
Experimental Transmission of the Yeast, Metschnikowia bicuspidata, in the Chinese Mitten Crab, Eriocheir sinensis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Yeast Stains
2.3. Horizontal Transmission
2.4. Vertical Transmission Detection
2.5. DNA Extraction and PCR Assay
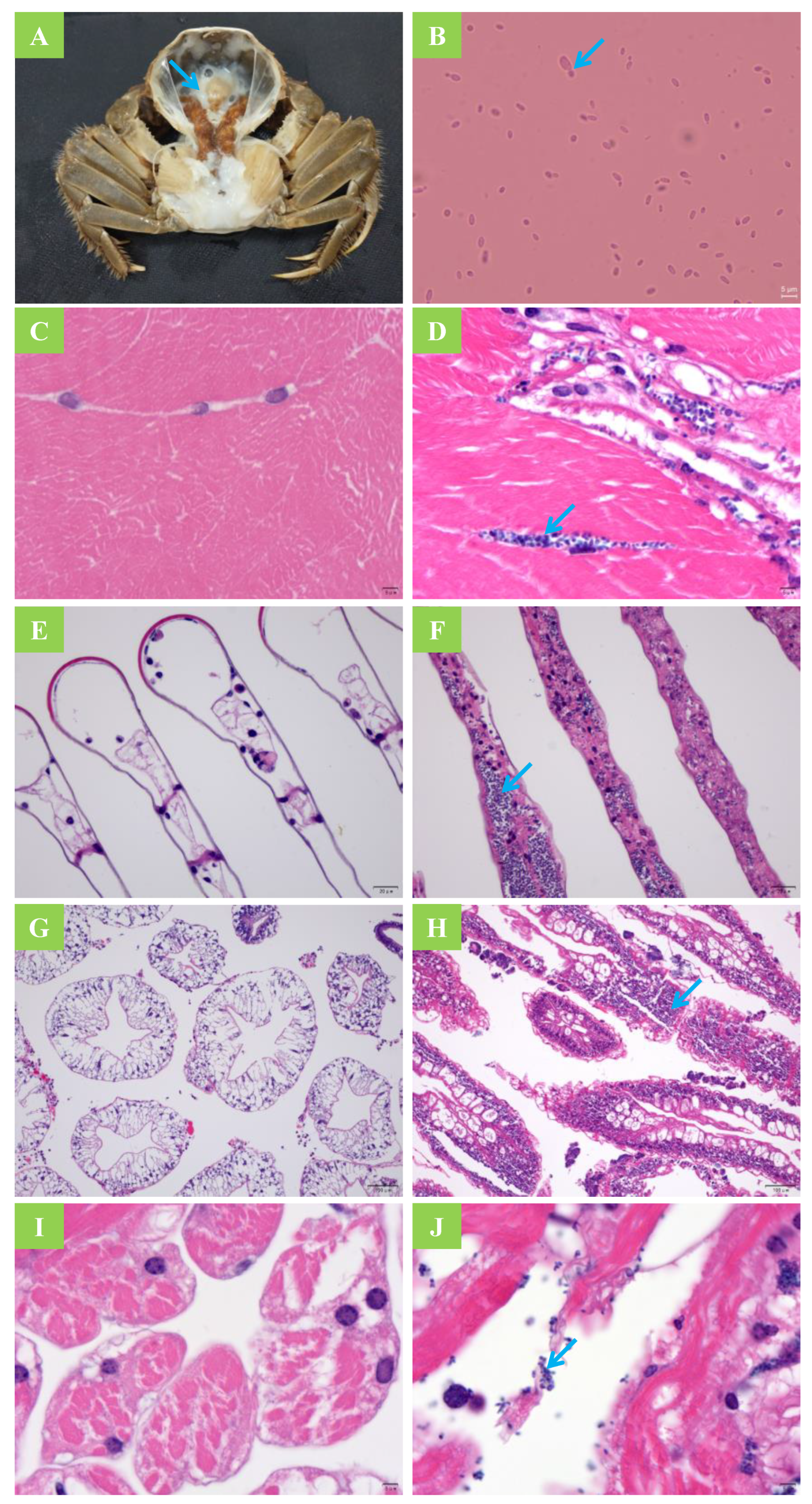
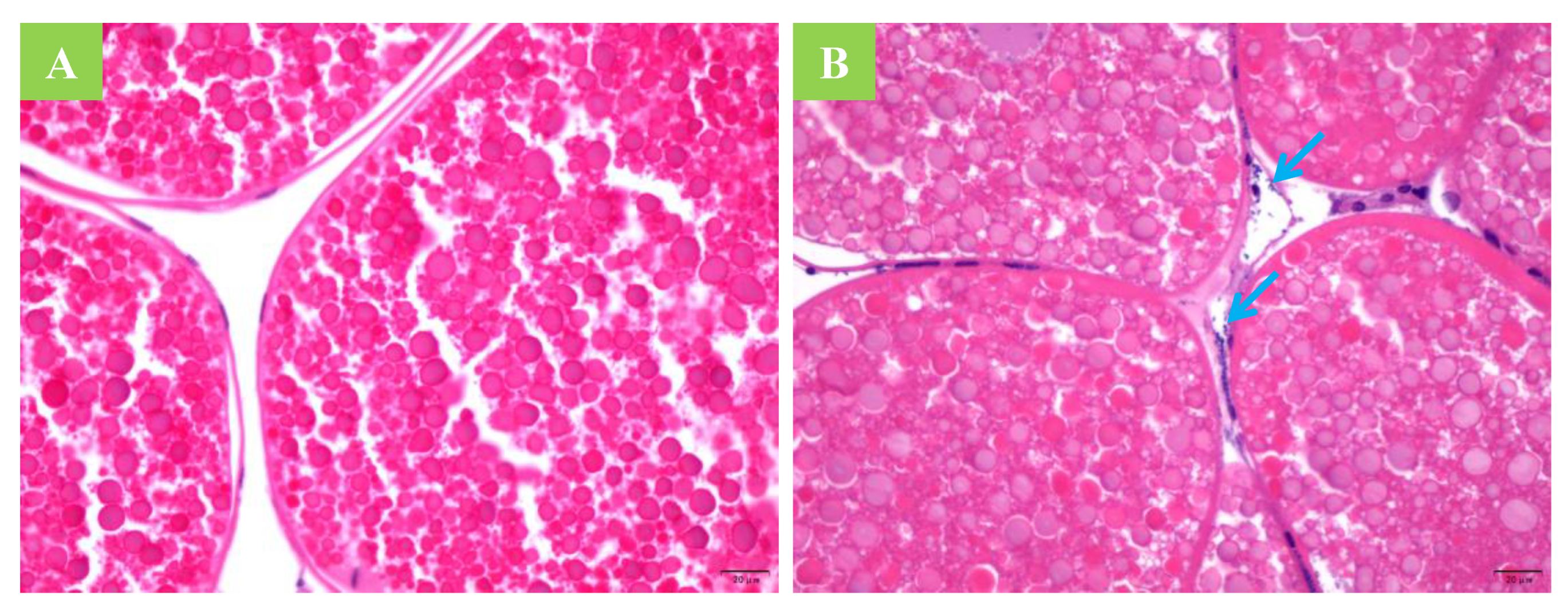
2.6. Histological Section Observation and Smear Examination
2.7. Statistical Analyses
3. Results
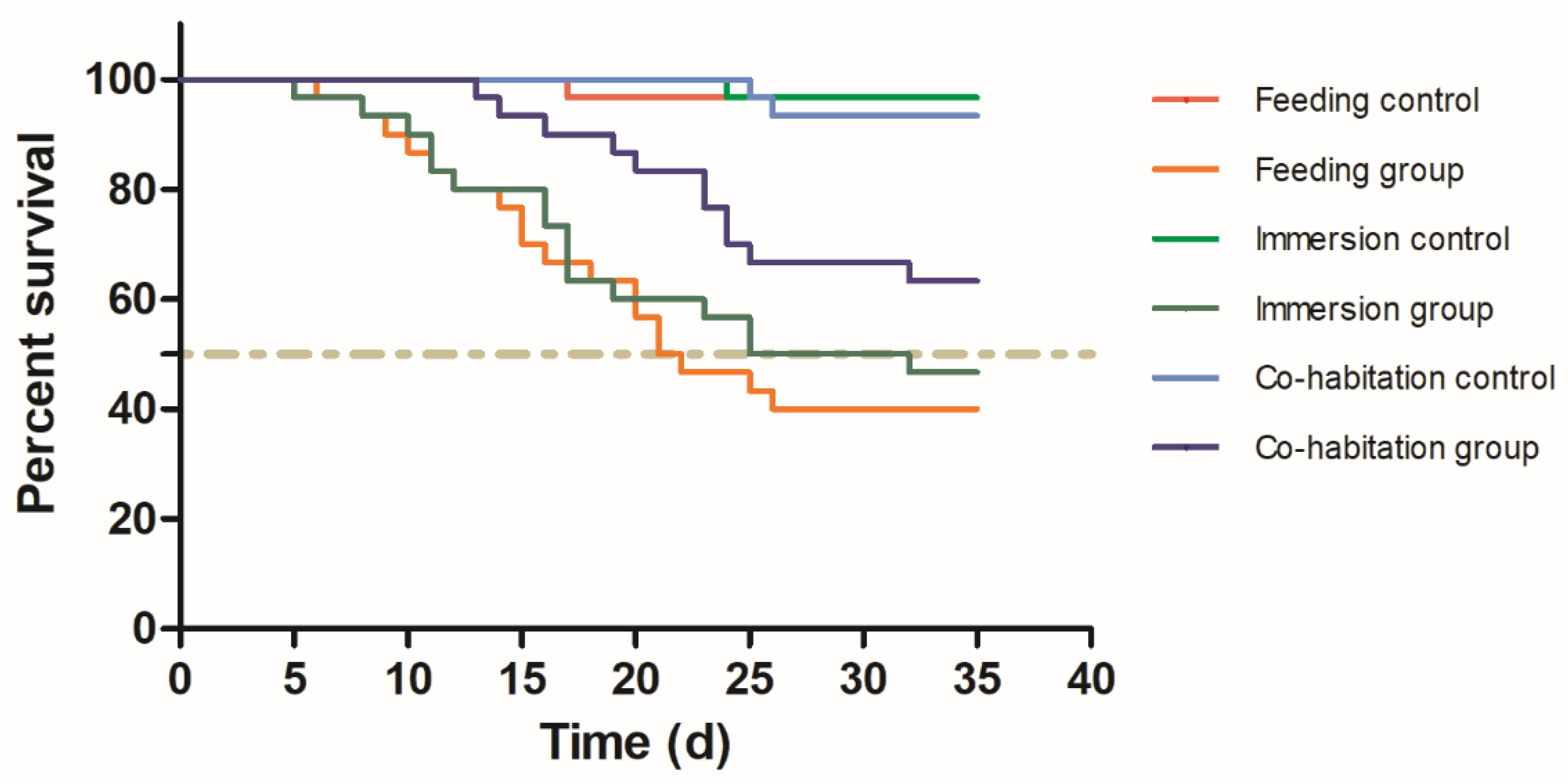
3.1. Horizontal Transmission
3.2. Vertical Transmission
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stentiford, G.D. Diseases of the European edible crab (Cancer pagurus): A review. J. Mar. Sci. 2008, 65, 1578–1592. [Google Scholar] [CrossRef] [Green Version]
- Guerra, R.S.; do Nascimento, M.M.F.; Miesch, S.; Najafzadeh, M.J.; Ribeiro, R.O.; Ostrensky, A.; de Hoog, S.; Vicente, V.; Boeger, W.A. Black yeast biota in the mangrove, in search of the origin of the lethargic crab disease (LCD). Mycopathologia 2013, 175, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.Q. A study on pathology of the disease caused by Torulopsis Mogii in giant freshwater prawn Macrobrachium rosenbergii. J. Fish. China 1996, 20, 13–17. (In Chinese) [Google Scholar]
- Xu, W.; Xu, H.; Shi, H.; Qian, D. Preliminary study on Candida oleophila disease in Portunus trituberculatus. J. Fish. China 2005, 29, 831–836. (In Chinese) [Google Scholar]
- Lu, C.C.; Tang, K.F.J.; Chen, S.N. Identification and genetic characterization of yeasts isolated from freshwater prawns, Macrobrachium rosenbergii de man, in Taiwan. J. Fish Dis. 1998, 21, 185–192. [Google Scholar] [CrossRef]
- Chen, S.C.; Chen, T.H.; Wang, P.C.; Chen, Y.C.; Liaw, L.L. Metschnikowia bicuspidate and Enterococcus faecium coinfection in the giant freshwater prawn Macrobrachium rosenbergii. Dis. Aquat. Org. 2003, 55, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Vicente, V.A.; Orélis-Ribeiro, R.; Najafzadeh, M.J.; Sun, J.F.; Guerra, R.S.; Miesch, S.; Ostrensky, A.; Meis, J.F.; Klaassen, C.H.; Hoog, G.S.; et al. Black yeastlike fungi associated with lethargic crab disease (LCD) in the mangrove-land crab, Ucides cordatus (Ocypodidae). Vet. Microbiol. 2012, 158, 109–122. [Google Scholar] [CrossRef]
- Wang, X.; Chi, Z.; Yue, L.; Li, J.; Li, M.; Wu, L. A marine killer yeast against the pathogenic yeast strain in crab (Portunus trituberculatus) and an optimization of the toxin production. Microbiol. Res. 2007, 162, 77–85. [Google Scholar] [CrossRef]
- Fishery Administration of the Ministry of Agriculture and Rural Areas. China Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2020; p. 34.
