
Morphological and Molecular Identification of Plant Pathogenic Fungi Associated with Dirty Panicle Disease in Coconuts (Cocos nucifera) in Thailand

 ,
,  ,
,  ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Observation, Symptom Recognition, and Sample Collection
2.2. Fungal Isolation
2.3. Pathogenicity Test
2.4. Morphological Identification
2.5. Molecular Identification
3. Results
3.1. Symptom Recognition
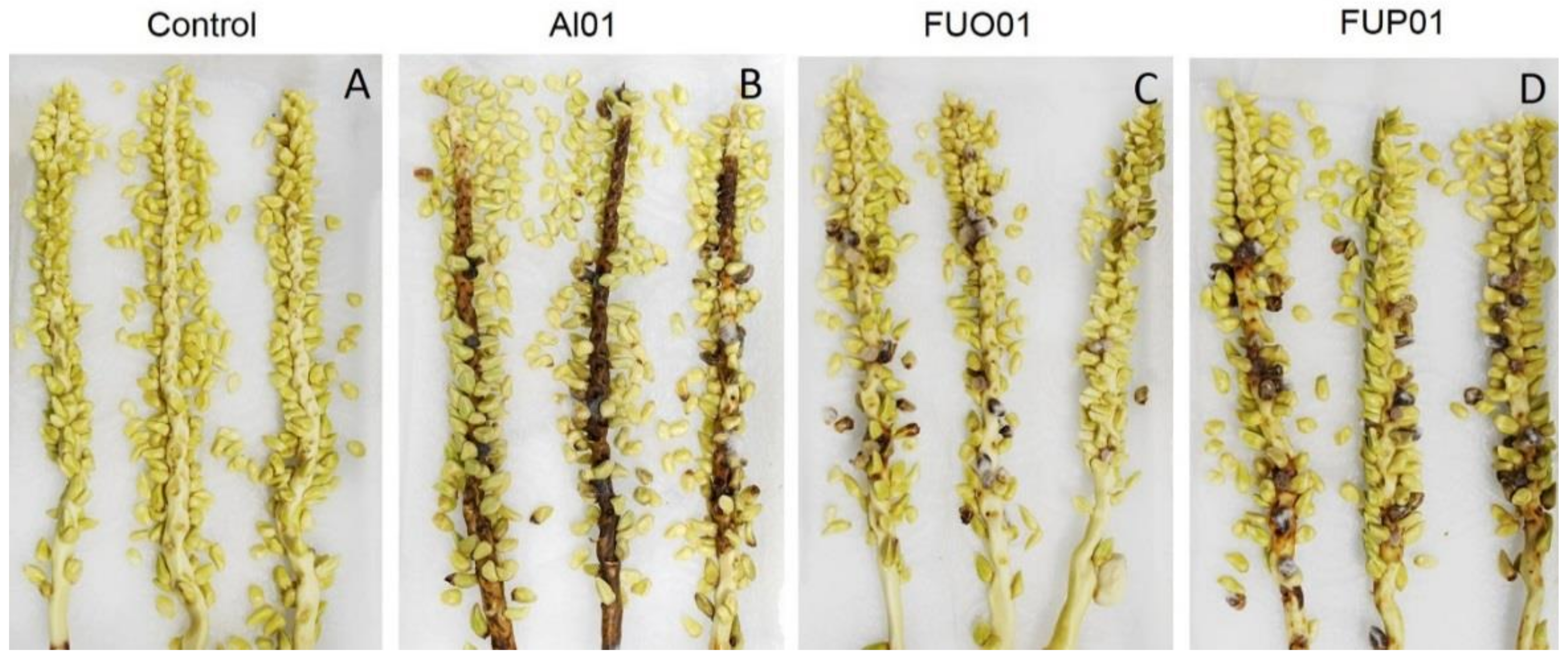
3.2. Pathogenicity Test
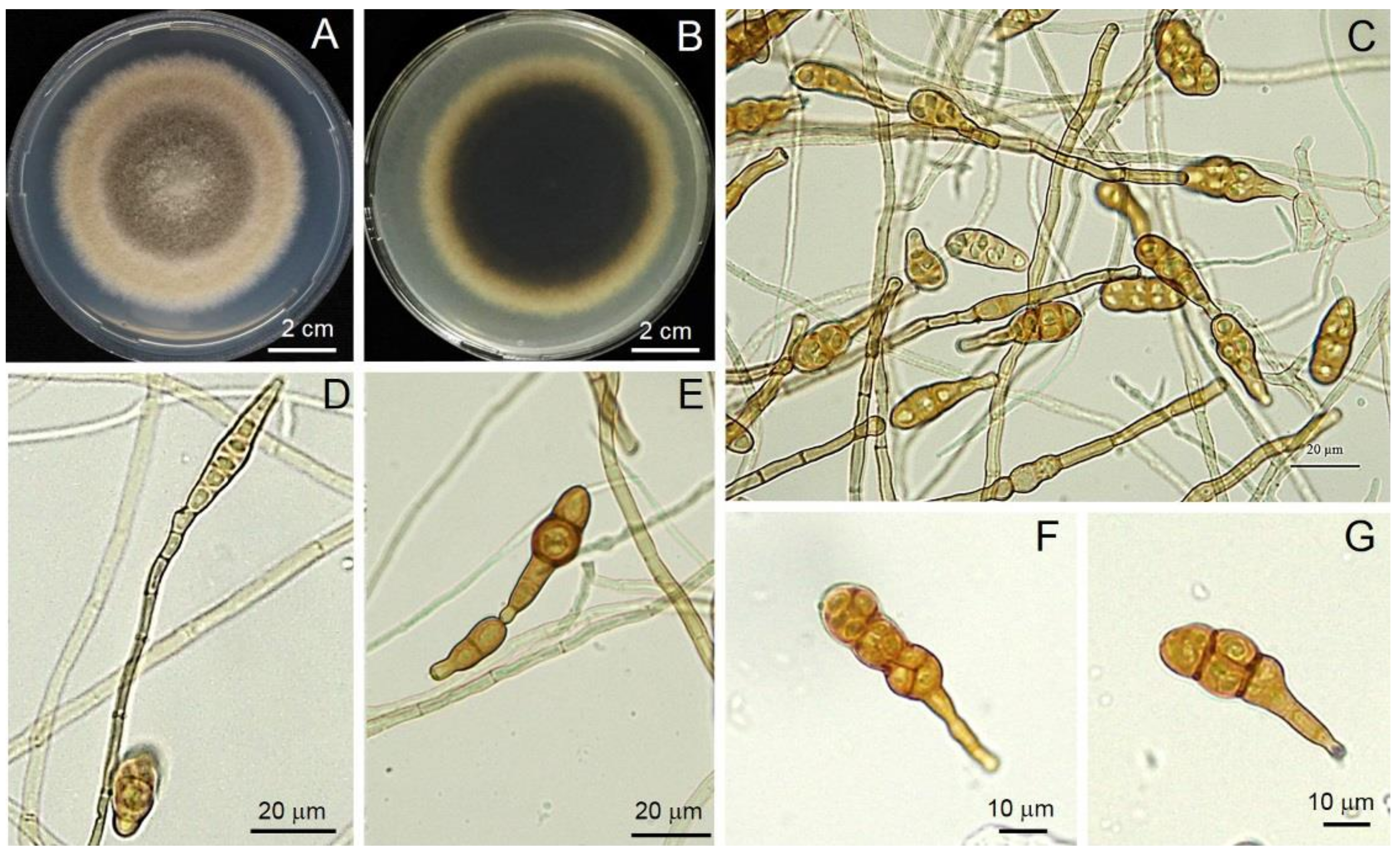
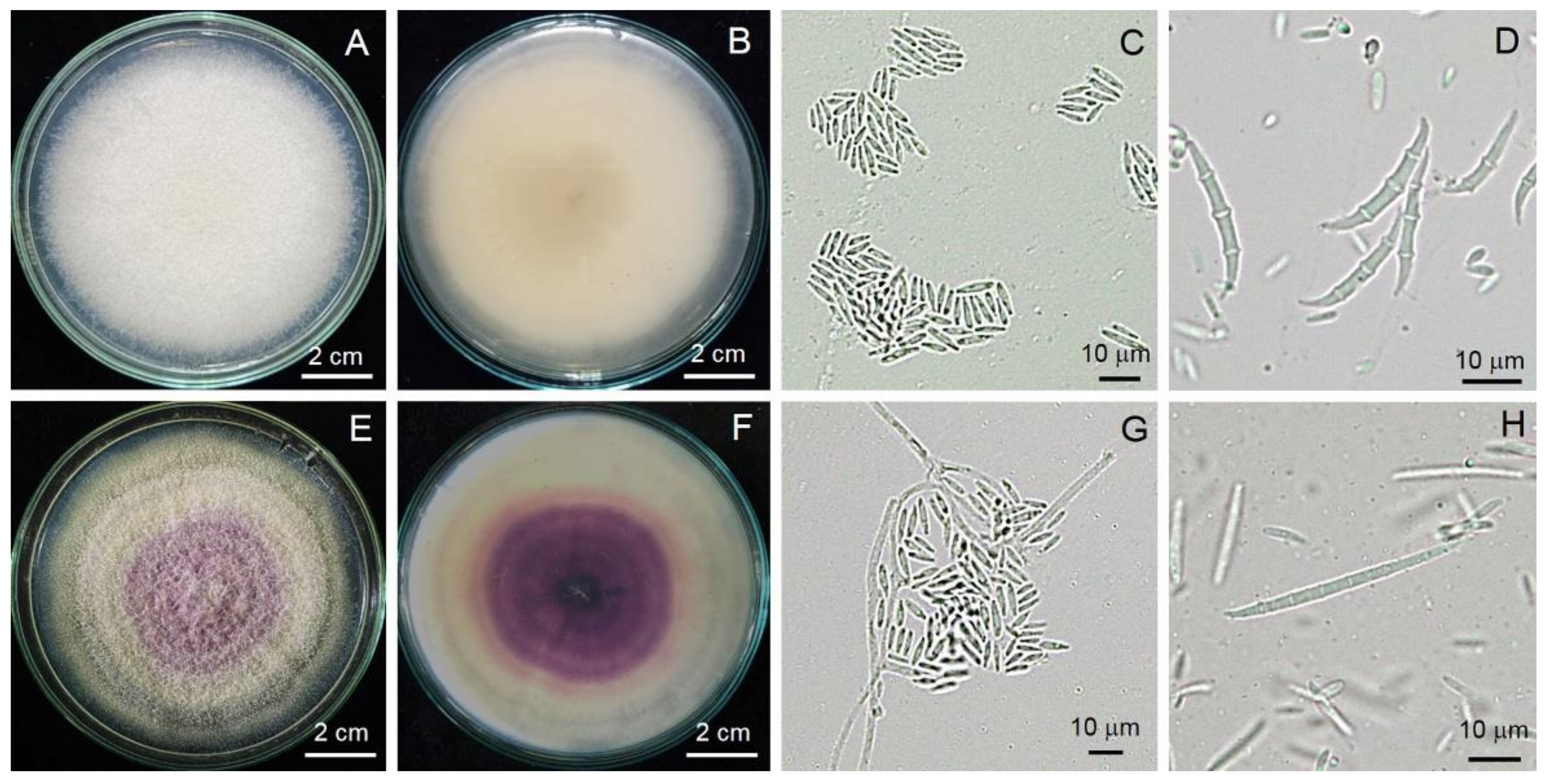
3.3. Morphological Identification
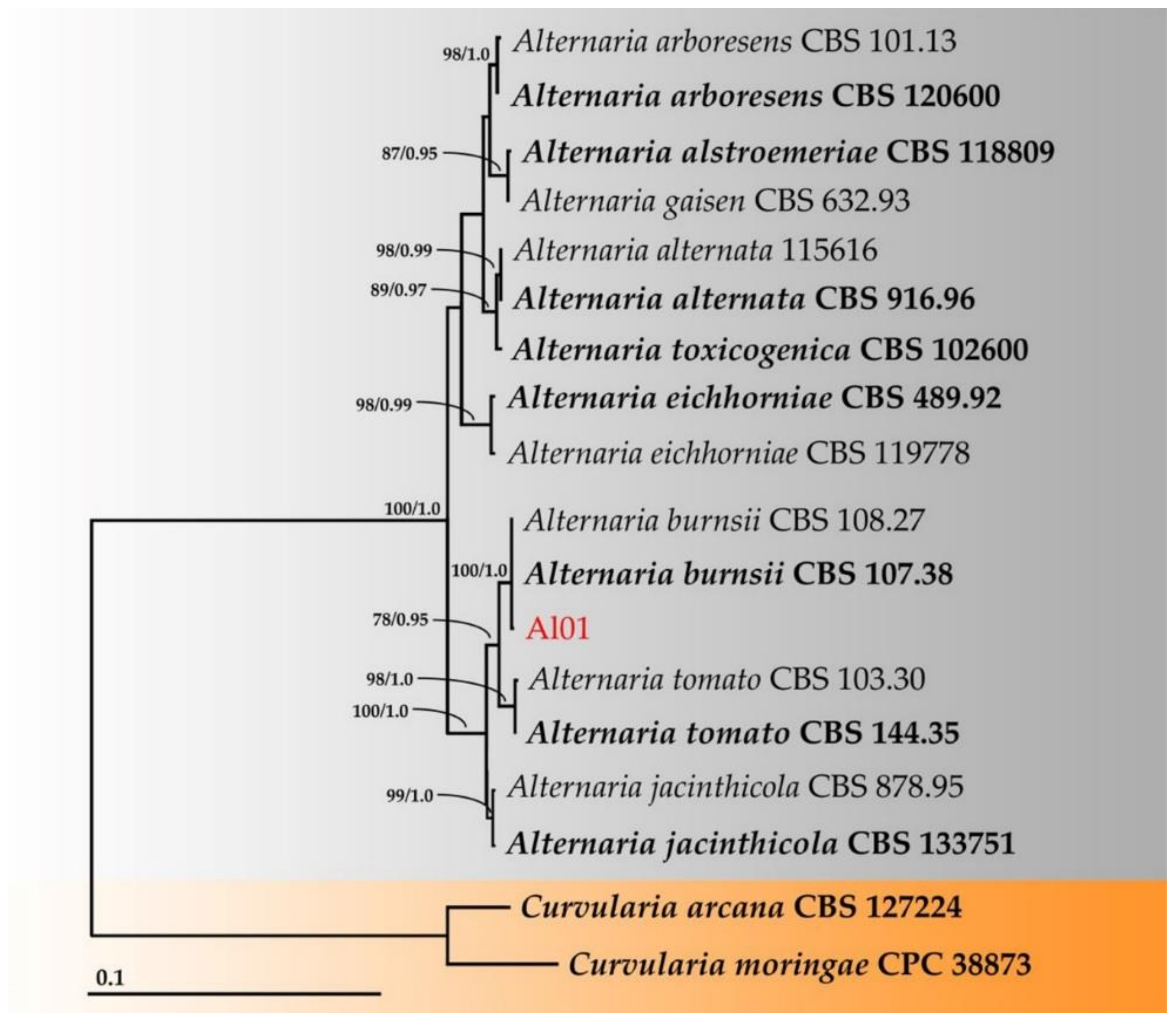
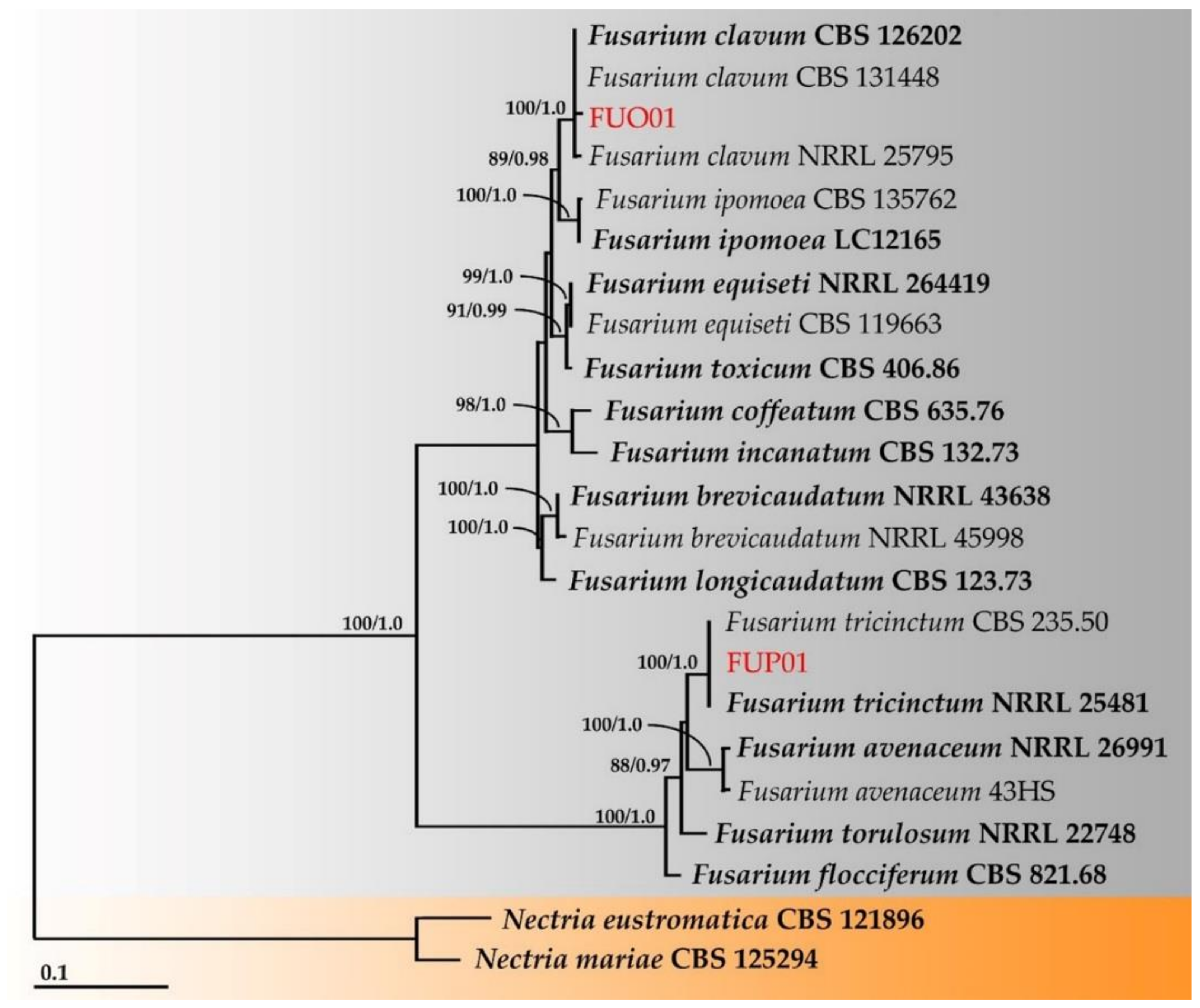
3.4. Molecular Identification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adkins, S.; Foale, M.; Samosir, Y. Coconut revival-new possibilities for the ‘tree of life’. In International Coconut Forum, ACIAR Proceedings No. 125; ACT: Australian Centre for International Agricultural Research: Canberra, Australia, 2006; pp. 1–103. [Google Scholar]
