
Fungal Pathogens Associated with Crown and Root Rot of Wheat in Central, Eastern, and Southeastern Kazakhstan


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Isolation and Maintenance of Cultures
2.3. DNA Extraction
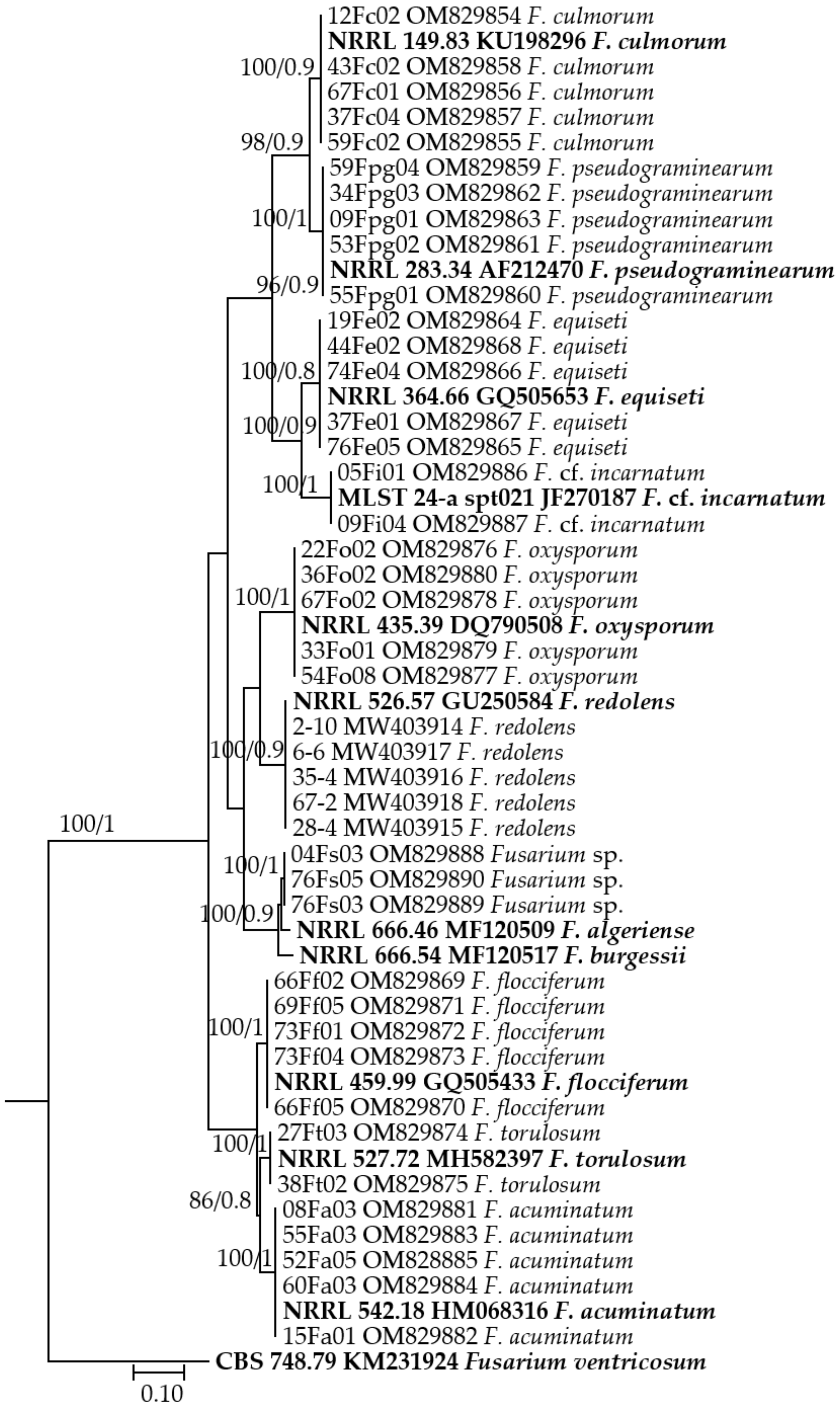
2.4. Species Identification and Phylogenetic Analysis
2.5. Isolation Frequency and Incidence of Fungal Species in the Fields
2.6. Pathogenicity Tests
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shiferaw, B.; Smale, M.; Braun, H.J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef] [Green Version]
- FAOSTAT. Food and Agriculture Organization Statistical Database. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 20 February 2022).
- Rsaliyev, A.S.; Rsaliyev, S.S. Principal approaches and achievements in studying race composition of wheat stem rust. Vavilov J. Genet. Breed. 2018, 22, 967–977. [Google Scholar] [CrossRef]
- Rsaliyev, A.; Yskakova, G.; Maulenbay, A.; Zakarya, K.; Rsaliyev, S. Virulence and race structure of Puccinia graminis f. sp. tritici in Kazakhstan. Plant Protect. Sci. 2020, 56, 275–284. [Google Scholar] [CrossRef]
- Koyshybaev, M. Wheat Diseases, 1st ed.; FAO: Ankara, Turkey, 2018; p. 365. [Google Scholar]
- Kuttibayevich, S.S.; Slyamkhanovna, T.Z.; Zharakovich, M.M.; Tontaevna, Y.A.; Mukashevna, T.O.; Amirzhanovna, I.G.; Klara, I.; Gulnara, N.; Dossym, B.; Faritovna, S.G. Classification of wheat yellow rust populations in the conditions of Kazakhstan (Puccinia striiformis West. f. sp. tritici Erikss. et Henn.). Bull. Natl. Acad. Sci. Repub. Kazakhstan 2019, 1, 263–268. [Google Scholar] [CrossRef]
- Kokhmetova, A.M.; Atishova, M.N.; Kumarbayeva, M.T.; Leonova, I.N. Phytopathological screening and molecular marker analysis of wheat germplasm from Kazakhstan and CIMMYT for resistance to tan spot. Вавилoвский Журнал Генетики И Селекции 2019, 23, 879–886. [Google Scholar] [CrossRef]
- Kokhmetova, A.; Kremneva, O.; Volkova, G.; Atishova, M.; Sapakhova, Z. Evaluation of wheat cultivars growing in Kazakhstan and Russia for resistance to tan spot. J. Plant Pathol. 2017, 99, 161–167. [Google Scholar] [CrossRef]
- Gultyaeva, E.I.; Kovalenko, N.M.; Shamanin, V.P.; Tyunin, V.A.; Shreyder, E.R.; Shaydayuk, E.L.; Morgunov, A.I. Population structure of leaf pathogens of common spring wheat in the West Asian regions of Russia and North Kazakhstan in 2017. Vavilov J. Genet. Breed. 2018, 22, 363–369. [Google Scholar] [CrossRef]
- Gultyaeva, E.; Yusov, V.; Rosova, M.; Mal’chikov, P.; Shaydayuk, E.; Kovalenko, N.; Rsaliyev, A. Evaluation of resistance of spring durum wheat germplasm from Russia and Kazakhstan to fungal foliar pathogens. Cereal Res. Commun. 2020, 48, 71–79. [Google Scholar] [CrossRef]
- Singh, R.P.; Singh, P.K.; Rutkoski, J.; Hodson, D.P.; He, X.; Jørgensen, L.N. Disease impact on wheat yield potential and prospects of genetic control. Annu. Rev. Phytopathol. 2016, 54, 303–322. [Google Scholar] [CrossRef] [Green Version]
- Kumar, J.; Schäfer, P.; Hückelhoven, R.; Langen, G.; Baltruschat, H.; Stein, E. Bipolaris sorokiniana, a cereal pathogen of global concern: Cytological and molecular approaches towards better control. Mol. Plant Pathol. 2002, 3, 185–195. [Google Scholar] [CrossRef]
- Kazan, K.; Gardiner, D.M. Fusarium crown rot caused by Fusarium pseudograminearum in cereal crops: Recent progress and future prospects. Mol. Plant Pathol. 2018, 19, 1547–1562. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Zhao, J.; Zhao, S.; Li, M.; Pang, S.; Kang, Z.; Zhen, W.; Chen, S.; Chen, F.; Wang, X. Genetics of resistance to common root rot (spot blotch), Fusarium crown rot, and sharp eyespot in wheat. Front. Genet. 2021, 12, 699342. [Google Scholar] [CrossRef]
- Bockus, W.W.; Bowden, R.L.; Hunger, R.M.; Morrill, W.L.; Murray, T.D.; Smiley, R.W. Compendium of Wheat Diseases and Pests, 3rd ed.; APS Press: St. Paul, MN, USA, 2010. [Google Scholar]
- Dyer, A.T.; Johnston, R.H.; Hogg, A.C.; Johnston, J.A. Comparison of pathogenicity of the Fusarium crown rot (FCR) complex (F. culmorum, F. pseudograminearum and F. graminearum) on hard red spring and durum wheat. Eur. J. Plant Pathol. 2009, 125, 387–395. [Google Scholar] [CrossRef]
- Bockus, W.W.; Bowden, R.L.; Hunger, R.M.; Morrill, W.L.; Murray, T.D.; Smiley, R.W.; Cook, R.J. Fusarium root, crown, and foot rots and associated seedling diseases. In Compendium of Wheat Diseases and Pests, 3rd ed.; APS Press: St. Paul, MN, USA, 2010; pp. 37–39. [Google Scholar]
- Fernandez, M.R.; Conner, R.L. Root and crown rot of wheat. Prairie Soils Crops J. 2011, 4, 151–157. [Google Scholar]
- Xu, F.; Yang, G.; Wang, J.; Song, Y.; Liu, L.; Zhao, K.; Li, Y.; Han, Z. Spatial distribution of root and crown rot fungi associated with winter wheat in the North China Plain and its relationship with climate variables. Front. Microbiol. 2018, 9, 1054. [Google Scholar] [CrossRef]
- Smiley, R.W.; Gourlie, J.A.; Easley, S.A.; Patterson, L.M. Pathogenicity of fungi associated with the wheat crown rot complex in Oregon and Washington. Plant Dis. 2005, 89, 949–957. [Google Scholar] [CrossRef] [Green Version]
- Özer, G.; İmren, M.; Bayraktar, H.; Paulitz, T.C.; Muminjanov, H.; Dababat, A.A. First report of Fusarium hostae causing crown rot on wheat in Azerbaijan. Plant Dis. 2019, 103, 3278. [Google Scholar] [CrossRef]
- White, D.J.; Chen, W.; Schroeder, K.L. Assessing the contribution of ethaboxam in seed treatment cocktails for the management of metalaxylresistant Pythium ultimum var. ultimum in Pacific Northwest spring wheat production. Crop Prot. 2019, 115, 7–12. [Google Scholar] [CrossRef]
- Özer, G.; Paulitz, T.C.; İmren, M.; Alkan, M.; Muminjanov, H.; Dababat, A.A. Identity and pathogenicity of fungi associated with crown and root rot of dryland winter wheat in Azerbaijan. Plant Dis. 2020, 104, 2149–2157. [Google Scholar] [CrossRef]
- Grudzinska-Sterno, M.; Yuen, J.; Stenlid, J.; Djurle, A. Fungal communities in organically grown winter wheat affected by plant organ and development stage. Eur. J. Plant Pathol. 2016, 146, 401–417. [Google Scholar] [CrossRef]
- Smiley, R.W.; Gourlie, J.A.; Easley, S.A.; Patterson, L.M.; Whittaker, R.G. Crop damage estimates for crown rot of wheat and barley in the Pacific northwest. Plant Dis. 2005, 89, 595–604. [Google Scholar] [CrossRef] [Green Version]
- Moya-Elizondo, E.A.; Rew, L.J.; Jacobsen, B.J.; Hogg, A.C.; Dyer, A.T. Distribution and prevalence of Fusarium crown rot and common root rot pathogens of wheat in Montana. Plant Dis. 2011, 95, 1099–1108. [Google Scholar] [CrossRef] [Green Version]
- Peralta, A.L.; Sun, Y.; Mcdaniel, M.D.; Lennon, J.T. Crop rotational diversity increases disease suppressive capacity of soil microbiomes. Ecosphere 2018, 9, e02235. [Google Scholar] [CrossRef]
- Pariyar, S.R.; Erginbas-Orakci, G.; Dadshani, S. Dissecting the genetic complexity of Fusarium crown rot resistance in wheat. Sci. Rep. 2020, 10, 3200. [Google Scholar] [CrossRef] [Green Version]
- Al-Sadi, A.M. Bipolaris sorokiniana-induced black point, common root rot, and spot blotch diseases of wheat: A review. Front. Cell. Infect. Microbiol. 2021, 11, 1–9. [Google Scholar] [CrossRef]
- Scherm, B.; Balmas, V.; Spanu, F.; Pani, G.; Delogu, G.; Pasquali, M.; Migheli, Q. Fusarium culmorum: Causal agent of foot and root rot and head blight on wheat. Mol. Plant Pathol. 2013, 14, 323–341. [Google Scholar] [CrossRef]
- Karlsson, I.; Persson, P.; Friberg, H. Fusarium head blight from a microbiome Perspective. Front Microbiol. 2021, 12, 628373. [Google Scholar] [CrossRef]
- Alkan, M.; Bayraktar, H.; İmren, M.; Özdemir, F.; Lahlali, R.; Mokrini, F.; Özer, G. Monitoring of Host Suitability and Defense-Related Genes in Wheat to Bipolaris sorokiniana. J. Fungi 2022, 8, 149. [Google Scholar] [CrossRef]
- Pérez-Méndez, N.; Miguel-Rojas, C.; Jimenez-Berni, J.A.; Gomez-Candon, D.; Pérez-de-Luque, A.; Fereres, E.; Catala-Forner, M.; Villegas, D.; Sillero, J.C. Plant breeding and management strategies to minimize the impact of water scarcity and biotic stress in cereal crops under Mediterranean conditions. Agronomy 2022, 12, 75. [Google Scholar] [CrossRef]
- Bozoğlu, T.; Özer, G.; İmren, M.; Paulitz, T.C.; Dababat, A.A. First report of crown rot caused by Fusarium redolens on wheat in Kazakhstan. Plant Dis. 2021, 105, 3302. [Google Scholar] [CrossRef]
- Özer, G.; İmren, M.; Bozoğlu, T.; Dababat, A.A. First report of Rhizoctonia solani AG2-1 on roots of wheat in Kazakhstan. Plant Dis. 2021, 105, 3744. [Google Scholar] [CrossRef]
- Eken, C.; Spanbayev, A.; Tulegenova, Z.; