105–130. [Google Scholar]
- Zhang, Y.; Fournier, J.; Crous, P.W.; Pointing, S.B.; Hyde, K.D. Phylogenetic and morphological assessment of two new species of Amniculicola and their allies (Pleosporales). Persoonia 2009, 23, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Calatayud, V.; Sanz, M.J.; Aptroot, A. Lichenopyrenis galligena (Pleomassariaceae), a new genus of gall-forming lichenicolous fungi on Leptochidium. Mycol. Res. 2001, 105, 634–637. [Google Scholar] [CrossRef]
- Kruys, Å.; Eriksson, O.E.; Wedin, M. Phylogenetic relationships of coprophilous Pleosporales (Dothideomycetes, Ascomycota), and the classification of some bitunicate taxa of unknown position. Mycol. Res. 2006, 110, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.Y.; Cai, Y.Z.; Hyde, K.D.; Corke, H.; Sun, M. Biodiversity of endophytic fungi associated with 29 traditional Chinese medicinal plants. Fungal Divers. 2008, 33, 61–75. [Google Scholar] [CrossRef] [Green Version]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Kirk, P.M. Index Fungorum. Available online: http://www.indexfungorum.org/names/names.asp (accessed on 25 January 2022).
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Glez-Peña, D.; Gómez-Blanco, D.; Reboiro-Jato, M.; Fdez-Riverola, F.; Posada, D. FALTER: Program oriented conversion of DNA and protein alignments. Nucleic Acids Res. 2010, 38, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Stamatakis, A. RAxML Version 8: A tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nylander, J.A.; Wilgenbusch, J.C.; Warren, D.L.; Swofford, D.L. AWTY (are we there yet?): A system for graphical exploration of MCMC convergence in Bayesian phylogenetics. Bioinformatics 2008, 24, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree v1. 4.0. A Graphical Viewer of Phylogenetic Trees. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 1 August 2021).
- Hyde, K.D. Tropical Australasian fungi. IX. Vaginatispora aquatica gen. et sp. nov. Nova Hedwigia 1995, 61, 233–241. [Google Scholar]
- Zhang, H.; Hyde, K.D.; Zhao, Y.C.; McKenzie, E.H.C. Freshwater ascomycetes: Lophiostoma vaginatispora comb. nov. (Dothideomycetes, Pleosporales, Lophiostomaceae) based on morphological and molecular data. Phytotaxa 2014, 176, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Thambugala, K.M.; Hyde, K.D.; Tanaka, K.; Tian, Q.; Wanasinghe, D.N.; Ariyawansa, H.A.; Jayasiri, S.C.; Boonmee, S.; Camporesi, E.; Hashimoto, A.; et al. Towards a natural classification and backbone tree for Lophiostomataceae, Floricolaceae and Amorosiaceae fam. nov. Fungal Divers. 2015, 74, 199–266. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Jones, E.B.G.; Dissanayake, A.J.; Hyde, K.D. Saprobic Dothideomycetes in Thailand: Vaginatispora appendiculata sp. nov. (Lophiostomataceae) introduced based on morphological and molecular data. Stud. Fungi 2016, 1, 56–58. [Google Scholar] [CrossRef]
- Hashimoto, A.; Hirayama, K.; Takahashi, H.; Matsumura, M.; Okada, G.; Chen, C.Y.; Huang, J.W.; Kakishima, M.; Ono, T.; Tanaka, K. Resolving the Lophiostoma bipolare complex: Insights into complicated delimitations within the Lophiostomataceae genera. Stud. Mycol. 2018, 90, 161–189. [Google Scholar] [CrossRef]
- Devadatha, B.; Sarma, V.V.; Wanasinghe, D.N.; Hyde, K.D.; Jones, E.B.G. Introducing the new Indian mangrove species, Vaginatispora microarmatispora (Lophiostomataceae) based on morphology and multigene phylogenetic analysis. Phytotaxa 2017, 329, 139–149. [Google Scholar] [CrossRef]
- Liew, E.C.Y.; Aptroot, A.; Hyde, K.D. An evaluation of the monophyly of Massarina based on ribosomal DNA sequences. Mycologia 2002, 94, 803–813. [Google Scholar] [CrossRef] [Green Version]
- Jayasiri, S.C.; Hyde, K.D.; Jones, E.; Mckenzie, E.; Ka Runarathna, S.C. Diversity, morphology and molecular phylogeny of dothideomycetes on decaying wild seed pods and fruits. Mycosphere 2019, 10, 1–186. [Google Scholar] [CrossRef]
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.; Rossi, W.; Leonardi, M.; Lee, H.B.; Mun, H.Y.; Houbraken, J.; et al. Fungal diversity notes 1036–1150: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Tian, Q.; Liu, J.K.; Hyde, K.D.; Wanasinghe, D.N.; Boonmee, S.; Jayasiri, S.C.; Luo, Z.L.; Taylor, J.E.; Phillips, A.J.L.; Bhat, D.J.; et al. Phylogenetic relationships and morphological reappraisal of Melanommataceae (Pleosporales). Fungal Divers. 2015, 74, 267–324. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Chen, J.; Dissanayake, A.J.; Doilom, M.; Hongsanan, S.; Jayawardena, R.S.; Jeewon, R.; Perera, R.H.; Thongbai, B.; et al. Thailand’s amazing diversity: Up to 96% of fungi in northern Thailand may be novel. Fungal Divers. 2018, 93, 215–239. [Google Scholar] [CrossRef]
- Matsushima, T. Matsushima mycological memoirs 9. Matsushima Mycol. Memoirs. 1996, 9, 1–30. [Google Scholar]
- Prabhugaonkar, A.; Bhat, D.J. New record of Megacapitula villosa and Paradictyoarthrinium diffractum from India. Mycosphere 2011, 2, 463–467. [Google Scholar]
- Phukhamsakda, C.; Jeewon, R.; McKenzie, E.H.C.; Xu, J.C. Morphology and phylogeny of Phaeoseptum mali sp. nov. (Phaeoseptaceae, Pleosporales) on bark of Malus halliana. Asian J. Mycol. 2019, 2, 118–128. [Google Scholar] [CrossRef]
- Crous, P.W.; Carris, L.M.; Giraldo, A.; Groenewald, J.Z.; Hawksworth, D.L.; Hemández-Restrepo, M.; Jaklitsch, W.M.; Lebrun, M.H.; Schumacher, R.K.; Stielow, J.B.; et al. The Genera of Fungi-fixing the application of the type species of generic names-G2: Allantophomopsis, Latorua, Macrodiplodiopsis, Macrohilum, Milospium, Protostegia, Pyricularia, Robillarda, Rotula, Septoriella, Torula, and Wojnowicia. IMA Fungus 2015, 6, 163–198. [Google Scholar] [CrossRef]