- FAO. FAO Statistics; Food and Agriculture Organization of the United Nations: Rome, Italy, 2014; Volume 2014. [Google Scholar]
- Bourke, R.M.; Harwood, T. Food and Agriculture in Papua New Guinea; ANU E Press; The Australian National University: Canberra, Australia, 2009. [Google Scholar]
- Yu, F.-Y.; Niu, X.-Q.; Tang, Q.-H.; Zhu, H.; Song, W.-W.; Qin, W.-Q.; Lin, C.-H. First report of stem bleeding in coconut caused by Ceratocyctis paradoxa in Hainan, China. Plant Dis. 2012, 96, 290. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.-Q.; Zhu, H.; Yu, F.-Y.; Tang, Q.-H.; Song, W.-W.; Qin, W.-Q. First report of Pestalotiopsis menezesiana causing leaf blight of coconut in Hainan, China. Plant Dis. 2015, 99, 554. [Google Scholar] [CrossRef]
- Venugopal, S.; Mohanan, R.C. Role of fungi in fruit rot and immature nut fall of coconut. Cord 2006, 22, 33–40. [Google Scholar]
- Niu, X.-Q.; Yu, F.-Y.; Zhu, H.; Qin, W.-Q. First report of leaf spot disease in coconut seedling caused by Bipolaris setariae in China. Plant Dis. 2014, 98, 1742. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Niu, X.L. First report of Lasiodiplodia theobromae causing postharvest stem end rot on coconut in China. Plant Dis. 2019, 103, 1420. [Google Scholar] [CrossRef]
- Riangwong, K.; Wanchana, S.; Aesomnuk, W.; Saensuk, C.; Nubankoh, P.; Ruanjaichon, V.; Kraithong, T.; Toojinda, T.; Vanavichit, A.; Arikit, S. Mining and validation of novel genotyping-by-sequencing (GBS)-based simple sequence repeats (SSRs) and their application for the estimation of the genetic diversity and population structure of coconuts (Cocos nucifera L.) in Thailand. Hortic. Res. 2020, 7, 156. [Google Scholar] [CrossRef]
