
Integration of In Silico and In Vitro Analysis of Gliotoxin Production Reveals a Narrow Range of Producing Fungal Species

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioinformatic Study
2.1.1. Search for A. fumigatus Af293 GliP Homologous Proteins in Other Species
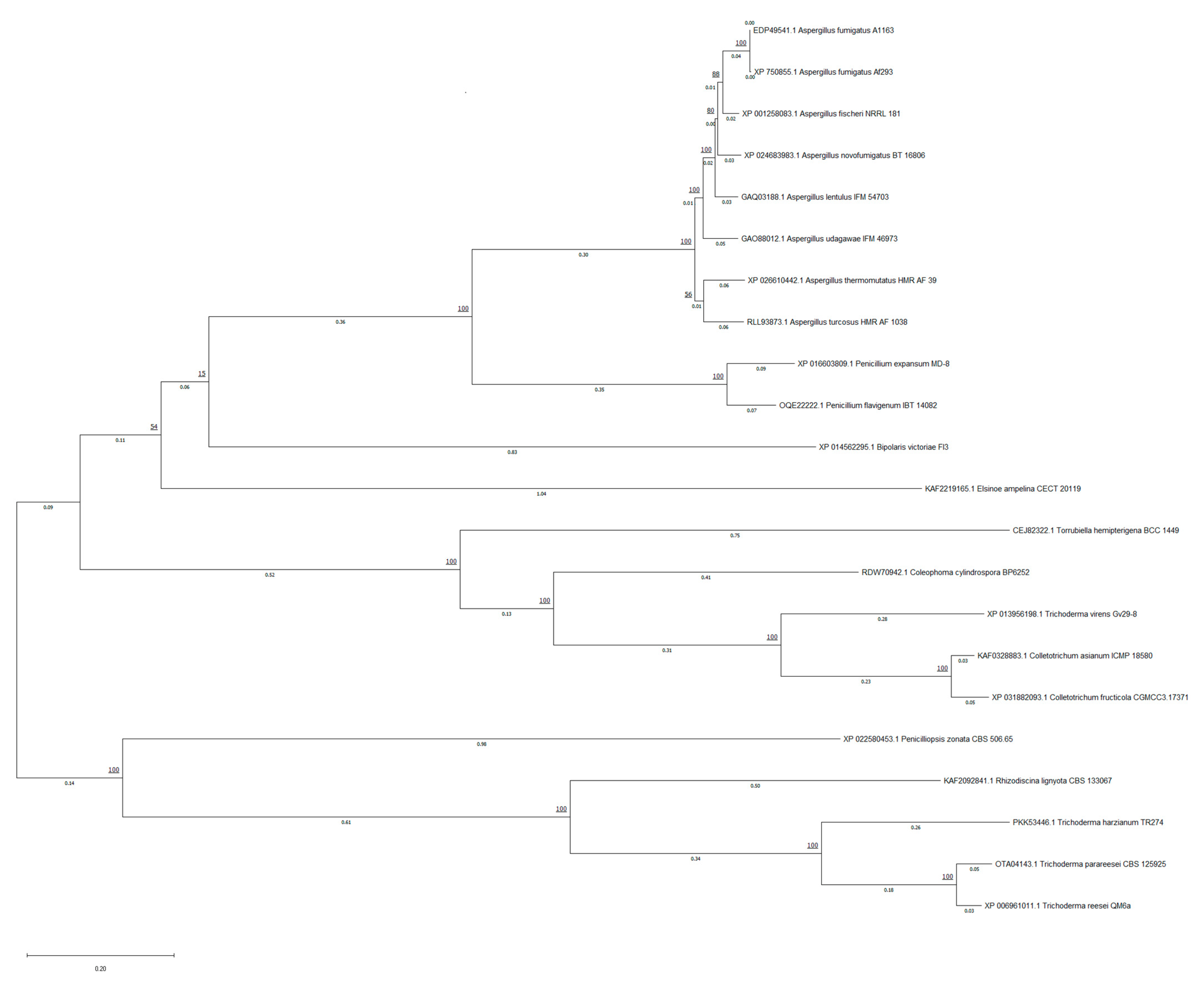
2.1.2. Phylogenetic Tree
2.1.3. Search for Possible Clusters of Secondary Metabolites in Various Strains Using antiSMASH
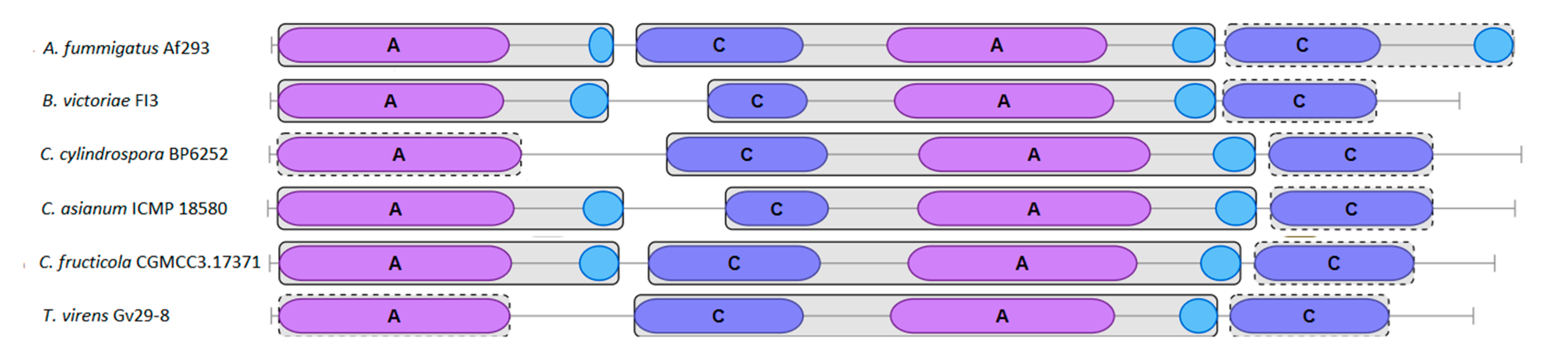
Identification of the Rest of gli Genes Surrounding the Selected NRPSs
Synteny and Gene Location Inside the Clusters
Homology between A. fumigatus Af293 gli Genes and Their Counterparts in the Other Fungi Studied
2.2. Analysis and Quantification of GT Production by Several Fungal Strains
2.2.1. Fungal Strains and Culturing Conditions
2.2.2. GT and bmGT Chemical Extraction
2.2.3. HPLC Analysis and Quantification
3. Results
3.1. Bioinformatic Study
3.1.1. Search for A. fumigatus Af293 GliP Homologous Proteins in Other Species
3.1.2. Phylogenetic Tree
3.1.3. Search for Possible Clusters of Secondary Metabolites in Various Strains Using antiSMASH
3.2. Analysis and Quantification of GT Production by Several Fungal Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Waring, P.; Beaver, J. Gliotoxin and related epipolythiodioxopiperazines. Gen. Pharmacol. 1996, 27, 1311–1316. [Google Scholar] [CrossRef]
- Gardiner, D.M.; Waring, P.; Howlett, B.J. The epipolythiodioxopiperazine (ETP) class of fungal toxins: Distribution, mode of action, functions and biosynthesis. Microbiology 2005, 151, 1021–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon-Chung, K.J.; Sugui, J.A. What do we know about the role of gliotoxin in the pathobiology of Aspergillus fumigatus? Med. Mycol. J. 2009, 47, S97–S103. [Google Scholar] [CrossRef] [Green Version]
- Stanley, S.A.; Grant, S.S.; Kawate, T.; Iwase, N.; Shimizu, M.; Wivagg, C.; Silvis, M.; Kazyanskaya, E.; Aquadro, J.; Golas, A.; et al. Identification of novel inhibitors of M. tuberculosis growth using whole cell based high-throughput screening. ACS Chem. Biol. 2012, 7, 1377–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tompsett, R.; McDermott, W.; Kidd, J.G. Tuberculostatic activity of blood and urine from animals given gliotoxin. J. Immunol. Res. 1950, 65, 59–63. [Google Scholar]
- Esteban, P.; Redrado, S.; Comas, L.; Domingo, M.P.; Millán-Lou, M.I.; Seral, C.; Algarate, S.; Lopez, C.; Rezusta, A.; Pardo, J.; et al. In Vitro and In Vivo Antibacterial Activity of Gliotoxin Alone and in Combination with Antibiotics against Staphylococcus aureus. Toxins 2021, 13, 85. [Google Scholar] [CrossRef]
- Coleman, J.J.; Ghosh, S.; Okoli, I.; Mylonakis, E. Antifungal activity of microbial secondary metabolites. PLoS ONE 2011, 6, e25321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatabu, T.; Hagiwara, M.; Taguchi, N.; Kiyozawa, M.; Suzuki, M.; Kano, S.; Sato, K. Plasmodium falciparum: The fungal metabolite gliotoxin inhibits proteasome proteolytic activity and exerts a plasmodicidal effect on P. falciparum. Exp. Parasitol. 2006, 112, 179–183. [Google Scholar] [CrossRef]
- Aljofan, M.; Sganga, M.L.; Lo, M.K.; Rootes, C.L.; Porotto, M.; Meyer, A.G.; Saubern, S.; Moscona, A.; Mungall, B.A. Antiviral activity of gliotoxin, gentian violet and brilliant green against Nipah and Hendra virus in vitro. Virol. J. 2009, 6, 187. [Google Scholar] [CrossRef] [Green Version]