- Li, J.F.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Mapook, A.; Camporesi, E.; Shang, Q.J.; Chukeatirote, E.; Bakhali, A.H.; Hyde, K.D. Molecular taxonomy and morphological characterization reveal new species and new host records of Torula species (Torulaceae, Pleosporales). Mycol. Prog. 2017, 16, 447–461. [Google Scholar] [CrossRef]
- Su, X.J.; Luo, Z.L.; Jeewon, R.; Bhat, D.J.; Bao, D.F.; Li, W.L.; Hao, Y.E.; Su, H.Y.; Hyde, K.D. Morphology and multigene phylogeny reveal new genus and species of Torulaceae from freshwater habitats in northwestern Yunnan, China. Mycol. Prog. 2018, 17, 531–545. [Google Scholar] [CrossRef]
- Li, G.J.; Hyde, K.D.; Zhao, R.L.; Hongsanan, S.; Abdel-Aziz, F.A.; Abdel-Wahab, M.A.; Alvarado, P.; Alves-Silva, G.; Ammirati, J.F.; Ariyawansa, H.A.; et al. Fungal diversity notes 253–366: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 78, 1–237. [Google Scholar] [CrossRef]
- Tanaka, K.; Hashimoto, A.; Matsumura, M.; Sato, T. Brevicollum, a new genus in Neohendersoniaceae, Pleosporales. Mycologia 2017, 109, 608–619. [Google Scholar] [CrossRef]
- Cooke, M.C.; Plowright, C.B. British Sphaeriacei. Grevillea 1879, 7, 77–89. [Google Scholar] [CrossRef]
- Zhang, Y.; Crous, P.W.; Schoch, C.L.; Hyde, K.D. Pleosporales. Fungal Divers. 2012, 53, 1–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyde, K.D.; Taylor, J.E.; Fröhlich, J. Genera of ascomycetes from palms. Fungal Divers. 2000, 2, 1–247. [Google Scholar]
- Tennakoon, D.S.; Jeewon, R.; Kuo, C.H.; Hyde, K.D. Phylogenetic and morphological characterization of Byssosphaeria macarangae sp. nov., and B. taiwanense sp. nov. from Macaranga tanarius. Phytotaxa 2018, 364, 211–226. [Google Scholar] [CrossRef]
- Li, W.Y.; Zhuang, W.Y. Notes on the genus Byssosphaeria (Melanommataceae) from China. Mycosystema 2008, 27, 48–53. [Google Scholar]
- Fabre, J.H. Essai sur les Sphériacées du Départment de Vaucluse. Ann. Sci. Nat. Bot. Sér. 1879, 6, 66–118. [Google Scholar]
- Ariyawansa, H.A.; Jones, E.B.; Suetrong, S.; Alias, S.A.; Kang, J.C.; Hyde, K.D. Halojulellaceae a new family of the order Pleosporales. Phytotaxa 2013, 130, 14–24. [Google Scholar] [CrossRef]
- Zhang, Y.; Fournier, J.; Phookamsak, R.; Bahkali, A.H.; Hyde, K.D. Halotthiaceae fam. nov. (Pleosporales) accommodates the new genus Phaeoseptum and several other aquatic genera. Mycologia 2013, 105, 603–609. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Jones, E.G.; Camporesi, E.; Mortimer, P.E.; Xu, J.; Bahkali, A.H.; Hyde, K.D. The genus Murispora. Cryptogam. Mycol. 2015, 36, 419–448. [Google Scholar] [CrossRef]
- Ariyawansa, H.A.; Hyde, K.D.; Jayasiri, S.C.; Buyck, B.; Chethana, K.T.; Dai, D.Q.; Dai, Y.C.; Daranagama, D.A.; Jayawardena, R.S.; Luecking, R.; et al. Fungal diversity notes 111–252—Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2015, 75, 27–274. [Google Scholar] [CrossRef]
