
The Multifaceted Roles of Fungal Cutinases during Infection


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

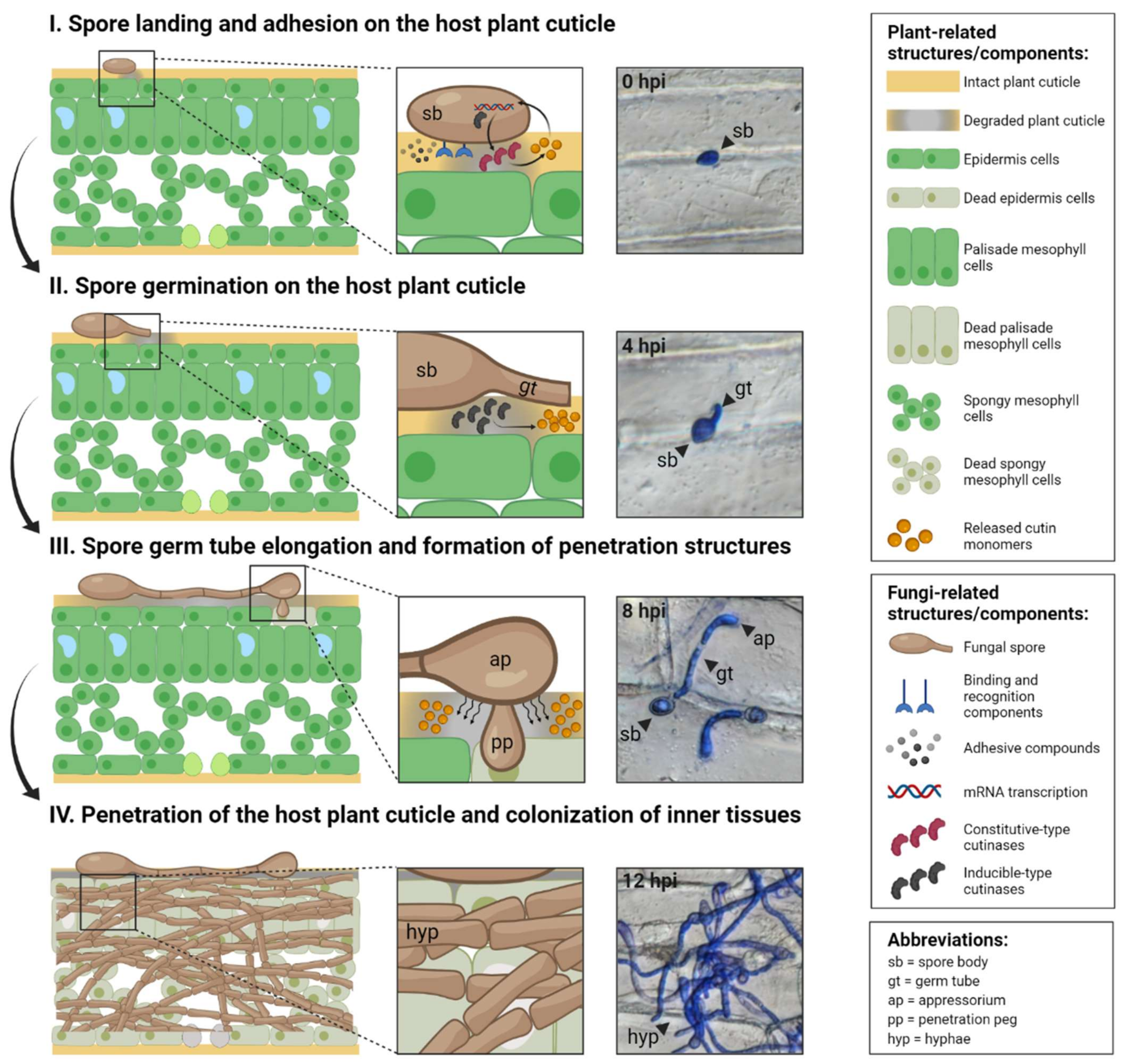
2. Fungal Cutinases Play Multifaceted Roles during Different Stages of Infection
2.1. Spore Landing and Adhesion to the Host Plant Cuticle

2.2. Spore Germination on the Host Plant Cuticle
2.3. Spore Germ Tube Elongation and the Formation of Penetrating Structures
2.4. Penetration of the Host Plant Cuticle and Inner Tissue Colonisation
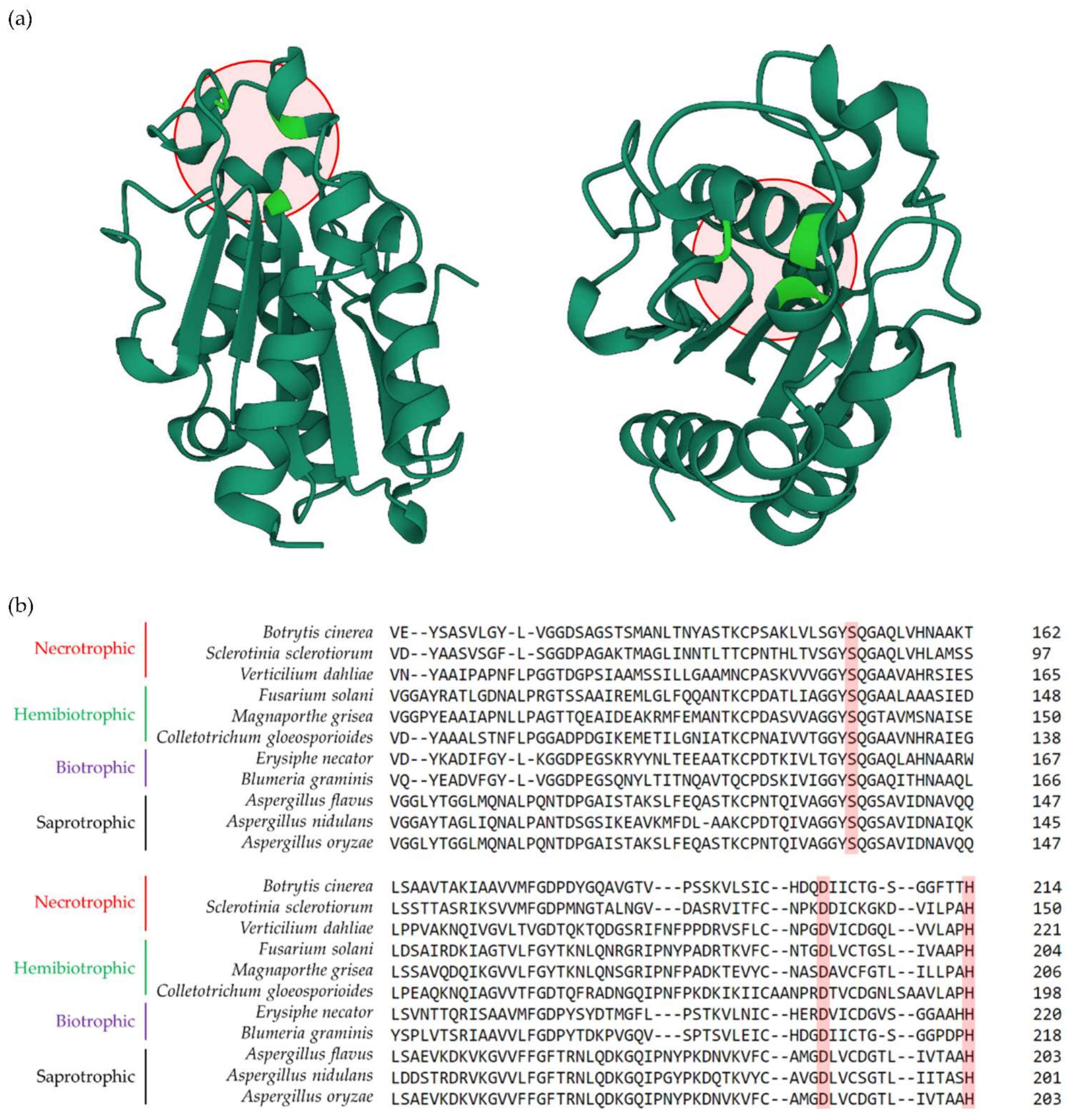
3. The Association between Lifestyles of Fungi and Cutinase Activity

4. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cohen, H.; Szymanski, J.; Aharoni, A. Assimilation of “omics” strategies to study the cuticle layer and suberin lamellae in plants. J. Exp. Bot. 2017, 68, 5389–5400. [Google Scholar] [CrossRef] [PubMed]
- Philippe, G.; Geneix, N.; Petit, J.; Guillon, F.; Sandt, C.; Rothan, C.; Lahaye, M.; Marion, D.; Bakan, B. Assembly of tomato fruit cuticles: A cross-talk between the cutin polyester and cell wall polysaccharides. New Phytol. 2020, 226, 809–822. [Google Scholar] [CrossRef] [PubMed]
- Niklas, K.J.; Cobb, E.D.; Matas, A.J. The evolution of hydrophobic cell wall biopolymers: From algae to angiosperms. J. Exp. Bot. 2017, 68, 5261–5269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollard, M.; Beisson, F.; Li, Y.; Ohlrogge, J.B. Building lipid barriers: Biosynthesis of cutin and suberin. Trends Plant Sci. 2008, 13, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, M.; Keyl, A.; Feussner, I. Wax biosynthesis in response to danger: Its regulation upon abiotic and biotic stress. New Phytol. 2020, 227, 698–713. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Komatsuda, T.; Ma, J.F.; Li, C.; Yamaji, N.; Nevo, E. A functional cutin matrix is required for plant protection against water loss. Plant Signal. Behav. 2011, 6, 1297–1299. [Google Scholar] [CrossRef] [Green Version]
- Isaacson, T.; Kosma, D.K.; Matas, A.J.; Buda, G.J.; He, Y.; Yu, B.; Pravitasari, A.; Batteas, J.D.; Stark, R.E.; Jenks, M.A.; et al. Cutin deficiency in the tomato fruit cuticle consistently affects resistance to microbial infection and biomechanical properties, but not transpirational water loss. Plant J. 2009, 60, 363–377. [Google Scholar] [CrossRef]
- Ziv, C.; Zhao, Z.; Gao, Y.G.; Xia, Y. Multifunctional roles of plant cuticle during plant-pathogen interactions. Front. Plant Sci. 2018, 9, 1088. [Google Scholar] [CrossRef]
- Kolattukudy, P.E. Enzymatic penetration of the plant cuticle by fungal pathogens. Annu. Rev. Phytopathol. 1985, 23, 223–250. [Google Scholar] [CrossRef]