- McDougall, J.K. Antiviral action of gliotoxin. Arch. Gesamte Virusforsch. 1969, 27, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Rightsel, W.A.; Schneider, H.G.; Schneider, H.G.; Sloan, B.J.; Graf, P.R.; Miller, F.A.; Bartz, O.R.; Ehrlich, J.; Dixon, G.J. Antiviral activity of gliotoxin and gliotoxin acetate. Nature 1964, 204, 1333–1334. [Google Scholar] [CrossRef]
- Larin, N.M.; Copping, M.P.; Herbst-Laier, R.H.; Roberts, B.; Wenham, R.B. Antiviral activity of gliotoxin. Chemotherapy 1965, 10, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Sugui, J.A.; Pardo, J.; Chang, Y.C.; Zarember, K.A.; Nardone, G.; Galvez, E.M.; Müllbacher, A.; Gallin, J.I.; Simon, M.M.; Kwon-Chung, K.J. Gliotoxin is a virulence factor of Aspergillus fumigatus: gliP deletion attenuates virulence in mice immunosuppressed with hydrocortisone. Eukaryot. Cell 2007, 6, 1562–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spikes, S.; Xu, R.; Nguyen, C.K.; Chamilos, G.; Kontoyiannis, D.P.; Jacobson, R.H.; Ejzykowicz, D.E.; Chiang, L.Y.; Filler, S.G.; May, G.S. Gliotoxin Production in Aspergillus fumigatus Contributes to Host-Specific Differences in Virulence. J. Infect. Dis. 2008, 197, 479–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bok, J.W.; Chung, D.; Balajee, S.A.; Marr, K.A.; Andes, D.; Nielsen, K.F.; Frisvad, J.C.; Kirby, K.A.; Keller, N.P. GliZ, a transcriptional regulator of gliotoxin biosynthesis, contributes to Aspergillus fumigatus virulence. Infect. Immun. 2006, 74, 6761–6768. [Google Scholar] [CrossRef] [Green Version]
- Sugui, J.A.; Chung, D.; Balajee, S.A.; Marr, K.A.; Andes, D.; Nielsen, K.F.; Frisvad, J.C.; Kirby, K.A.; Keller, N.P. Host immune status-specific production of gliotoxin and bis-methyl-gliotoxin during invasive aspergillosis in mice. Sci. Rep. 2017, 7, 10977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichner, R.D.; Al Salami, M.; Wood, P.R.; Müllbacher, A. The effect of gliotoxin upon macrophage function. Int. J. Immunopharmacol. 1986, 8, 789–797. [Google Scholar] [CrossRef]
- Müllbacher, A.; Eichner, R.D.; Eichner, R.D. Immunosuppression in vitro by a metabolite of a human pathogenic fungus. Proc. Natl. Acad. Sci. USA 1984, 81, 3835–3837. [Google Scholar] [CrossRef] [Green Version]
- Pahl, H.L.; Krauss, B.; Schulze-Osthoff, K.; Decker, T.; Traenckner, E.B.; Vogt, M.; Myers, C.; Parks, T.; Warring, P.; Mühlbacher, A.; et al. The immunosuppressive fungal metabolite gliotoxin specifically inhibits transcription factor NF-kappaB. J. Exp. Med. 1996, 183, 1829–1840. [Google Scholar] [CrossRef] [PubMed]
- Waring, P.; Eichner, R.D.; Müllbacher, A.; Sjaarda, A. Gliotoxin induces apoptosis in macrophages unrelated to its antiphagocytic properties. Int. J. Biol. Chem. 1988, 263, 18493–18499. [Google Scholar] [CrossRef]
- Stanzani, M.; Orciuolo, E.; Lewis, R.; Kontoyiannis, D.P.; Martins, S.L.; St John, L.S.; Komanduri, K.V. Aspergillus fumigatus suppresses the human cellular immune response via gliotoxin-mediated apoptosis of monocytes. Blood 2005, 105, 2258–2265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niide, O.; Suzuki, Y.; Yoshimaru, T.; Inoue, T.; Takayama, T.; Ra, C. Fungal metabolite gliotoxin blocks mast cell activation by a calcium- and superoxide-dependent mechanism: Implications for immunosuppressive activities. J. Clin. Immunol. 2006, 118, 108–116. [Google Scholar] [CrossRef]