- Saensuk, C.; Wanchana, S.; Choowongkomon, K.; Wongpornchai, S.; Kraithong, T.; Imsabai, W.; Chaichoompu, E.; Ruanjaichon, V.; Toojinda, T.; Vanavichit, A.; et al. De novo transcriptome assembly and identification of the gene conferring a “pandan-like” aroma in coconut (Cocos nucifera L.). Plant Sci. 2016, 252, 324–334. [Google Scholar] [CrossRef]
- Dumhai, R.; Wanchana, S.; Saensuk, C.; Choowongkomon, K.; Mahatheeranont, S.; Kraithong, T.; Toojinda, T.; Vanavichit, A.; Arikit, S. Discovery of a novel CnAMADH2 allele associated with higher levels of 2-acetyl-1-pyrroline (2AP) in yellow dwarf coconut (Cocos nucifera L.). Sci. Hortic. 2019, 243, 490–497. [Google Scholar] [CrossRef]
- Wonglom, P.; Thithuan, N.; Bunjongsiri, P.; Sunpapao, A. Plant-parasitic algae (Cephaleuros spp.) in Thailand, including four new records. Pac. Sci. 2018, 72, 363–371. [Google Scholar] [CrossRef]
- Thithuan, N.; Bunjonsiri, P.; Sunpapao, A. Morphology and behavior of gametes and zoospores from the plant-parasitic green algae, Cephaleuros (Chlorophyta, Ulvophyceae). Pac. Sci. 2019, 73, 403–410. [Google Scholar] [CrossRef]
- Daengsuwan, W.; Wonglom, P.; Sunpapao, A. First report of Lasiodiplodia theobromae causing spadix rot in Anthurium andraeanum. J. Phytopathol. 2020, 168, 129–133. [Google Scholar] [CrossRef]
- Daengsuwan, W.; Wonglom, P.; Arikit, S.; Sunpapao, A. Morphological and molecular identification of the fungus Neopestalotiopsis clavispora associated with flower blight on Anthurium andraeanum in Thailand. Hortic. Plant J. 2021, 7, 573–578. [Google Scholar] [CrossRef]
- Pornsuriya, C.; Chairin, T.; Thaochan, N.; Sunpapao, A. Identification and characterization of Neopestalotiopsis fungi associated with a novel leaf fall disease of rubber trees (Hevea brasiliensis) in Thailand. J. Phytopathol. 2020, 168, 416–427. [Google Scholar] [CrossRef]
- Ruangwong, O.-U.; Kunasakdakul, K.; Wonglom, P.; Dy, K.S.; Sunpapao, A. Morphological and molecular studies of a rare mucoralean species causing flower rot in Hylocereus polyrhizus. J. Phytopathol. 2022, 170, 217–220. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Tayler, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenies. In PCR Protocols: A Guide to Methods and Applications; Innis, A.M., Gelfelfard, D.H., Snindky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Larsson, K.; Ansell, R.; Eriksson, P.; Adler, L. A gene encoding snglycerol 3-phosphate dehydrogenase (NAD+) complements an osmosensitive mutant of Saccharomyces cerevisiae. Mol. Microbiol. 1993, 10, 1101–1111. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [Green Version]
- Hall, T. Bioedit Version 6.0.7. 2004. Available online: http://www.mbio.ncsu.edu/bioedit/bioedit.html (accessed on 20 November 2021).
