
A New Lichenized Fungus, Psoroglaena humidosilvae, from a Forested Wetland of Korea, with a Taxonomic Key to the Species of Psoroglaena


Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological and Chemical Analyses
2.2. Isolation, DNA Extraction, Amplification, and Sequencing
2.3. Phylogenetic Analyses
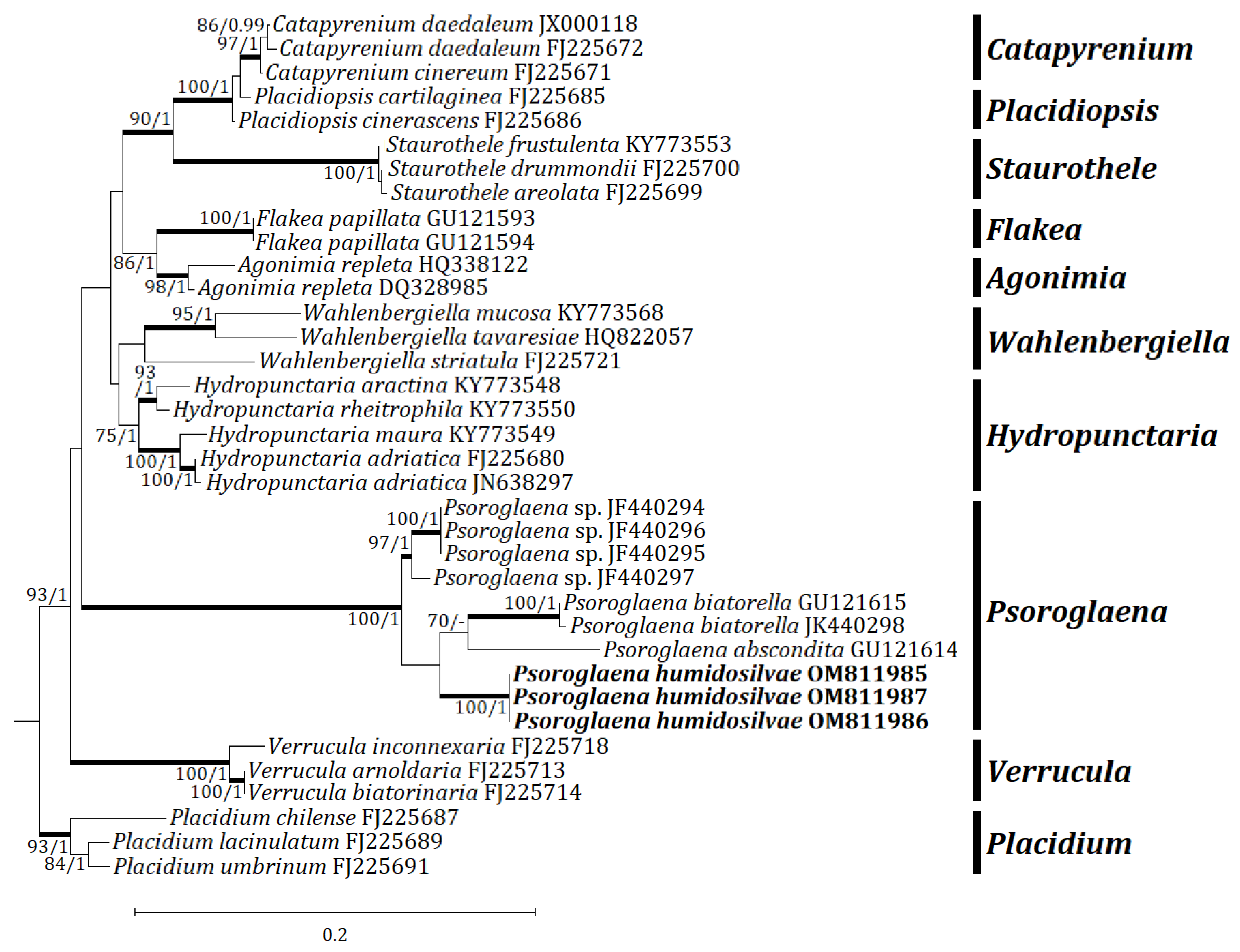
3. Results
3.1. Phylogenetic Analyses
3.2. Taxonomy
3.2.1. Psoroglaena humidosilvae B.G. Lee sp. nov.
3.2.2. Key to the Species of Psoroglaena
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Institutional Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Müller, J. Argoviensis. Lichenologische Beiträge. XXXV. Flora 1891, 74, 371–382. [Google Scholar]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L.; Haelewaters, D.; Rajeshkumar, K.C.; Zhao, R.L.; Aptroot, A.; Leontyev, D.V.; Saxena, R.K.; et al. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]
- Harada, H. Psoroglaena japonica (lichenized Ascomycota, Verrucariaceae), a new species from Chiba-ken, central Japan, with notes on Psoroglaena. Lichenology 2003, 2, 5–10. [Google Scholar]
- Lücking, R. Foliicolous Lichenized Fungi; New York Botanical Garden Press: New York, NY, USA, 2008; pp. 1–866. [Google Scholar]
- McCarthy, P.M.; Kantvilas, G. Psoroglaena halmaturina sp. nov. (lichenised Ascomycota, Verrucariaceae) from Kangaroo Island, South Australia. J. Adel. Bot. Gard. 2013, 26, 1–4. [Google Scholar]
- Orange, A. British and Other Pyrenocarpous Lichens. Available online: https://museum.wales/media/13849/Orange-A-2013-British-and-other-pyrenocarpous-lichens.pdf (accessed on 28 February 2022).
- Smith, C.W.; Aptroot, A.; Coppins, B.J.; Fletcher, A.; Gilbert, O.L.; James, P.W.; Wolseley, P.A. The Lichens of Great Britain and Ireland; The British Lichen Society: London, UK, 2009; pp. 1–1046. [Google Scholar]
- Halda, J.P.; Oh, S.O.; Liu, D.; Lee, B.G.; Kondratyuk, S.Y.; Lőkös, L.; Park, J.S.; Woo, J.J.; Hur, J.S. Two new lichen species, Thelopsis ullungdoensis and Phylloblastia gyeongsangbukensis from Korea. Mycobiology 2020, 48, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Muggia, L.; Gueidan, C.; Grube, M. Phylogenetic placement of some morphologically unusual members of Verrucariales. Mycologia 2010, 102, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Guzow-Krzemińska, B.; Halda, J.P.; Czarnota, P. A new Agonimia from Europe with a flabelliform thallus. Lichenologist 2012, 44, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Gueidan, C.; Roux, C.; Lutzoni, F. Using a multigene phylogenetic analysis to assess generic delineation and character evolution in Verrucariaceae (Verrucariales, Ascomycota). Mycol. Res. 2007, 111, 1145–1168. [Google Scholar] [CrossRef]
- Gueidan, C.; Savić, S.; Thüs, H.; Roux, C.; Keller, C.; Tibell, L.; Prieto, M.; Heiðmarsson, S.; Breuss, O.; Orange, A.; et al. Generic classification of the Verrucariaceae (Ascomycota) based on molecular and morphological evidence: Recent progress and remaining challenges. Taxon 2009, 58, 184–208. [Google Scholar] [CrossRef]