- Hongsanan, S.; Phookamsak, R.; Goonasekara, I.D.; Thambugala, K.M.; Hyde, K.D.; Bhat, J.D.; Suwannarach, N.; Cheewangkoon, R. Introducing a new pleosporale an family Sublophiostomataceae fam. nov. to accommodate Sublophiostoma gen. nov. Sci. Rep. 2021, 11, 9496. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–491: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.P.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Wang, B.; Hyde, K.D.; McKenzie, E.H.C.; Raja, H.A.; Tanaka, K.; Abdel-Wahab, M.A.; Abdel, W.; Wa, F.A.; Doilom, M.; et al. Freshwater Dothideomycetes. Fungal Divers. 2020, 105, 319–575. [Google Scholar] [CrossRef]
- Prasher, I.B.; Verma, R.K. The genus from Himachal Pradesh. Kavaka 2016, 47, 138–142. [Google Scholar]
- Liu, J.K.; Luo, Z.L.; Liu, N.G.; Cheewangkoon, R.; To-Anun, C. Two novel species of Paradictyoarthrinium from decaying wood. Phytotaxa 2018, 338, 285–293. [Google Scholar] [CrossRef]
- Liu, J.K.; Hyde, K.D.; Jones, E.G.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.; McKenzie, E.H.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal diversity notes 1–110: Taxonomic and phylogenetic contributions to fungal species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
- Hyde, K.D.; Chaiwan, N.; Norphanphoun, C.; Boonmee, S.; Camporesi, E.; Chethana, K.W.T.; Dayarathne, M.C.; De Silva, N.I.; Dissanayake, A.J.; Ekanayaka, A.H.; et al. Mycosphere notes 169–224. Mycosphere 2018, 9, 271–430. [Google Scholar] [CrossRef]
- Dayarathne, M.C.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Devadatha, B.; Sarma, V.V.; Khongphinitbunjong, K.; Chomnunti, P.; Hyde, K.D. Morpho-molecular characterization of microfungi associated with marine based habitats. Mycosphere 2020, 11, 1–188. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Mortimer, P.E.; Senwanna, C.; Cheewangkoon, R. Saprobic Dothideomycetes in Thailand: Phaeoseptum hydei sp. nov., a new terrestrial ascomycete in Phaeoseptaceae. Phytotaxa 2020, 449, 149–163. [Google Scholar] [CrossRef]
- Persoon, C.H. Neuer Versuch einer systematischen Eintheilung der Schwämme. Neues Mag. Bot. 1794, 1, 63–80. [Google Scholar]
- Li, J.F.; Jeewon, R.; Mortimer, P.E.; Doilom, M.; Phookamsak, R.; Promputtha, I. Multigene phylogeny and taxonomy of Dendryphion hydei and Torula hydei spp. nov. from herbaceous litter in northern Thailand. PLoS ONE 2020, 15, e0228067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bose, S.K. Studies on Massarina Sacc. and related genera. J. Phytopathol. 1961, 41, 151–213. [Google Scholar] [CrossRef] [Green Version]
- Aptroot, A. A world revision of Massarina (Ascomycota). Nova Hedwigia 1998, 66, 89–162. [Google Scholar] [CrossRef]
- Lu, L.; Hyde, K.D.; Karunarathna, S.C.; Bhat, D.J.; Dai, D.Q.; Jayawardena, R.S.; Tibpromma, S. Crassiparies yunnanensis sp. nov., (Neohendersoniaceae, Pleosporales) from Arabica coffee in China. 2022; under review. [Google Scholar]
- Zhang, C.X.; Xie, D.H.; Chen, Y.F.; Bai, T.Q.; Ni, Z.G. The development status of Yunnan mango industry. China Fruits 2020, 6, 112–117. (In Chinese) [Google Scholar]