- Longhi, S.; Cambillau, C. Structure-activity of cutinase, a small lipolytic enzyme. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids. 1999, 1441, 185–196. [Google Scholar] [CrossRef]
- Martinez, C.; de Geus, P.; Lauwereys, M.; Matthyssens, G.; Cambillau, C. Fusarium solani cutinase is a lipolytic enzyme with a catalytic serine accessible to solvent. Nature 1992, 356, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Nikolaivits, E.; Kanelli, M.; Dimarogona, M.; Topakas, E. A middle-aged enzyme still in its prime: Recent advances in the field of cutinases. Catalysts 2018, 8, 612. [Google Scholar] [CrossRef] [Green Version]
- Heinen, W. Über Den Enzymatischen Cutin-Abbau v. Mitteilung: Die Lyse von Peroxyd-Brücken Im cutin durch eine peroxydase aus Penicillium spinulosum Thom. Acta Bot. Neerl. 1963, 12, 51–57. [Google Scholar] [CrossRef]
- Shishiyama, J.; Araki, F.; Akai, S. Studies on cutin-esterase II. characteristics of cutin-esterase from Botrytis cinerea and its activity on tomato-cutin. Plant Cell Physiol. 1970, 11, 937–947. [Google Scholar] [CrossRef]
- Purdy, R.E.; Kolattukudy, P.E. Depolymerization isolation of a hydroxy enzyme some and fatty from acid Fusarium solani f. sp. pisi. isolation and some properties of the enzyme. Arch. Biochem. Biophys. 1973, 159, 61–69. [Google Scholar] [CrossRef]
- Purdy, R.E.; Kolattukudy, P.E. Hydrolysis of plant cuticle by plant pathogens. properties of cutinase i, cutinase ii, and a nonspecific esterase isolated from Fusarium solani pisi. Biochemistry 1975, 14, 2832–2840. [Google Scholar] [CrossRef]
- Morid, B.; Zare, R.; Rezaee, S.; Zamani-Zadeh, H.; Hajmansour, S. The relationship between cutinases and the pathogenicity/virulence of Fusarium solani in potato tubers. Phytopathol. Mediterr. 2009, 48, 403–410. [Google Scholar] [CrossRef]
- Arya, G.C.; Sarkar, S.; Manasherova, E.; Aharoni, A.; Cohen, H. The plant cuticle: An ancient guardian barrier set against long-standing rivals. Front. Plant Sci. 2021, 12, 663165. [Google Scholar] [CrossRef]
- Aragón, W.; Reina-Pinto, J.J.; Serrano, M. The intimate talk between plants and microorganisms at the leaf surface. J. Exp. Bot. 2017, 68, 5339–5350. [Google Scholar] [CrossRef]
- Fich, E.A.; Segerson, N.A.; Rose, J.K.C. The plant polyester cutin: Biosynthesis, structure, and biological roles. Annu. Rev. Plant Biol. 2016, 67, 207–233. [Google Scholar] [CrossRef]
- Serrano, M.; Coluccia, F.; Torres, M.; L’Haridon, F.; Métraux, J.P. The cuticle and plant defense to pathogens. Front. Plant Sci. 2014, 5, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longhi, S.; Czjzek, M.; Lamzin, V.; Nicolas, A.; Cambillau, C. Atomic resolution (1.0 Å) crystal structure of Fusarium solani cutinase: Stereochemical analysis. J. Mol. Biol. 1997, 268, 779–799. [Google Scholar] [CrossRef] [PubMed]
- Braun, E.J.; Howard, R.J. Adhesion of fungal spores and germlings to host plant surfaces. Protoplasma 1994, 181, 202–212. [Google Scholar] [CrossRef]
- Osherov, N.; May, G.S. The molecular mechanisms of conidial germination. FEMS Microbiol. Lett. 2001, 199, 153–160. [Google Scholar] [CrossRef]
- Mendgen, K.; Hahn, M.; Deising, H. Morphogenesis and mechanisms of penetration by plant pathogenic fungi. Annu. Rev. Phytopathol. 1996, 34, 367–386. [Google Scholar] [CrossRef] [Green Version]
- Tucker, S.L.; Talbot, N.J. Surface attachment and pre-penetration stage development by plant pathogenic fungi. Annu. Rev. Phytopathol. 2001, 39, 385–417. [Google Scholar] [CrossRef]
- Tudzynski, P.; Sharon, A. Fungal Pathogenicity Genes. In Applied Mycology and Biotechnology; Elsevier: Amsterdam, The Netherlands, 2003; pp. 187–212. [Google Scholar]
- Nicholson, R.L.; Epstein, L. Adhesion of Fungi to the Plant Surface. In The Fungal Spore and Disease Initiation in Plants and Animals; Springer US: Boston, MA, USA, 1991; pp. 3–23. [Google Scholar]
- Köller, W.; Allan, C.R.; Kolattukudy, P.E. Protection of Pisum sativum from Fusarium solani f. sp. pisi by inhibition of cutinase with organophosphorus pesticides. Phytopathology 1982, 72, 1425–1430. [Google Scholar] [CrossRef]
- Lin, T.S.; Kolattukudy, P.E. Induction of a biopolyester hydrolase (cutinase) by low levels of cutin monomers in Fusarium solani f. sp. pisi. J. Bacteriol. 1978, 133, 942–951. [Google Scholar] [CrossRef] [Green Version]
- Soliday, C.L.; Dickman, M.B.; Kolattukudy, P.E. Structure of the cutinase gene and detection of promoter activity in the 5’-flanking region by fungal transformation. J. Bacteriol. 1989, 171, 1942–1951. [Google Scholar] [CrossRef] [Green Version]
- Bajar, A.; Podila, G.K.; Kolattukudy, P.E. Identification of a fungal cutinase promoter that is inducible by a plant signal via a phosphorylated trans-acting factor. Proc. Natl. Acad. Sci. USA 1991, 88, 8208–8212. [Google Scholar] [CrossRef] [Green Version]
- Kämper, J.T.; Kämper, U.; Rogers, L.M.; Kolattukudy, P.E. Identification of regulatory elements in the cutinase promoter from Fusarium solani f. sp. pisi (Nectria haematococca). J. Biol. Chem. 1994, 269, 9195–9204. [Google Scholar] [CrossRef]
- Kolattukudy, P.E.; Rogers, L.M.; Li, D.; Hwang, C.S.; Flaishman, M.A. Surface signaling in pathogenesis. Proc. Natl. Acad. Sci. USA 1995, 92, 4080–4087. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Kolattukudy, P.E. Cloning of cutinase transcription factor 1, a transactivating protein containing Cys6Zn2 binuclear cluster dna-binding motif. J. Biol. Chem. 1997, 272, 12462–12467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Sirakova, T.; Rogers, L.; Ettinger, W.F.; Kolattukudy, P.E. Regulation of constitutively expressed and induced cutinase genes by different zinc finger transcription factors in Fusarium solani f. sp. pisi (Nectria Haematococca). J. Biol. Chem. 2002, 277, 7905–7912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gindro, K.; Pezet, R. Evidence for a constitutive cytoplasmic cutinase in ungerminated conidia of Botrytis cinerea Pers.: Fr. FEMS Microbiol. Lett. 1997, 149, 89–92. [Google Scholar] [CrossRef]
- Leroch, M.; Kleber, A.; Silva, E.; Coenen, T.; Koppenhöfer, D.; Shmaryahu, A.; Valenzuela, P.D.T.; Hahn, M. Transcriptome profiling of Botrytis cinerea conidial germination reveals upregulation of infection-related genes during the prepenetration stage. Eukaryot. Cell 2013, 12, 614–626. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Hou, J.; Wang, Y.; Jin, Y.; Borth, W.; Zhao, F.; Liu, Z.; Hu, J.; Zuo, Y. Genome-wide identification, classification and expression analysis in fungal–plant interactions of cutinase gene family and functional analysis of a putative ClCUT7 in Curvularia lunata. Mol. Genet. Genom. 2016, 291, 1105–1115. [Google Scholar] [CrossRef]
- Davies, K.A.A.; De Lorono, I.; Foster, S.J.J.; Li, D.; Johnstone, K.; Ashby, A.M.M. Evidence for a role of cutinase in pathogenicity of Pyrenopeziza brassicae on Brassicas. Physiol. Mol. Plant Pathol. 2000, 57, 63–75. [Google Scholar] [CrossRef]
- Oliver, R.P.; Ipcho, S.V.S. Arabidopsis pathology breathes new life into the necrotrophs-vs.-biotrophs classification of fungal pathogens. Mol. Plant Pathol. 2004, 5, 347–352. [Google Scholar] [CrossRef]
- Skamnioti, P.; Gurr, S.J. Magnaporthe grisea cutinase2 mediates appressorium differentiation and host penetration and is required for full virulence. Plant Cell 2007, 19, 2674–2689. [Google Scholar] [CrossRef] [Green Version]
- Auyong, A.S.; Ford, R.; Taylor, P.W. The Role of Cutinase and its impact on pathogenicity of Colletotrichum truncatum. J. Plant Pathol. Microbiol. 2015, 06, 1000259. [Google Scholar] [CrossRef] [Green Version]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal effectors and plant susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef] [PubMed]
- Deising, H.; Nicholson, R.L.; Haug, M.; Howard, R.J.; Mendgen, K. Adhesion pad formation and the involvement of cutinase and esterases in the attachment of uredospores to the host cuticle. Plant Cell. 1992, 4, 1101–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baltussen, T.J.H.; Zoll, J.; Verweij, P.E.; Melchers, W.J.G. Molecular mechanisms of conidial germination in Aspergillus spp. Microbiol. Mol. Biol. Rev. 2019, 84, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Pascholati, S.F.; Yoshioka, H.; Kunoh, H.; Nicholson, R.L. Preparation of the infection court by Erysiphe graminis f. sp. hordei: Cutinase is a component of the conidial exudate. Physiol. Mol. Plant Pathol. 1992, 41, 53–59. [Google Scholar] [CrossRef]
- Pascholati, S.F.; Deising, H.; Leiti, B.; Anderson, D.; Nicholson, R.L. Cutinase and non-specific esterase activities in the conidial mucilage of Colletotrichum graminicola. Physiol. Mol. Plant Pathol. 1993, 42, 37–51. [Google Scholar] [CrossRef]
- Van Kan, J.A.L.; Van’t Klooster, J.W.; Wagemakers, C.A.M.; Dees, D.C.T.; Van Der Vlugt-Bergmans, C.J.B. Cutinase A of Botrytis cinerea is expressed, but not essential, during penetration of gerbera and tomato. Mol. Plant Microbe Interact. 1997, 10, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Van Der Vlugt-Bergmans, C.J.B.; Wagemakers, C.A.M.; Van Kan, J.A.L. Cloning and expression of the cutinase a gene of Botrytis cinerea. Mol. Plant Microbe Interact. 1997, 10, 21–29. [Google Scholar] [CrossRef]
- Woloshuk, C.P.; Kolattukudy, P.E. Mechanism by which contact with plant cuticle triggers cutinase gene expression in the spores of Fusarium solani f. sp. pisi. Proc. Natl. Acad. Sci. USA 1986, 83, 1704–1708. [Google Scholar] [CrossRef] [Green Version]
- Francis, S.A.; Dewey, F.M.; Gurr, S.J. The Role of Cutinase in germling development and infection by Erysiphe graminis f. sp. hordei. Physiol. Mol. Plant Pathol. 1996, 49, 201–211. [Google Scholar] [CrossRef]
- Gilbert, R.D.; Johnson, A.M.; Dean, R.A. Chemical signals responsible for appressorium formation in the rice blast fungus Magnaporthe grisea. Physiol. Mol. Plant Pathol. 1996, 48, 335–346. [Google Scholar] [CrossRef]
- Van den Ende, G.; Linskens, H.F. Cutinolytic enzymes in relation to pathogenesis. Annu. Rev. Phytopathol. 1974, 12, 247–258. [Google Scholar] [CrossRef]
- Lin, T.S.; Kolattukudy, P.E. Structural studies on cutinase, a glycoprotein containing novel amino acids and glucuronic acid amide at the N terminus. Eur. J. Biochem. 1980, 106, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.M.L.; Aires-Barros, M.R.; Cabral, J.M.S. Cutinase: From molecular level to bioprocess development. Biotechnol. Bioeng. 1999, 66, 17–34. [Google Scholar] [CrossRef]
- Tenhaken, R.; Arnemann, M.; Köhler, G.; Barz, W. Characterization and cloning of cutinase from Ascochyta rabiei. Z. Naturforsch. C J. Biosci. 1997, 52, 197–208. [Google Scholar] [CrossRef]
- Wang, G.-Y.; Hammock, B.; Lee, Y.-M.; Bostock, R. Affinity purification and characterization of a cutinase from the fungal plant pathogen Monilinia fructicola (Wint.) honey. Arch. Biochem. Biophys. 2000, 382, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Belbahri, L.; Calmin, G.; Mauch, F.; Andersson, J.O. Evolution of the cutinase gene family: Evidence for lateral gene transfer of a candidate Phytophthora virulence factor. Gene 2008, 408, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Bashi, Z.D.; Roger Rimmer, S.; Khachatourians, G.G.; Hegedus, D.D. Factors governing the regulation of Sclerotinia sclerotiorum cutinase a and polygalacturonase 1 during different stages of infection. Can. J. Microbiol. 2012, 58, 605–616. [Google Scholar] [CrossRef]
- D’Enfert, C. Fungal spore germination: Insights from the molecular genetics of Aspergillus nidulans and Neurospora crassa. Fungal Genet. Biol. 1997, 21, 163–172. [Google Scholar] [CrossRef]
- Zhang, Z.; Henderson, C.; Perfect, E.; Carver, T.L.W.W.; Thomas, B.J.; Skamnioti, P.; Gurr, S.J. Of genes and genomes, needles and haystacks: Blumeria graminis and functionality. Mol. Plant Pathol. 2005, 6, 561–575. [Google Scholar] [CrossRef]
- DeZwaan, T.M.; Carroll, A.M.; Valent, B.; Sweigard, J.A. Magnaporthe grisea Pth11p is a novel plasma membrane protein that mediates appressorium differentiation in response to inductive substrate cues. Plant Cell 1999, 11, 2013–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köller, W.; Smith, F.D.; Reynolds, K.L. Phenotypic instability of flusilazole sensitivity in Venturia inaequalis. Plant Pathol. 1991, 40, 608–611. [Google Scholar] [CrossRef]
- Agrios, G. Plant Pathology, 5th ed.; Elsevier Academic Press: Amsterdam, The Netherlands, 2005; ISBN 0080473784. [Google Scholar]
- Gui, Y.J.; Zhang, W.Q.; Zhang, D.D.; Zhou, L.; Short, D.P.G.; Wang, J.; Ma, X.F.; Li, T.G.; Kong, Z.Q.; Wang, B.L.; et al. A Verticillium dahliae extracellular cutinase modulates plant immune responses. Mol. Plant Microbe Interact. 2018, 31, 260–273. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Zhang, B.; Gai, Y.; Sun, X.; Chung, K.R.; Li, H. Cell-wall-degrading enzymes required for virulence in the host selective toxin-producing necrotroph Alternaria alternata of citrus. Front. Microbiol. 2019, 10, 1–15. [Google Scholar] [CrossRef]