- Gueidan, C.; Thüs, H.; Pérez-Ortega, S. Phylogenetic position of the brown algae-associated lichenized fungus Verrucaria tavaresiae (Verrucariaceae). Bryologist 2011, 114, 563–569. [Google Scholar] [CrossRef]
- Prieto, M.; Martínez, I.; Aragón, G.; Gueidan, C.; Lutzoni, F. Molecular phylogeny of Heteroplacidium, Placidium, and related catapyrenioid genera (Verrucariaceae, lichen-forming Ascomycota). Am. J. Bot. 2012, 99, 23–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pykälä, J.; Myllys, L. Three new species of Atla from calcareous rocks (Verrucariaceae, lichenized Ascomycota). Lichenologist 2016, 48, 111–120. [Google Scholar] [CrossRef]
- Nyati, S.; Beck, A.; Honegger, R. Fine structure and phylogeny of green algal photobionts in the microfilamentous genus Psoroglaena (Verrucariaceae, lichen-forming ascomycetes). Plant Biol. 2007, 9, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Thüs, H.; Muggia, L.; Pérez-Ortega, S.; Favero-Longo, S.E.; Joneson, S.; O’Brien, H.; Nelsen, M.P.; Duque-Thüs, R.; Grube, M.; Friedl, T.; et al. Revisiting photobiont diversity in the lichen family Verrucariaceae (Ascomycota). Eur. J. Phycol. 2011, 46, 399–415. [Google Scholar] [CrossRef] [Green Version]
- Kondratyuk, S.Y.; Lőkös, L.; Halda, J.P.; Haji Moniri, M.; Farkas, E.; Park, J.S.; Lee, B.G.; Oh, S.O.; Hur, J.S. New and noteworthy lichen-forming and lichenicolous fungi 4. Acta Bot. Hung. 2016, 58, 75–136. [Google Scholar] [CrossRef]
- Kondratyuk, S.Y.; Lőkös, L.; Halda, J.P.; Upreti, D.K.; Mishra, G.K.; Haji Moniri, M.; Farkas, E.; Park, J.S.; Lee, B.G.; Liu, D.; et al. New and noteworthy lichen-forming and lichenicolous fungi 5. Acta Bot. Hung. 2016, 58, 319–396. [Google Scholar] [CrossRef] [Green Version]
- Kondratyuk, S.Y.; Lőkös, L.; Halda, J.P.; Roux, C.; Upreti, D.K.; Schumm, F.; Mishra, G.K.; Nayaka, S.; Farkas, E.; Park, J.S.; et al. New and noteworthy lichen-forming and lichenicolous fungi 6. Acta Bot. Hung. 2017, 59, 137–260. [Google Scholar] [CrossRef]
- Kondratyuk, S.Y.; Lőkös, L.; Halda, J.P.; Farkas, E.; Upreti, D.K.; Thell, A.; Woo, J.J.; Oh, S.O.; Hur, J.S. New and noteworthy lichen-forming and lichenicolous fungi 7. Acta Bot. Hung. 2018, 60, 115–184. [Google Scholar] [CrossRef]
- Liu, D.; Halda, J.P.; Oh, S.O.; Park, C.H.; Hur, J.S. New records of pyrenocarpous lichens from Jeju Island, South Korea. Mycotaxon 2018, 133, 127–139. [Google Scholar] [CrossRef]
- Orange, A.; James, P.W.; White, F.J. Microchemical Methods for the Identification of Lichens; The British Lichen Society: London, UK, 2001; pp. 1–101. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, D., Sninsky, J., White, T., Eds.; Academic Press: Cambridge, MA, USA, 1990; Volume 1, pp. 315–322. [Google Scholar]