- Arias, M.; Santiago, L.; Vidal-García, M.; Redrado, S.; Lanuza, P.; Comas, L.; Domingo, M.P.; Rezusta, A.; Gálvez, E.M. Preparations for Invasion: Modulation of Host Lung Immunity During Pulmonary Aspergillosis by Gliotoxin and Other Fungal Secondary Metabolites. Front. Immunol. 2018, 9, 2549. [Google Scholar] [CrossRef] [PubMed]
- Piva, T.J. Gliotoxin induces apoptosis in mouse L929 fibroblast cells. Int. J. Biochem. Mol. Biol. 1994, 33, 411–419. [Google Scholar]
- Kweon, Y.O.; Paik, Y.H.; Schnabl, B.; Qian, T.; Lemasters, J.J.; Brenner, D.A. Gliotoxin-mediated apoptosis of activated human hepatic stellate cells. J. Hepatol. 2003, 39, 38–46. [Google Scholar] [CrossRef]
- Nguyen, V.T.; Lee, J.S.; Qian, Z.J.; Li, Y.X.; Kim, K.N.; Heo, S.J.; Jeon, Y.J.; Park, W.S.; Choi, I.W.; Je, J.Y.; et al. Gliotoxin isolated from marine fungus Aspergillus sp. induces apoptosis of human cervical cancer and chondrosarcoma cells. Mar. Drugs 2013, 24, 69–87. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, C.; Huang, C.; Lin, M.; Wang, Z.; Liang, W.; Iwamoto, A.; Yang, X.; Liu, H. Gliotoxin Inhibits Proliferation and Induces Apoptosis in Colorectal Cancer Cells. Mar. Drugs 2015, 13, 6259–6273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comas, L.; Polo, E.; Domingo, M.P.; Hernández, Y.; Arias, M.; Esteban, P.; Martínez-Lostao, L.; Pardo, J.; Martínez de la Fuente, J.; Gálvez, E.M. Intracellular Delivery of Biologically-Active Fungal Metabolite Gliotoxin Using Magnetic Nanoparticles. Materials 2019, 12, 1092. [Google Scholar] [CrossRef] [Green Version]
- Ráduly, Z.; Szabó, L.; Madar, A.; Pócsi, I.; Csernoch, L. Toxicological and Medical Aspects of Aspergillus-Derived Mycotoxins Entering the Feed and Food Chain. Front. Microbiol. 2020, 10, 2908. [Google Scholar] [CrossRef] [Green Version]
- Gallo, A.; Giuberti, G.; Frisvad, J.C.; Bertuzzi, T.; Nielsen, K.F. Review on Mycotoxin Issues in Ruminants: Occurrence in Forages, Effects of Mycotoxin Ingestion on Health Status and Animal Performance and Practical Strategies to Counteract Their Negative Effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef]
- Novak, B.; Rainer, V.; Sulyok, M.; Haltrich, D.; Schatzmayr, G.; Mayer, E. Twenty-Eight Fungal Secondary Metabolites Detected in Pig Feed Samples: Their Occurrence, Relevance and Cytotoxic Effects In Vitro. Toxins 2019, 11, 537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardiner, D.M.; Howlett, B.J. Bioinformatic and expression analysis of the putative gliotoxin biosynthetic gene cluster of Aspergillus fumigatus. FEMS Microbiol. Lett. 2005, 248, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Balibar, C.J.; Walsh, C.T. GliP, a multimodular nonribosomal peptide synthetase in Aspergillus fumigatus, makes the diketopiperazine scaffold of gliotoxin. Biochemistry 2006, 45, 15029–15038. [Google Scholar] [CrossRef] [PubMed]
- Dolan, S.K.; O’Keeffe, G.; Jones, G.W.; Doyle, S. Resistance Is Not Futile: Gliotoxin Biosynthesis, Functionality and Utility, in Trends in Microbiology; Elsevier Ltd.: Amsterdam, The Netherlands, 2015; pp. 419–428. [Google Scholar]
- Chang, S.L.; Chiang, Y.M.; Yeh, H.H.; Wu, T.K.; Wang, C.C. Reconstitution of the early steps of gliotoxin biosynthesis in Aspergillus nidulans reveals the role of the monooxygenase GliC. Bioorg. Med. Chem. Lett. 2013, 23, 2155–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, C.; Carberry, S.; Schrettl, M.; Singh, I.; Stephens, J.C.; Barry, S.M.; Kavanagh, K.; Challis, G.L.; Brougham, D.; Doyle, S. The role of glutathione S-transferase GliG in gliotoxin biosynthesis in Aspergillus fumigatus. Chem. Biol. 2011, 18, 542–552. [Google Scholar] [CrossRef] [Green Version]