- Fu, H.; Chung, K.R.; Gai, Y.; Mao, L.; Li, H. The basal transcription factor II H subunit Tfb5 is required for stress response and pathogenicity in the tangerine pathotype of Alternaria alternata. Mol. Plant. Pathol. 2020, 21, 1337–1352. [Google Scholar] [CrossRef] [PubMed]
- Maiti, I.B.; Kolattukudy, P.E. Prevention of fungal infection of plants by specific inhibition of cutinase a relationship between DNA helix stability and recognition sites for RNA polymerase. Sceince 1979, 205, 507–508. [Google Scholar] [CrossRef]
- Dantzig, A.H.; Zuckerman, S.H.; Andonov-Roland, M.M. Isolation of a Fusarium solani mutant reduced in cutinase activity and virulence. J. Bacteriol. 1986, 168, 911–916. [Google Scholar] [CrossRef] [Green Version]
- Rogers, L.M.; Flaishman, M.; Kolattukudy, P.E. Cutinase gene disruption in Fusarium solani f. sp. pisi decreases Its Virulence on Pea. Plant Cell 1994, 6, 935–945. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Ashby, A.M.; Johnstone, K. Molecular evidence that the extracellular cutinase Pbc1 is required for pathogenicity of Pyrenopeziza brassicae on oilseed rape. Mol. Plant Microbe Interact. 2003, 16, 545–552. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, J.; Li, D.W.; Zheng, L.; Huang, J. CglCUT1 gene required for cutinase activity and pathogenicity of Colletotrichum gloeosporioides causing anthracnose of Camellia oleifera. Eur. J. Plant Pathol. 2017, 147, 103–114. [Google Scholar] [CrossRef]
- Lee, M.H.; Chlu, C.M.; Roubtsova, T.; Chou, C.M.; Bostock, R.M. Overexpression of a redox-regulated cutinase gene, MfCUTI, increases virulence of the brown rot pathogen Monilinia fructicola on Prunus spp. Mol. Plant Microbe Interact. 2010, 23, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Dickman, M.B.; Podila, G.K.; Kolattukudy, P.E. Insertion of cutinase gene into a wound pathogen enables it to infect intact host. Nature 1989, 342, 446–448. [Google Scholar] [CrossRef]
- Rajarammohan, S. Redefining Plant-Necrotroph Interactions: The Thin Line Between Hemibiotrophs and Necrotrophs. Front. Microbiol. 2021, 12, 673518. [Google Scholar] [CrossRef] [PubMed]
- Van Kan, J.A.; Shaw, M.W.; Grant-Downton, R.T. Botrytis species: Relentless necrotrophic thugs or endophytes gone rogue? Mol. Plant Pathol. 2014, 15, 957–961. [Google Scholar] [CrossRef] [PubMed]