- Zoller, S.; Scheidegger, C.; Sperisen, C. PCR primers for the amplification of mitochondrial small subunit ribosomal DNA of lichen-forming ascomycetes. Lichenologist 1999, 31, 511–516. [Google Scholar] [CrossRef]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Ekman, S. Molecular phylogeny of the Bacidiaceae (Lecanorales, lichenized Ascomycota). Mycol. Res. 2001, 105, 783–797. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. raxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Methods Ecol. Evol. 2021, 12, 373–377. [Google Scholar] [CrossRef]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouckaert, R.R.; Drummond, A.J. bModelTest: Bayesian phylogenetic site model averaging and model comparison. BMC Evol. Biol. 2017, 17, e42. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree v1.4.2. Edinburgh: University of Edinburgh. Available online: http://tree.bio.ed.ac.uk/software/figtree (accessed on 28 February 2022).
- Lee, B.G.; Hur, J.S. A new lichenized fungus, Lecanora baekdudaeganensis, from South Korea, with a taxonomic key for Korean Lecanora species. MycoKeys 2020, 70, 39–58. [Google Scholar] [CrossRef]
- Orange, A. Macentina stigonemoides (Verrucariaceae), a new lichenized species from Great Britain and Ireland. Lichenologist 1989, 21, 229–236. [Google Scholar] [CrossRef]
- Coppins, B.J.; Vězda, A. Macentina, a lichen genus new to Europe. Lichenologist 1977, 9, 47–49. [Google Scholar] [CrossRef]
- Lee, B.G.; Hur, J.S. Two new calcicolous caloplacoid lichens from South Korea, with a taxonomic key to the species of Huriella and Squamulea. MycoKeys 2021, 84, 35–55. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Species | ITS | mtSSU | LSU | Voucher |
|---|---|---|---|---|---|
| 1 | Agonimia repleta | - | DQ328985 | DQ329014 | F 446 |
| 2 | Agonimia repleta | JF509161 | - | - | GPN 5935 |
| 3 | Agonimia repleta | JF509160 | HQ338122 | - | Palice 12970 |
| 4 | Agonimia repleta | - | - | GU121554 | Palice 6464 |
| 5 | Catapyrenium cinereum | GQ344598 | - | GQ344587 | MA16301 |
| 6 | Catapyrenium cinereum | - | FJ225671 | EF643747 | S. Heidmarsson 2017 |
| 7 | Catapyrenium daedaleum | - | FJ225672 | EF643748 | C. Gueidan 115 |
| 8 | Catapyrenium daedaleum | JX000099 | JX000118 | - | Prieto 3051 |
| 9 | Flakea papillata | - | GU121593 | - | Perlmutter 1267 |
| 10 | Flakea papillata | KT232210 | - | KT232216 | T. Tonsberg 34186 |
| 11 | Flakea papillata | LC574000 | - | - | TNS:YO12329 |
| 12 | Flakea papillata | - | GU121594 | FJ455775 | Perlmutter 1263 |
| 13 | Hydropunctaria adriatica | - | FJ225680 | EF643783 | C. Gueidan 669 |
| 14 | Hydropunctaria adriatica | JN638251 | JN638297 | - | NMW < GBR_:C.2011.014.61 |
| 15 | Hydropunctaria aractina | KY697128 | KY773548 | KY773248 | DUKE:LA31123 |
| 16 | Hydropunctaria maura | KY697129 | KY773549 | KY773249 | AMNH:LA31903 |
| 17 | Hydropunctaria rheitrophila | KY697130 | KY773550 | KY773250 | AMNH:LA30737 |
| 18 | Placidiopsis cartilaginea | - | FJ225685 | EF643758 | C. Gueidan 458 |
| 19 | Placidiopsis cinerascens | - | FJ225686 | EF643759 | C. Gueidan 585 |
| 20 | Placidiopsis cinerascens | GQ344609 | - | - | MA161305 |
| 21 | Placidiopsis tenella | GQ344616 | - | GQ344575 | Ll 281352 |
| 22 | Placidium chilense | - | FJ225687 | - | O. Breuss (DUKE, s.n.) |
| 23 | Placidium fingens | GU228989 | - | GU228935 | LI 271006 |
| 24 | Placidium lachneum | GU228982 | - | GU228926 | M. Prieto 322 |
| 25 | Placidium lacinulatum | - | FJ225689 | EF643762 | K. Knudsen 733 |
| 26 | Placidium pilosellum | GU228968 | - | GU228907 | M. Prieto 439 |
| 27 | Placidium tenellum | GU228990 | - | GQ344562 | MA16300 |
| 28 | Placidium umbrinum | - | FJ225691 | - | O. Breuss (LI, 14.461) |
| 29 | Psoroglaena abscondita | - | GU121614 | GU121572 | JN1742 |
| 30 | Psoroglaena biatorella | - | GU121615 | GU121573 | Hafellner 64389 |
| 31 | Psoroglaena biatorella | - | JF440298 | - | KRAM-51901 |
| 32 | Psoroglaena humidosilvae | OM811988 | OM811985 | OM811982 | KBA-L-00002005 |
| 33 | Psoroglaena humidosilvae | OM811989 | OM811986 | OM811983 | KBA-L-00002007 |
| 34 | Psoroglaena humidosilvae | OM811990 | OM811987 | OM811984 | KBA-L-00002008 |
| 35 | Psoroglaena sp. | JF939053 | - | - | KRAM:L-64117 |
| 36 | Psoroglaena sp. | - | JF440296 | - | KRAM:L-64117 (KRAP 13/4) |
| 37 | Psoroglaena sp. | - | JF440295 | - | KRAM:L-64117 (KRAP 13/9) |
| 38 | Psoroglaena sp. | - | JF440294 | - | KRAM:L-64117 (KRAP 13/5) |
| 39 | Psoroglaena sp. | - | JF440297 | - | KRAM:L-64117 (KRAP 18/8) |
| 40 | Psoroglaena stigonemoides | - | - | GU121574 | - |
| 41 | Psoroglaena stigonemoides | MZ159334 | - | - | K(M):98005 |
| 42 | Staurothele areolata | EU006543 | FJ225699 | EF643772 | C. Gueidan 378 |
| 43 | Staurothele drummondii | KY769539 | FJ225700 | EF643774 | C. Gueidan 831 |
| 44 | Staurothele frustulenta | KY697133 | KY773553 | KY773278 | DUKE:Heidmarsson 2066 |
| 45 | Verrucula arnoldaria | EU006534 | FJ225713 | EF643816 | C. Gueidan 594 |
| 46 | Verrucula biatorinaria | EU006533 | FJ225714 | EF643815 | C. Roux 24443 |
| 47 | Verrucula inconnexaria | EU006538 | FJ225718 | EF643821 | C. Roux 22721 |
| 48 | Wahlenbergiella mucosa | KY697148 | KY773568 | KY773263 | AMNH:LA31918 |
| 49 | Wahlenbergiella striatula | - | FJ225721 | EF643810 | C. Gueidan 688 |
| 50 | Wahlenbergiella striatula | MT809484 | - | - | FH:BHI-F1163 |
| 51 | Wahlenbergiella tavaresiae | HQ822056 | HQ822057 | HQ822059 | C. Gueidan 1101 |
| Overall | 30 | 36 | 34 |
| Species | Psoroglaena humidosilvae | Psoroglaena abscondita | Psoroglaena chirisanensis | Psoroglaena stigonemoides |
|---|---|---|---|---|
| Projections on thallus | locally a few projections | not observed | not observed | minute projections developed, which form minutely fruticose thallus |
| Perithecia color | pale yellow | pale brown | pale yellow to pale yellow-brown | pale brown |
| Perithecia (mm in diam.) | 0.1–0.2 | c. 0.1 | c. 0.2 | 0.2–0.4 |
| Perithecia shape in section | globose | globose | – | ovoid-obpyriform |
| Exciple thickness (μm) | c. 20 | c. 10 | 25–30 | 25–30 |
| Asci (μm) | 30–65 × 10–20 | 45–50 × 10–15 | c. 40 × 12 | 80–100 × 10–12 * |
| Ascospore septation | (1-)3-septate | 1–3-septate | (1-)3-septate | 3–4-(5-)septate |
| Ascospores (μm) | 12–18 × 4.5–6.5 | 12–20 × 3.5–4.5 | 18–20 × 5.5–6.5 | 16–21 × 5–6 13–23 × 4–6 * |
| Ascospore constriction at septa | generally not constricted, or slightly constricted for 1-septate spores | not constricted | distinctly constricted | slightly constricted |
| Substrate | bark (Salix koreensis) | bark (Sambucus, Juniperus) | bark (Robinia pseudoacasia) | bark (Sambucus, Acer, Quercus, Salix, Ulmus); moss; leaf |
| Reference | KBA-L-0002000 (paratype), KBA-L-0002005 (paratype), KBA-L-0002006 (paratype), KBA-L-0002007 (holotype) | [34,35] | [18] | [4,34] |
| 1. | On rock | 2 |
| – | On bark, moss or leaf | 4 |
| 2. | Perithecia colorless, 0.1–0.2 mm diam.; goniocyst absent; ascospores 18–22 × 5.5–6.5 μm | P. coreana |
| – | Perithecia orangish to dark brown, 0.2–0.4 mm diam.; goniocyst present; ascospores 11–17 × 5–6.5 μm | 3 |
| 3. | Perithecia orange-pink; goniocyst 20–40 μm diam.; ascospores generally 1-septate (rarely 3-septate), 11.5–13.5 × 5.5–6.5 μm | P. infossa |
| – | Perithecia pale orange-brown to dark brown; goniocyst 10–20 μm diam.; ascospores 3-septate, 13–17 × 5–6 μm | P. japonica |
| 4. | On moss or leaf | 5 |
| – | On bark | 14 |
| 5. | On moss | 6 |
| – | On leaf | 8 |
| 6. | Thallus inconspicuous, not minutely fruticose; perithecia less than 0.2 mm diam.; particularly growing on Radula flaccida | P. hepaticicola |
| – | Thallus crustose to minutely fruticose; perithecia over 0.2 mm diam. | 7 |
| 7. | Thallus generally granular-verrucose, with soredia-like granules; perithecia 0.2–0.6 mm diam.; ascospores 3-septate (young) to submuriform, 28–38 × 10–14 μm; sometimes growing on humus-rich ground | P. biatorella |
| – | Thallus generally minutely fruticose, locally with soredia-like granules; perithecia 0.2–0.4 mm diam.; ascospores transversely 3–5-septate, 16–21 × 5–6 μm | P. stigonemoides |
| 8. | Thallus appressed filamentous to microsquamulose; photobiont cells in distinct, uni- or biseriate filaments | 9 |
| – | Thallus crustose, smooth or minutely arachnoid-tomentose by fungal hyphae (not algal filaments!); photobiont cells in short, irregular threads or irregular plates to almost solitary | 12 |
| 9. | Thallus microsquamulose with erect, irregularly branched, bi- or triseriate filaments; ascospores transversely 3–5-septate | P. stigonemoides |
| – | Thallus appressed filamentous with regularly branched to almost unbranched, uni-or rarely biseriate filaments; ascospores (as far known) submuriform to muriform | 10 |
| 10. | Thallus with punctiform soralia | P. sorediata |
| – | Thallus lacking soralia | 11 |
| 11. | Thallus rather large (up to 100 mm across), composed of mostly parallel, very long and rarely branched, uniseriate filaments, pale olive-green | P. epiphylla |
| – | Thallus smaller (up to 30 mm across), composed of richly branched, uni- or biseriate filaments forming rounded rosettes, vivid green | P. ornata |
| 12. | Thallus minutely arachnoid-tomentose; perithecia with algal layer between excipulum and (secondary) involucrellum | P. arachnoidea |
| – | Thallus smooth, perithecia lacking distinct algal layer | 13 |