- Sun, N.; Bao, J.; Liang, F.; Liu, F.; Jiang, H.; Li, X. Prevalence of ‘milky disease’ caused by Metschnikowia bicuspidata in Eriocheir sinensis in Panjin city, China. Aquac. Res. 2022, 53, 1136–1140. [Google Scholar] [CrossRef]
- Bao, J.; Jiang, H.; Shen, H.; Xing, Y.; Feng, C.; Li, X.; Chen, Q. First description of milky disease in the Chinese mitten crab Eriocheir sinensis caused by the yeast Metschnikowia bicuspidata. Aquaculture 2021, 532, 735984. [Google Scholar] [CrossRef]
- Ma, H.; Lu, X.; Liu, J.; Guo, S.; Zhao, X.; Ye, S. Metschnikowia bicuspidata isolated from milky diseased Eriocheir sinensis: Phenotypic and genetic characterization, antifungal susceptibility and challenge models. J. Fish Dis. 2021, 45, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Codreanu, R.; Codreanu-Balcescu, D. On two Metschnikowia yeast species producing hemocoelic infections in Daphnia magna and Artemia salina (Crustacea, Phyllopoda) from Romania. J. Invertebr. Pathol. 1981, 37, 22–27. [Google Scholar] [CrossRef]
- Shaw, C.L.; Bilich, R.; O’Brien, B.; Cáceres, C.E.; Hall, S.R.; James, T.Y.; Duffy, M.A. Genotypic variation in an ecologically important parasite is associated with host species, lake, and spore size. Parasitology 2021, 148, 1303–1312. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.M.; Strom, M.S. Infection and mortality by the yeast Metschnikowia bicuspidata var. bicuspidata in chinook salmon fed live adult brine shrimp (Artemia franciscana). Aquaculture 2003, 220, 43–57. [Google Scholar] [CrossRef]
- Chen, S.C.; Chen, Y.C.; Kwang, J.; Manopo, I.; Wang, P.C.; Chaung, H.C.; Liaw, L.L.; Chiu, S.H. Metschnikowia bicuspidata dominates in Taiwanese cold-weather yeast infections of Macrobrachium rosenbergii. Dis. Aquat. Org. 2007, 75, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.L.; Shan, J.; Chen, Y.E.; Li, Z. Study on pathogens and pathogenesis of emulsification disease of Portunus trituberculatus. Adv. Mar. Sci. 2006, 24, 526–531. [Google Scholar]
- Merrill, T.E.S.; Cáceres, C.E. Within-host complexity of a plankton-parasite interaction. Ecology 2018, 99, 2864–2867. [Google Scholar] [CrossRef]
- Lachance, M.A.; Miranda, M.; Miller, M.W.; Phaff, H.J. Dehiscence and active spore release in pathogenic strains of the yeast Metschnikowia bicuspidata var. australis: Possible predatory implication. Can. J. Microbiol. 1976, 22, 1756–1761. [Google Scholar] [CrossRef]
- Zhang, H.Q.; Chi, Z.; Liu, G.L.; Zhang, M.; Hu, Z.; Chi, Z.M. Metschnikowia bicuspidate associated with a milky disease in Eriocheir sinensis and its effective treatment by Massoia lactone. Microbiol. Res. 2021, 242, 126641. [Google Scholar] [CrossRef]
- Metschnikoff, E. A disease of Daphnia caused by a yeast. A contribution to the theory of phagocytes as agents for attack on disease-causing organisms. Archiv. Pathol. Anat. Physiol. Klin. Med. 1884, 96, 177–195. [Google Scholar] [CrossRef]
- Jiang, H.; Bao, J.; Xing, Y.; Feng, C.; Li, X.; Chen, Q. Proteomic Analysis of the Hemolymph after Metschnikowia bicuspidata Infection in the Chinese Mitten Crab Eriocheir sinensis. Front. Immunol. 2021, 12, 1046. [Google Scholar] [CrossRef] [PubMed]
- Bright, M.; Bulgheresi, S. A complex journey: Transmission of microbial symbionts. Nat. Rev. Microbiol. 2010, 8, 218–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stentiford, G.D.; Feist, S.W.; Stone, D.M.; Bateman, K.S.; Dunn, A.M. Microsporidia: Diverse, dynamic, and emergent pathogens in aquatic systems. Trends Parasitol. 2013, 29, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, J.; Mrugała, A.; Kozubíková-Balcarová, E.; Petrusek, A. Hosts and transmission of the crayfish plague pathogen Aphanomyces astaci: A review. J. Fish Dis. 2017, 40, 127–140. [Google Scholar] [CrossRef]
- Lipsitch, M.; Siller, S.; Nowak, M.A. The evolution of virulence in pathogens with vertical and horizontal transmission. Evolution 1996, 50, 1729–1741. [Google Scholar] [CrossRef]
- Alizon, S. Parasite co-transmission and the evolutionary epidemiology of virulence. Evol. Int. J. Org. Evol. 2013, 67, 921–933. [Google Scholar] [CrossRef]
- Pagán, I.; Montes, N.; Milgroom, M.G.; García-Arenal, F. Vertical transmission selects for reduced virulence in a plant virus and for increased resistance in the host. PLoS Pathog. 2014, 10, e1004293. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.Z.; Pang, Y.Y.; Huang, G.Y.; Xu, M.J.; Zhang, C.; He, L.; Lv, J.H.; Song, Y.M.; Song, X.Z.; Cheng, Y.X. The serotonin or dopamine by cyclic adenosine monophosphate-protein kinase a pathway involved in the agonistic behaviour of Chinese mitten crab, Eriocheir sinensis. Physiol. Behav. 2019, 209, 112621. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, Q.; Fan, S.; Sun, N.; Li, X.D.; Zheng, Y. Aggressive behavior variation and experience effects in three families of juvenile Chinese mitten crab (Eriocheir sinensis). Behav. Processes 2019, 165, 44–50. [Google Scholar] [CrossRef]
- Lotz, J.M.; Soto, M.A. Model of white spot syndrome virus (WSSV) epidemics in Litopenaeus vannamei. Dis. Aquat. Org. 2002, 50, 199–209. [Google Scholar] [CrossRef] [Green Version]
- Dey, B.K.; Dugassa, G.H.; Hinzano, S.M.; Bossier, P. Causative agent, diagnosis and management of white spot disease in shrimp: A review. Rev. Aquac. 2020, 12, 822–865. [Google Scholar] [CrossRef]
- Soto, M.A.; Lotz, J.M. Epidemiological parameters of white spot syndrome virus infections in Litopenaeus vannamei and L. setiferus. J. Invertebr. Pathol. 2001, 78, 9–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, M.; Summerfelt, R.C. The role of mechanical injury in an experimental transmission of Flexibacter columnaris to fingerling walleye. J. Iowa Acad. Sci. JIAS 1991, 98, 93–98. [Google Scholar]
- Bader, J.A.; Nusbaum, K.E.; Shoemaker, C.A. Comparative challenge model of Flavobacterium columnare using abraded and unabraded channel catfish, Ictalurus punctatus (Rafinesque). J. Fish Dis. 2003, 26, 461–467. [Google Scholar] [CrossRef]
- Bader, J.A.; Moore, S.A.; Nusbaum, K.E. The effect of cutaneous injury on a reproducible immersion challenge model for Flavobacterium columnare infection in channel catfish (Ictalurus punctatus). Aquaculture 2006, 253, 1–9. [Google Scholar] [CrossRef]