- Kabbage, M.; Yarden, O.; Dickman, M.B. Pathogenic attributes of Sclerotinia sclerotiorum: Switching from a biotrophic to necrotrophic lifestyle. Plant Sci. 2015, 233, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Spanu, P.D.; Panstruga, R. Editorial: Biotrophic plant-microbe interactions. Front. Plant Sci. 2017, 8, 192. [Google Scholar] [CrossRef] [Green Version]
- Skamnioti, P.; Furlong, R.F.; Gurr, S.J. Evolutionary history of the ancient cutinase family in five filamentous ascomycetes reveals differential gene duplications and losses and in Magnaporthe grisea shows evidence of sub- and neo-functionalization. New Phytol. 2008, 180, 711–721. [Google Scholar] [CrossRef]
- Schmitt, I. Fruiting Body Evolution in the Ascomycota: A Molecular Perspective Integrating Lichenized and Non-Lichenized Groups. In Evolution of Fungi and Fungal-Like Organisms; Springer: Berlin/Heidelberg, Germany, 2011; pp. 187–204. [Google Scholar]
- Whelan, S.; Goldman, N. A general empirical model of protein evolution derived from multiple protein families using a maximum-likelihood approach. Mol. Biol. Evol. 2001, 18, 691–699. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Köller, W.; Yao, C.; Trial, F.; Parker, P.M. Role of cutinase in the invasion of plants. Can. J. Bot. 1995, 73, 1109–1118. [Google Scholar] [CrossRef]
- Novy, V.; Carneiro, L.V.; Shin, J.H.; Larsbrink, J.; Olsson, L. Phylogenetic analysis and in-depth characterization of functionally and structurally diverse CE5 cutinases. J. Biol. Chem. 2021, 297, 101302. [Google Scholar] [CrossRef] [PubMed]
- Krijger, J.J.; Thon, M.R.; Deising, H.B.; Wirsel, S.G.R. Compositions of fungal secretomes indicate a greater impact of phylogenetic histroy than lifestyle adaptation. BMC Genom. 2014, 15, 722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandaranayaka, E.P.J.; Frenkel, O.; Elad, Y.; Prusky, D.; Harel, A. Network analysis exposes core functions in major lifestyles of fungal and oomycete plant pathognes. BMC Genom. 2019, 20, 1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arya, G.C.; Cohen, H. The Multifaceted Roles of Fungal Cutinases during Infection. J. Fungi 2022, 8, 199. https://doi.org/10.3390/jof8020199
Arya GC, Cohen H. The Multifaceted Roles of Fungal Cutinases during Infection. Journal of Fungi. 2022; 8(2):199. https://doi.org/10.3390/jof8020199
Chicago/Turabian StyleArya, Gulab Chand, and Hagai Cohen. 2022. "The Multifaceted Roles of Fungal Cutinases during Infection" Journal of Fungi 8, no. 2: 199. https://doi.org/10.3390/jof8020199
APA StyleArya, G. C., & Cohen, H. (2022). The Multifaceted Roles of Fungal Cutinases during Infection. Journal of Fungi, 8(2), 199. https://doi.org/10.3390/jof8020199