- Felsenstein, J. Confidence intervals on phylogenetics: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the cipres science gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE: Manhattan, NY, USA; pp. 1–8. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree Tree Figure Drawing Tool Version 131, Institute of Evolutionary Biology, University of Edinburgh. Available online: http://treebioedacuk/software/figtree (accessed on 5 February 2022).
- Paul, N.C.; Deng, J.X.; Lee, H.B.; Yu, S.H. Characterization and pathogenicity of Alternaria burnsii from seeds of Cucurbita maxima (Cucurbitaceae) in Bangladesh. Mycobiology 2015, 43, 384–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilardi, G.; Matic, S.; Guarnaccia, V.; Garibaldi, A.; Gullino, M.L. First report of Fusarium clavum causing leaf spot and fruit rot on tomato in Italy. Plant Dis. 2021, 105, 2250. [Google Scholar] [CrossRef]
- Garibaldi, A.; Tabone, G.; Guarnaccia, V.; Luongo, I.; Gullino, M.L. First report of Fusarium tricinctum species complex causing leaf spot on Hosta fortunei in Italy. J. Plant Pathol. 2021, 103, 1071. [Google Scholar] [CrossRef]
- Al-Nadabi, H.H.; Maharachchikumbura, S.S.N.; Agrama, H.; Al-Azri, M.; Nasehi, A.; Al-Sadi, A.M. Molecular characterization and pathogenicity of Alternaria species on wheat and date palms in Oman. Eur. J. Plant Pathol. 2018, 152, 577–588. [Google Scholar] [CrossRef]
- Kosiak, B.; Torp, M.; Skjerve, E.; Andersen, B. Alternaria and Fusarium in Norwegian grains of reduced quality—A matched pair sample study. Int. J. Food Microbiol. 2004, 93, 51–62. [Google Scholar] [CrossRef]
- Wonglom, P.; Sunpapao, A. Fusarium incarnatum is associated with postharvest fruit rot of muskmelon (Cucumis melo). J. Phytopathol. 2020, 168, 204–210. [Google Scholar] [CrossRef]
- Bajwa, R.; Mukhtar, I.; Mushtaq, S. New report of Alternaria alternata causing leaf spot of Aloe vera in Pakistan. Can. J. Plant Pathol. 2010, 32, 4. [Google Scholar] [CrossRef]
- Hollaway, G.J.; Evans, M.L.; Wallwork, H.; Dyson, C.B.; McKay, A.C. Yield loss in cereals, caused by Fusarium culmorum and F. pseudograminearum, is related to fungal DNA in soil prior to planting, rainfall, and cereal type. Plant Dis. 2013, 97, 977–982. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.X.; Xu, W.X. First report of Alternaria alternata causing leaf spots of tea (Camellia sinensis) in China. Plant Dis. 2014, 98, 697. [Google Scholar] [CrossRef]
- Aloi, F.; Zamora-Ballesteros, C.; Martín-García, J.; Diez, J.J.; Cacciola, S.O. Co-infections by Fusarium circinatum and Phytophthora spp. on Pinus radiata: Complex phenotypic and molecular interactions. Plants 2021, 10, 1976. [Google Scholar] [CrossRef] [PubMed]