| 13. | Thallus continuous, without prothallus, lacking isidia, perithecia pale yellow; ascospores 3-septate, 16–24 × 3–4 μm | P. perminuta |
| – | Thallus dispersed into irregular patches on dark prothallus, with scattered, disc-shaped isidia; perithecia dark brownish gray to black; ascospores 3–5-septate, 22–27 × 5–7 μm | P. laevigata |
| 14. | Thallus filamentous to microsquamulose | 15 |
| – | Thallus crustose, leprose without elongated or branched projections | 20 |
| 15. | Perithecia not seen; thallus minutely fruticose, with dense cortical papillae (c. 2 × 1 μm) | P. spinosa |
| – | Perithecia present; thallus generally minutely fruticose or granular with locally soredia-like granules | 16 |
| 16. | Ascospores submuriform to muriform | 17 |
| – | Ascospores transversely septate only | 18 |
| 17. | Ascospores 60–70 × 15–20 μm | P. costaricensis |
| – | Ascospores 14–18 × 8–9 μm | P. sunchonensis |
| 18. | Ascospores 7-septate, 18–24 μm long | P. cubensis |
| – | Ascospores 3–5-septate, 12–21 μm long | 19 |
| 19. | Thallus minutely fruticose with heavy projections; perithecia pale brown, ovoid-obpyriform, 0.2–0.4 mm diam.; asci 80–100 × 10–12 μm; ascospores 3–5-septate, 16–21 × 5–6 μm | P. stigonemoides |
| – | Thallus smooth to granular with locally minute projections; perithecia pale yellow, globose, 0.1–0.2 mm diam.; asci 32–65 × 10–20 μm; ascospores (1-)3-septate, 12–18 × 4.5–6.5 μm | P. humidosilvae |
| 20. | Ascospores muriform | 21 |
| – | Ascospores transversely septate only | 22 |
| 21. | Ascospores up to 18 × 6 μm | P. dictyospora |
| – | Ascospores 22–29 × 7–9 μm | P. gangwondoensis |
| 22. | Perithecia dark olive-brown to dull black; ascospores (5-)7-septate | P. halmaturina |
| – | Perithecia pale yellow to pale brown; ascospores (1-)3-septate | 23 |
| 23. | Thallus leprose; perithecia 80–120 μm wide; exciple 10–12 μm thick; ascospores 3.5–4.5 μm wide | P. abscondita |
| – | Thallus granular to verrucose; perithecia 170–180 μm wide; exciple 25–30 μm thick; ascospores 5.5–6.5 μm wide | P. chirisanensis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, B.G.; Hur, J.-S. A New Lichenized Fungus, Psoroglaena humidosilvae, from a Forested Wetland of Korea, with a Taxonomic Key to the Species of Psoroglaena. J. Fungi 2022, 8, 392. https://doi.org/10.3390/jof8040392
Lee BG, Hur J-S. A New Lichenized Fungus, Psoroglaena humidosilvae, from a Forested Wetland of Korea, with a Taxonomic Key to the Species of Psoroglaena. Journal of Fungi. 2022; 8(4):392. https://doi.org/10.3390/jof8040392
Chicago/Turabian StyleLee, Beeyoung Gun, and Jae-Seoun Hur. 2022. "A New Lichenized Fungus, Psoroglaena humidosilvae, from a Forested Wetland of Korea, with a Taxonomic Key to the Species of Psoroglaena" Journal of Fungi 8, no. 4: 392. https://doi.org/10.3390/jof8040392
APA StyleLee, B. G., & Hur, J. -S. (2022). A New Lichenized Fungus, Psoroglaena humidosilvae, from a Forested Wetland of Korea, with a Taxonomic Key to the Species of Psoroglaena. Journal of Fungi, 8(4), 392. https://doi.org/10.3390/jof8040392