- Elsheery, N.I.; Wilske, B.; Zhang, J.L.; Cao, K.F. Seasonal variations in gas exchange and chlorophyll fluorescence in the leaves of five mango cultivars in southern Yunnan, China. J. Hortic. 2007, 82, 855–862. [Google Scholar] [CrossRef]
- Zhang, M.F.; Chen, S.Z.; Chen, X. Statistical Analysis on Ecological and Climatic Adaptability for Cultivation of Mangifera indica in Yunnan. Agric. Sci. Heilongjiang China 2009, 1, 68–69. (In Chinese) [Google Scholar]
- Zhang, C.; Xie, D.; Bai, T.; Luo, X.; Chen, Y. Diversity of a large collection of natural populations of mango (Mangifera indica Linn.) revealed by agro-morphological and quality traits. Diversity 2020, 12, 27. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.Q.; Gan, S.; Li, Y.G. The temporal and spatial variation characteristics analysis of precipitation in Honghe River basin. J. Yunnan Unvi. Nat. Sci. Ed. 2008, 30, 54–60. (In Chinese) [Google Scholar] [CrossRef]
- Sun, L.H. Yuanjiang mango production and variety review. Yunnan Agric. Sci. Tech. 2008, 5, 31–34. (In Chinese) [Google Scholar]
- Zhijia, G.; Xingwu, D.; Bing, L.; Jinming, H.; Jiaonan, H. The spatial distribution and temporal variation of rainfall erosivity in the Yunnan Plateau, Southwest China: 1960–2012. Catena 2016, 145, 291–300. [Google Scholar] [CrossRef]
- Long, Y.Q.; Wang, W.D.; Wang, M.C.; Pu, J.J.; Ni, Z.G.; Chen, Y.F.; Xie, D.H.; Chen, H.R. Investigation and Research on Mango Diseases and Pests in Yunnan. Chin. J. Trop. Crops. 2011, 32, 508–513. (In Chinese) [Google Scholar] [CrossRef]
- Braun, U.; Cook, R.T.A. Taxonomic Manual of Erysiphales (Powdery Mildews); CBS Biodiversity Series 11; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2012; pp. 1–707. [Google Scholar]
- Li, Q.; Bu, J.; Shu, J.; Yu, Z.; Tang, L.; Huang, S.; Guo, T.; Mo, J.; Luo, S.; Solangi, G.S.; et al. Colletotrichum species associated with mango in southern China. Sci. Rep. 2019, 9, 18891. [Google Scholar] [CrossRef] [Green Version]
- Shu, J.; Yu, Z.; Sun, W.; Zhao, J.; Li, Q.; Tang, L.; Guo, T.; Huang, S.; Mo, J.; Hsiang, T.; et al. Identification and characterization of pestalotioid fungi causing leaf spots on mango in southern China. Plant Dis. 2020, 104, 1207–1213. [Google Scholar] [CrossRef]
- Guo, Z.; Yu, Z.; Li, Q.; Tang, L.; Luo, S. Fusarium species associated with leaf spots of mango in china. Microb. Pathog. 2021, 150, 104736. [Google Scholar] [CrossRef]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
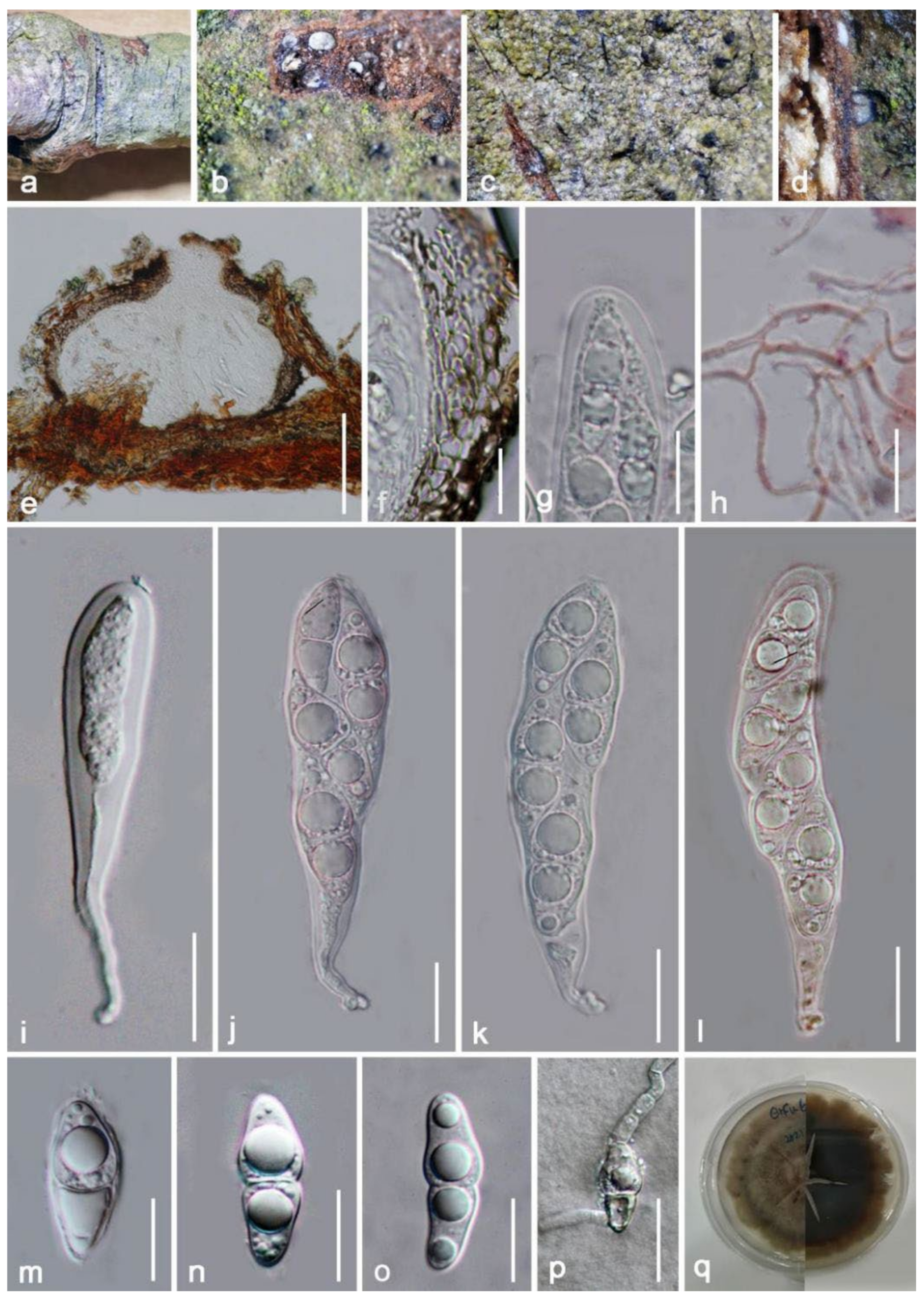{kind=link}
| Order | Family | Species | Host | Distribution | References |
|---|---|---|---|---|---|
| Pleosporales | Massarinaceae | Vaginatispora amygdali | Endocarp of Amygdalus persica | Wakayama, Japan | [56] |
| Dead branch of Mangifera indica | Yunnan, China | This study | |||
| Melanommataceae | Byssosphaeria siamensis | Decaying wood of unidentified host | Chiang Rai, Thailand | [61] | |
| Pandanus sp. | Phang Nga, Thailand | [62] | |||
| Dead endocarp of Mangifera indica | Yunnan, China | This study | |||
| Neomassariaceae | Neomassaria hongheensis | Dead branch of Mangifera indica | Yunnan, China | This study | |
| Parabambusicolaceae | Paramonodictys hongheensis | Dead branch of Mangifera indica | Yunnan, China | This study | |
| Paramonodictys yunnanensis | Dead branch of Mangifera indica | Yunnan, China | This study | ||
| Paradictyoarthriniaceae | Paradictyoarthrinium diffractum | Dead twig in stream | Rustenburg, South Africa | [63] | |
| Spathe of Cocos nucifera | Goa, India | [64] | |||
| Dead stumps and stems of Tectona grandis | Chiang Rai, Thailand | [13,62] | |||
| Dead branch of Mangifera indica | Yunnan, China | This study | |||
| Phaeoseptaceae | Phaeoseptum mali | Decaying twigs of Malus halliana | Yunnan, China | [65] | |
| Dead branch of Mangifera indica | Yunnan, China | This study | |||
| Torulaceae | Torula fici | Ficus sp. | Cuba | [66] | |
| Dead stems of Chromolaena odorata | Chiang Rai, Thailand | [16,67] | |||
| Dead leaf of Pandanus sp. | Chiang Mai, Thailand | [14] | |||
| Decaying cone of Magnolia grandifora | Yunnan, China | [59] | |||
| Decaying fruit pericarp of Garcinia sp. | Ranong, Thailand | [59] | |||
| Submerged decaying wood | Yunnnan, China | [68] | |||
| Dead branch of Mangifera indica | Yunnan, China | This study | |||
| Incertae sedis | Crassiparies quadrisporus | Twigs of Acer sp. | Mie, Janpan | [69,70] | |
| Dead branch of Mangifera indica | Yunnan, China | This study | |||
| Mangifericomes hongheensis | Dead branch of Mangifera indica | Yunnan, China | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, E.-F.; Tibpromma, S.; Karunarathna, S.C.; Phookamsak, R.; Xu, J.-C.; Zhao, Z.-X.; Karunanayake, C.; Promputtha, I. Taxonomy and Phylogeny of Novel and Extant Taxa in Pleosporales Associated with Mangifera indica from Yunnan, China (Series I). J. Fungi 2022, 8, 152. https://doi.org/10.3390/jof8020152
Yang E-F, Tibpromma S, Karunarathna SC, Phookamsak R, Xu J-C, Zhao Z-X, Karunanayake C, Promputtha I. Taxonomy and Phylogeny of Novel and Extant Taxa in Pleosporales Associated with Mangifera indica from Yunnan, China (Series I). Journal of Fungi. 2022; 8(2):152. https://doi.org/10.3390/jof8020152
Chicago/Turabian StyleYang, Er-Fu, Saowaluck Tibpromma, Samantha C. Karunarathna, Rungtiwa Phookamsak, Jian-Chu Xu, Zhen-Xiong Zhao, Chathurika Karunanayake, and Itthayakorn Promputtha. 2022. "Taxonomy and Phylogeny of Novel and Extant Taxa in Pleosporales Associated with Mangifera indica from Yunnan, China (Series I)" Journal of Fungi 8, no. 2: 152. https://doi.org/10.3390/jof8020152
APA StyleYang, E. -F., Tibpromma, S., Karunarathna, S. C., Phookamsak, R., Xu, J. -C., Zhao, Z. -X., Karunanayake, C., & Promputtha, I. (2022). Taxonomy and Phylogeny of Novel and Extant Taxa in Pleosporales Associated with Mangifera indica from Yunnan, China (Series I). Journal of Fungi, 8(2), 152. https://doi.org/10.3390/jof8020152