- Scharf, D.H.; Chankhamjon, P.; Scherlach, K.; Heinekamp, T.; Willing, K.; Brakhage, A.A.; Hertweck, C. Epidithiodiketopiperazine biosynthesis: A four-enzyme cascade converts glutathione conjugates into transannular disulfide bridges. Angew. Chem. Int. Ed. 2013, 11, 11092–11095. [Google Scholar] [CrossRef] [PubMed]
- Schrettl, M.; Carberry, S.; Kavanagh, K.; Haas, H.; Jones, G.W.; O’Brien, J.; Nolan, A.; Stephens, J.; Fenelon, O.; Doyle, S. Self-protection against gliotoxin—A component of the gliotoxin biosynthetic cluster, GliT, completely protects Aspergillus fumigatus against exogenous gliotoxin. PLoS Pathog. 2010, 10, e1000952. [Google Scholar] [CrossRef] [PubMed]
- Scharf, D.H.; Remme, N.; Heinekamp, T.; Hortschansky, P.; Brakhage, A.A.; Hertweck, C. Transannular disulfide formation in gliotoxin biosynthesis and its role in self-resistance of the human pathogen Aspergillus fumigatus. J. Am. Chem. Soc. 2010, 132, 10136–10141. [Google Scholar] [CrossRef] [PubMed]
- Ries, L.N.A.; Pardeshi, L.; Dong, Z.; Tan, K.; Steenwyk, J.L.; Colabardini, A.C.; Ferreira Filho, J.A.; de Castro, P.A.; Silva, L.P.; Preite, N.W.; et al. The Aspergillus fumigatus transcription factor RglT is important for gliotoxin biosynthesis and self-protection, and virulence. PLoS Pathog. 2020, 15, e1008645. [Google Scholar] [CrossRef]
- Dolan, S.K.; Owens, R.A.; O’Keeffe, G.; Hammel, S.; Fitzpatrick, D.A.; Jones, G.W.; Doyle, S. Regulation of Nonribosomal Peptide Synthesis: Bis-Thiomethylation Attenuates Gliotoxin Biosynthesis in Aspergillus fumigatus. Chem. Biol. 2014, 21, 999–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scharf, D.H.; Groll, M.; Habel, A.; Heinekamp, T.; Hertweck, C.; Brakhage, A.A.; Huber, E.M. Flavoenzyme-catalyzed formation of disulfide bonds in natural products. Angew. Chem. Int. Ed. 2014, 53, 2221–2224. [Google Scholar] [CrossRef]
- Cramer, R.A.; Gamcsik, M.P.; Brooking, R.M.; Najvar, L.K.; Kirkpatrick, W.R.; Patterson, T.F.; Balibar, C.J.; Graybill, J.R.; Perfect, J.R.; Abraham, S.N.; et al. Disruption of a nonribosomal peptide synthetase in Aspergillus fumigatus eliminates gliotoxin production. Eukaryot. Cell 2006, 5, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.N.; Toyotome, T.; Muraosa, Y.; Watanabe, A.; Wuren, T.; Bunsupa, S.; Aoyagi, K.; Yamazaki, M.; Takino, M.; Kamei, K. GliA in Aspergillus fumigatus is required for its tolerance to gliotoxin and affects the amount of extracellular and intracellular gliotoxin. Med. Mycol. J. 2014, 52, 506–518. [Google Scholar] [CrossRef] [Green Version]
- Weindling, R. Trichoderma lignorum as a parasite of other soil fungi. Phytopath 1932, 22, 837–845. [Google Scholar]
- Anitha, R.; Murugesan, K. Production of gliotoxin on natural substrates by Trichoderma virens. J. Basic Microbiol. 2005, 45, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Bruce, W.; Dutcher, J.D.; Johnson, J.R.; Miller, L.L. Gliotoxin, the Antibiotic Principle of Gliocladium fimbriatum. II. General Chemical Behavior and Crystalline Derivatives1. J. Am. Chem. Soc. 2002, 66, 614–616. [Google Scholar] [CrossRef]
- Shah, D.T.; Larsen, B. Clinical isolates of yeast produce a gliotoxin-like substance. Mycopathologia 1991, 116, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Glister, G.A.; Williams, T.I. Production of Gliotoxin by Aspergillus fumigatus mut. helvola Yuill. Nature 1944, 153, 651. [Google Scholar] [CrossRef]