- Gavrilova, O.; Gannibal, P.; Gagkaeva, T. Fusarium and Alternaria fungi in grain of oats grown in the north-western Russia regarding cultivar specificity. Sel’skokhozyaistvennaya Biol. 2016, 51, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Schiro, G.; Verch, G.; Grimm, V.; Müller, M.E.H. Alternaria and Fusarium fungi: Differences in distribution and spore deposition in a topographically heterogeneous wheat field. J. Fungi 2018, 4, 63. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, X.; Du, T.; Xue, L.; Jin, L. Disease survey and pathogen identification on Notopterygium incisum in Gansu Province. China J. Chin. Mater. Med. 2009, 34, 1898–1901. [Google Scholar]
- Kamkar, B.; Koocheki, A.; Mahallati, M.N.; da Silva, J.A.T.; Moghaddam, P.R.; Kafi, M. Fungal diseases and inappropriate sowing dates, the most important reducing factors in cumin fields of Iran, a case study in Khorasan provinces. Crop Prot. 2011, 30, 208–215. [Google Scholar] [CrossRef]
- Matić, S.; Tabone, G.; Guarnaccia, V.; Gullino, M.L.; Garibaldi, A. Emerging leafy vegetable crop diseases caused by the Fusarium incarnatum-equiseti species complex. Phytopathol. Mediterr. 2020, 59, 303–317. [Google Scholar]
- Manganiello, G.; Traversari, S.; Nesi, B.; Cacini, S.; Pane, C. Rose: A new host plant of Fusarium clavum (F. incarnatum-equiseti species complex 5) causing brown spot of petals. Crop Prot. 2021, 146, 105675. [Google Scholar] [CrossRef]
- Castañares, E.; Stenglein, S.A.; Dinolfo, M.I.; Moreno, M.V. Fusarium tricinctum associated with head blight on wheat in argentina. Plant Dis. 2011, 95, 496. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, J.; Ma, L.; Li, E.; Ji, B.; Sun, C.; Zhou, J.; Xu, B. First report of a new disease caused by Fusarium tricinctum on apple tree in China. Plant Dis. 2021. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sunpapao, A.; Suwannarach, N.; Kumla, J.; Dumhai, R.; Riangwong, K.; Sanguansub, S.; Wanchana, S.; Arikit, S. Morphological and Molecular Identification of Plant Pathogenic Fungi Associated with Dirty Panicle Disease in Coconuts (Cocos nucifera) in Thailand. J. Fungi 2022, 8, 335. https://doi.org/10.3390/jof8040335
Sunpapao A, Suwannarach N, Kumla J, Dumhai R, Riangwong K, Sanguansub S, Wanchana S, Arikit S. Morphological and Molecular Identification of Plant Pathogenic Fungi Associated with Dirty Panicle Disease in Coconuts (Cocos nucifera) in Thailand. Journal of Fungi. 2022; 8(4):335. https://doi.org/10.3390/jof8040335
Chicago/Turabian StyleSunpapao, Anurag, Nakarin Suwannarach, Jaturong Kumla, Reajina Dumhai, Kanamon Riangwong, Sunisa Sanguansub, Samart Wanchana, and Siwaret Arikit. 2022. "Morphological and Molecular Identification of Plant Pathogenic Fungi Associated with Dirty Panicle Disease in Coconuts (Cocos nucifera) in Thailand" Journal of Fungi 8, no. 4: 335. https://doi.org/10.3390/jof8040335
APA StyleSunpapao, A., Suwannarach, N., Kumla, J., Dumhai, R., Riangwong, K., Sanguansub, S., Wanchana, S., & Arikit, S. (2022). Morphological and Molecular Identification of Plant Pathogenic Fungi Associated with Dirty Panicle Disease in Coconuts (Cocos nucifera) in Thailand. Journal of Fungi, 8(4), 335. https://doi.org/10.3390/jof8040335