- Stentiford, G.D.; Evans, M.; Bateman, K.; Feist, S.W. Co-infection by a yeast-like organism in Hematodinium-infected European edible crabs Cancer pagurus and velvet swimming crabs Necora puber from the English Channel. Dis. Aquat. Org. 2003, 54, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Sun, N.; Lu, X.; Liu, J.; Guo, S.; Zhao, X.; Ye, S. Isolation and identification of pathogen in milky disease of Chinese mitten crab Eriocheir sinensis in Liaoning Province. J. Dalian Ocean. Univ. 2020, 35, 714–718. (In Chinese) [Google Scholar]
- Wang, L.; Yue, L.; Chi, Z.; Wang, X. Marine killer yeasts active against a yeast strain pathogenic to crab Portunus trituberculatus. Dis. Aquat. Org. 2008, 80, 211–218. [Google Scholar] [CrossRef]
- Buzdar, M.A.; Chi, Z.; Wang, Q.; Hua, M.X.; Chi, Z.M. Production, purification, and characterization of a novel killer toxin from Kluyveromyces siamensis against a pathogenic yeast in crab. Appl. Microbiol. Biotechnol. 2011, 91, 1571–1579. [Google Scholar] [CrossRef]
- Tan, C.; Wang, L.; Xue, Y.; Yu, G.; Yang, S.; Lin, S. Marine killer yeast Metschnikowia saccharicola active against pathogenic yeast in crab and an optimization of the toxin production. Afr. J. Biotechnol. 2018, 17, 668–679. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Crab Number | Number of Deaths at Different Times | Cumulative Mortality (%) | Infection Rate (%) | |||||
|---|---|---|---|---|---|---|---|---|
| 1–7 d | 8–14 d | 15–21 d | 22–28 d | 29–35 d | ||||
| Feeding control | 30 | 0 | 0 | 1 | 0 | 0 | 3.3 ± 5.8 C | 0 C |
| Feeding group | 30 | 1 | 6 | 8 | 3 | 0 | 60 ± 10 A | 76.7 ± 10 A |
| Immersion control | 30 | 0 | 0 | 0 | 1 | 0 | 3.3 ± 5.8 C | 0 C |
| Immersion group | 30 | 1 | 5 | 6 | 3 | 1 | 53.3 ± 11.5 AB | 66.7 ± 5.8 AB |
| Cohabitation control | 30 | 0 | 0 | 0 | 2 | 0 | 6.7 ± 5.8 C | 0 C |
| Cohabitation group | 30 | 0 | 2 | 3 | 5 | 1 | 36.7 ± 5.8 B | 53.3 ± 11.5 B |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, H.; Bao, J.; Cao, G.; Xing, Y.; Feng, C.; Hu, Q.; Li, X.; Chen, Q. Experimental Transmission of the Yeast, Metschnikowia bicuspidata, in the Chinese Mitten Crab, Eriocheir sinensis. J. Fungi 2022, 8, 210. https://doi.org/10.3390/jof8020210
Jiang H, Bao J, Cao G, Xing Y, Feng C, Hu Q, Li X, Chen Q. Experimental Transmission of the Yeast, Metschnikowia bicuspidata, in the Chinese Mitten Crab, Eriocheir sinensis. Journal of Fungi. 2022; 8(2):210. https://doi.org/10.3390/jof8020210
Chicago/Turabian StyleJiang, Hongbo, Jie Bao, Gangnan Cao, Yuenan Xing, Chengcheng Feng, Qingbiao Hu, Xiaodong Li, and Qijun Chen. 2022. "Experimental Transmission of the Yeast, Metschnikowia bicuspidata, in the Chinese Mitten Crab, Eriocheir sinensis" Journal of Fungi 8, no. 2: 210. https://doi.org/10.3390/jof8020210
APA StyleJiang, H., Bao, J., Cao, G., Xing, Y., Feng, C., Hu, Q., Li, X., & Chen, Q. (2022). Experimental Transmission of the Yeast, Metschnikowia bicuspidata, in the Chinese Mitten Crab, Eriocheir sinensis. Journal of Fungi, 8(2), 210. https://doi.org/10.3390/jof8020210