- Kupfahl, C.; Michalka, A.; Lass-Flörl, C.; Fischer, G.; Haase, G.; Ruppert, T.; Geginat, G.; Hof, H. Gliotoxin production by clinical and environmental Aspergillus fumigatus strains. Int. J. Med. Microbiol. 2008, 298, 319–327. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Larsen, T.O. Extrolites of Aspergillus fumigatus and Other Pathogenic Species in Aspergillus Section Fumigati. Front. Microbiol. 2015, 6, 1485. [Google Scholar] [CrossRef] [PubMed]
- Larsen, T.O.; Smedsgaard, J.; Nielsen, K.F.; Hansen, M.A.; Samson, R.A.; Frisvad, J.C. Production of mycotoxins by Aspergillus lentulus and other medically important and closely related species in section Fumigati. Med. Mycol. 2007, 45, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Vidal-Garcia, M.; Redrado, S.; Domingo, M.P.; Marquina, P.; Colmenarejo, C.; Meis, J.F.; Rezusta, A.; Pardo, J.; Galvez, E.M. Production of the Invasive Aspergillosis Biomarker Bis(methylthio)gliotoxin Within the Genus Aspergillus: In Vitro and in Vivo Metabolite Quantification and Genomic Analysis. Front. Microbiol. 2018, 9, 1246. [Google Scholar] [CrossRef] [PubMed]
- Sayers, E.W.; Beck, J.; Bolton, E.E.; Bourexis, D.; Brister, J.R.; Canese, K.; Comeau, D.C.; Funk, K.; Kim, S.; Klimke, W.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2021, 8, D10–D17. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Higgins, D.G. Clustal Omega, accurate alignment of very large numbers of sequences. Methods Mol. Biol. 2014, 1079, 105–116. [Google Scholar] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2--a multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, G. Estimating the Dimension of a Model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Hurvich, C.M. Regression and time series model selection in small samples. Biometrika 1989, 76, 297–307. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783. [Google Scholar] [CrossRef] [PubMed]
- Efron, B. The Jackknife, the Bootstrap and Other Resampling Plans. In Proceedings of the CBMS-NSF Regional Conference Series in Applied Mathematics, Monograph 38, SIAM, Philadelphia, PA, USA; 1982. [Google Scholar]
- Thomas, R.H. Molecular Evolution and Phylogenetics. Heredity 2001, 86, 385. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Stein, N. Synteny (Syntenic Genes). In Brenner’s Encyclopedia of Genetics, 2nd ed.; Maloy, S., Hughes, K., Eds.; Academic Press: San Diego, CA, USA, 2013; pp. 623–626. [Google Scholar]
- Harrison, K.J.; Crécy-Lagard, V.; Zallot, R. Gene Graphics: A genomic neighborhood data visualization web application. Bioinformatics 2018, 34, 1406–1408. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.E.; Wiederhold, N.P.; Chi, J.; Han, X.Y.; Komanduri, K.V.; Kontoyiannis, D.P.; Prince, R.A. Detection of gliotoxin in experimental and human aspergillosis. Infect. Immun. 2005, 73, 635–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puri, A.; Ahmad, A.; Panda, B.P. Development of an HPTLC-based diagnostic method for invasive aspergillosis. Biomed. Chromatogr. 2010, 24, 887–892. [Google Scholar] [CrossRef]
- Lewis, R.E.; Wiederhold, N.P.; Lionakis, M.S.; Prince, R.A.; Kontoyiannis, D.P. Frequency and species distribution of gliotoxin-producing Aspergillus isolates recovered from patients at a tertiary-care cancer center. J. Clin. Microbiol. 2005, 43, 6120–6122. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.-J.; Yeh, H.H.; Chiang, Y.M.; Sanchez, J.F.; Chang, S.L.; Bruno, K.S.; Wang, C.C. Biosynthetic pathway for the epipolythiodioxopiperazine acetylaranotin in Aspergillus terreus revealed by genome-based deletion analysis. J. Am. Chem. Soc. 2013, 135, 7205–7213. [Google Scholar] [CrossRef] [Green Version]
- Chankhamjon, P.; Boettger-Schmidt, D.; Scherlach, K.; Urbansky, B.; Lackner, G.; Kalb, D.; Dahse, H.M.; Hoffmeister, D.; Hertweck, C. Biosynthesis of the Halogenated Mycotoxin Aspirochlorine in Koji Mold Involves a Cryptic Amino Acid Conversion. Angew. Chem. Int. Ed. 2014, 53, 13409–13413. [Google Scholar] [CrossRef]
- Vargas, W.A.; Mukherjee, P.K.; Laughlin, D.; Wiest, A.; Moran-Diez, M.E.; Kenerley, C.M. Role of gliotoxin in the symbiotic and pathogenic interactions of Trichoderma virens. Microbiology 2014, 160 Pt 10, 2319–2330. [Google Scholar] [CrossRef] [PubMed]
- Baccile, J.A.; Le, H.H.; Pfannenstiel, B.T.; Bok, J.W.; Gomez, C.; Brandenburger, E.; Hoffmeister, D.; Keller, N.P.; Schroeder, F. Diketopiperazine Formation in Fungi Requires Dedicated Cyclization and Thiolation Domains. Angew. Chem. Int. Ed. 2019, 58, 14589–14593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, P.K.; Horwitz, B.A.; Kenerley, C.M. Secondary metabolism in Trichoderma—A genomic perspective. Microbiology 2012, 158 Pt 1, 35–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rokas, A. Phylogenetic analysis of protein sequence data using the Randomized Axelerated Maximum Likelihood (RAXML) Program. Curr. Protoc. Mol. Biol. 2011, 19, 11. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.H.; Yu, Z.G.; Han, G.S.; Yang, W.F.; Anh, V. Whole-proteome based phylogenetic tree construction with inter-amino-acid distances and the conditional geometric distribution profiles. Mol. Phylogenetics Evol. 2015, 89, 37–45. [Google Scholar] [CrossRef]
- He, L.; Li, Y.; He, R.L.; Yau, S.S. A novel alignment-free vector method to cluster protein sequences. J. Theor. Biol. 2017, 427, 41–52. [Google Scholar] [CrossRef]
- Li, Y.; Tian, K.; Yin, C.; He, R.L.; Yau, S.S. Virus classification in 60-dimensional protein space. Mol. Phylogenetics Evol. 2016, 99, 53–62. [Google Scholar] [CrossRef]
- Dey, G.; Meyer, T. Phylogenetic Profiling for Probing the Modular Architecture of the Human Genome. Cell Syst. 2015, 1, 106–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, H.; Kang, S.; Park, Y.S.; Yun, C.W. The Role of Zinc in Gliotoxin Biosynthesis of Aspergillus fumigatus. Int. J. Mol. Sci. 2019, 20, 6192. [Google Scholar] [CrossRef] [Green Version]
- Tamiya, H.; Ochiai, E.; Kikuchi, K.; Yahiro, M.; Toyotome, T.; Watanabe, A.; Yaguchi, T.; Kamei, K. Secondary metabolite profiles and antifungal drug susceptibility of Aspergillus fumigatus and closely related species, Aspergillus lentulus, Aspergillus udagawae, and Aspergillus viridinutans. J. Infect. Chemother. 2015, 21, 385–391. [Google Scholar] [CrossRef]
- Mead, M.E.; Knowles, S.L.; Raja, H.A.; Beattie, S.R.; Kowalski, C.H.; Steenwyk, J.L.; Silva, L.P.; Chiaratto, J.; Ries, L.N.A.; Goldman, G.H.; et al. Characterizing the Pathogenic, Genomic, and Chemical Traits of Aspergillus fischeri, Close Relative of the Major Human Fungal Pathogen Aspergillus fumigatus. mSphere 2019, 4, e00018-19. [Google Scholar] [CrossRef] [Green Version]
- Knowles, S.L.; Mead, M.E.; Silva, L.P.; Raja, H.A.; Steenwyk, J.L.; Goldman, G.H.; Oberlies, N.H.; Rokas, A. Gliotoxin, a Known Virulence Factor in the Major Human Pathogen Aspergillus fumigatus, Is Also Biosynthesized by Its Nonpathogenic Relative Aspergillus fischeri. MBio 2020, 11, e03361-19. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Takada, K.; Takemoto, Y.; Yoshida, M.; Nogi, Y.; Okada, S.; Matsunaga, S. Gliotoxin analogues from a marine-derived fungus, Penicillium sp., and their cytotoxic and histone methyltransferase inhibitory activities. J. Nat. Prod. 2012, 75, 111–114. [Google Scholar]
- Johnson, J.; Gliotoxin, X.I. A Related Antibiotic from Penicillium terlikowski: Gliotoxin Monoacetate. J. Am. Chem. Soc. 1953, 75, 2. [Google Scholar] [CrossRef]
- Waring, P. Gliotoxin-E: A New Biologically Active Epipolythiodioxopiperazine Isolated From Penicillium terlikowskii. Aust. J. Chem. 1987, 40, 7. [Google Scholar] [CrossRef]
- Zhang, H.A.; An, Z. Gliotoxin Analogues from Endophytic Penicillium sp. of Eucommia ulmoides and Their Antimicrobial Activity. Chem. Nat. Compd. 2019, 55, 3. [Google Scholar] [CrossRef]
- Intaraudom, C.; Boonyuen, N.; Suvannakad, R.; Rachtawee, P.; Pittayakhajonwut, P. Penicolinates A–E from endophytic Penicillium sp. BCC16054. Tetrahedron Lett. 2013, 54, 6. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Strain | Solid Culturing Conditions | Liquid Culturing Conditions |
|---|---|---|
| A. fumigatus B5233 | 37 °C | 37 °C |
| A. fumigatus 1631562 | 37 °C | 37 °C |
| A. fischeri CBS 420.96 | 37 °C | 37 °C |
| A. lentulus 353 | 37 °C | 37 °C |
| A. turcosus CBS 140371 | 37 °C | 37 °C |
| A. pseudofischeri CBS 404.67 | 37 °C | 37 °C |
| P. flavigenum CBS 110407 | 30 °C | 30 °C |
| P. expansum MD-8 | 30 °C | 30 °C |
| B. victoriae CBS 174.57 | 30 °C | 30 °C |
| E. ampelina CBS 208.25 | 30 °C | 30 °C |
| C. cylindrospora CBS 449.70 | 30 °C | 30 °C |
| C. fructicola CBS 120005 | 30 °C | 30 °C |
| T. virens CBS 249.59 | RT, daylight | RT, daylight |
| P. zonata CBS 506.65 | RT, daylight | RT, daylight |
| T. reesei CBS 383.78 | RT, daylight | RT, daylight |
| R. ligniota CBS 133067 | 30 °C | 30 °C |
| T. harzianum CBS 226.95 | RT, daylight | RT, daylight |
| T. parareesei CBS 125925 | RT, daylight | RT, daylight |
| Fungal Strain | Locus Tag | Accession | Score | Query Coverage | E Value | Identity |
|---|---|---|---|---|---|---|
| Aspergillus fumigatus Af293 | AFUA_6G09660 | EAL88817 | 4396 | 100% | 0.0 | 100.00% |
| Aspergillus fumigatus A1163 | AFUB_075710 | EDP49541 | 4386 | 100% | 0.0 | 99.77% |
| Aspergillus fischerii NRRL 181 | NFIA_055350 | XP_001258083 | 4152 | 100% | 0.0 | 94.75% |
| Aspergillus lentulus IFM 54703 | ALT_0509 | GAQ03188 | 4101 | 100% | 0.0 | 93.13% |
| Aspergillus novofumigatus IBT 16806 | P174DRAFT_511160 | XP_024683983 | 4012 | 100% | 0.0 | 92.06% |
| Aspergillus udagawae IFM 46973 | AUD_6972 | GAO88012 | 3920 | 100% | 0.0 | 89.30% |
| Aspergillus turcosus HMR AF 1038 | CFD26_102651 | RLL93873 | 3896 | 100% | 0.0 | 88.42% |
| Aspergillus thermomutatus HMR Af 39 | CDV56_101444 | XP_026610442 | 3779 | 100% | 0.0 | 87.07% |
| Penicillium flavigenum IBT 14082 | PENFLA_c013G03821 | OQE22222 | 2652 | 99% | 0.0 | 60.95% |
| Penicillium expansum MD-8 | PEX2_011780 | XP_016603809 | 2644 | 99% | 0.0 | 61.24% |
| Bipolaris vicotriae FI3 | COCVIDRAFT_32734 | EUN32662 | 1969 | 95% | 0.0 | 47.29% |
| Elsinoe ampelina CECT 20119 | BDZ85DRAFT_305103 | KAF2219165 | 1712 | 99% | 0.0 | 44.93% |
| Coleophoma cylindrospora BP6252 | BP6252_07505 | RDW70942 | 1642 | 95% | 0.0 | 44.21% |
| Colletotrichum fructicola CGMCC3.17371 | CGMCC3_g11493 | KAE9572546.1 | 1640 | 94% | 0.0 | 44.00% |
| Colletotrichum asianum ICMP 18580 | GQ607_003908 | KAF0328883 | 1627 | 94% | 0.0 | 44.09% |
| Trichoderma virens Gv29-8 | TRIVIDRAFT_78708 | EHK22005 | 1621 | 94% | 0.0 | 43.99% |
| Torrubiella hemipterigena BCC 1449 | VHEMI02393 | CEJ82322 | 1718 | 94% | 0.0 | 41.43% |
| Penicilliopsis zonata CBS 506.65 | ASPZODRAFT_160119 | OJJ45943.1 | 1481 | 96% | 0.0 | 42.54% |
| Trichoderma reesei QM6a | TRIREDRAFT_24586 | EGR52474.1 | 1461 | 98% | 0.0 | 39.94% |
| Rhizodiscina lignyota CBS 133067 | NA57DRAFT_81988 | KAF2092841 | 1461 | 98% | 0.0 | 39.34% |
| Trichoderma harzianum TR274 | CI102_1861 | PKK53446 | 1453 | 99% | 0.0 | 38.55% |
| Trichoderma parareesei CBS 125925 | A9Z42_0047170 | OTA04143 | 1433 | 99% | 0.0 | 39.13% |
| Fungal Strain | Czapek-Dox | RPMI 1640 | ||
|---|---|---|---|---|
| GT (µg/mL) | bmGT (µg/mL) | GT (µg/mL) | bmGT (µg/mL) | |
| A. fumigatus B5233 | 11.067 ± 14.286 | 2.595 ± 3.225 | 8.373 ± 2.663 | 11.131 ± 15.538 |
| A. fumigatus 1631562 | 3.913 ± 4.278 | 0.845 ± 0.805 | 4.154 ± 7.442 | 0.519 ± 0.052 |
| A. fischeri CBS 420.96 | 0.135 ± 0.077 | 0.222 ± 0.206 | 0.062 | 0.311 ± 0.108 |
| A. lentulus 353 | ND | ND | ND | ND |
| A. turcosus CBS 140371 | 0.085 ± 0.0539 | 1.215 ± 0.884 | 0.491 ± 0.585 | 1.795 ± 1.732 |
| A. pseudofischeri CBS 404.67 | 0.840 ± 0.028 | 0.518 ± 0.297 | 0.295 ± 0.092 | 0.253 ± 0.226 |
| P. flavigenum CBS 110407 | ND | ND | ND | ND |
| P. expansum MD-8 | ND | ND | ND | ND |
| E. ampelina CBS 208.25 | ND | ND | ND | ND |
| C. cylindrospora CBS 449.70 | ND | ND | ND | ND |
| C. fructicola CBS 120005 | ND | ND | ND | ND |
| T. virens CBS 249.59 | 0.840 ± 0.824 | 0.518 ± 1.762 | 0.330 ± 0.178 | 2.417 ± 1.697 |
| P. zonata CBS 506.65 | ND | ND | ND | ND |
| T. reesei CBS 383.78 | ND | ND | ND | ND |
| R. ligniota CBS 133067 | ND | ND | ND | ND |
| T. harzianum CBS 226.95 | ND | ND | ND | ND |
| T. parareesei CBS 125925 | ND | ND | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Redrado, S.; Esteban, P.; Domingo, M.P.; Lopez, C.; Rezusta, A.; Ramirez-Labrada, A.; Arias, M.; Pardo, J.; Galvez, E.M. Integration of In Silico and In Vitro Analysis of Gliotoxin Production Reveals a Narrow Range of Producing Fungal Species. J. Fungi 2022, 8, 361. https://doi.org/10.3390/jof8040361
Redrado S, Esteban P, Domingo MP, Lopez C, Rezusta A, Ramirez-Labrada A, Arias M, Pardo J, Galvez EM. Integration of In Silico and In Vitro Analysis of Gliotoxin Production Reveals a Narrow Range of Producing Fungal Species. Journal of Fungi. 2022; 8(4):361. https://doi.org/10.3390/jof8040361
Chicago/Turabian StyleRedrado, Sergio, Patricia Esteban, María Pilar Domingo, Concepción Lopez, Antonio Rezusta, Ariel Ramirez-Labrada, Maykel Arias, Julián Pardo, and Eva M. Galvez. 2022. "Integration of In Silico and In Vitro Analysis of Gliotoxin Production Reveals a Narrow Range of Producing Fungal Species" Journal of Fungi 8, no. 4: 361. https://doi.org/10.3390/jof8040361
APA StyleRedrado, S., Esteban, P., Domingo, M. P., Lopez, C., Rezusta, A., Ramirez-Labrada, A., Arias, M., Pardo, J., & Galvez, E. M. (2022). Integration of In Silico and In Vitro Analysis of Gliotoxin Production Reveals a Narrow Range of Producing Fungal Species. Journal of Fungi, 8(4), 361. https://doi.org/10.3390/jof8040361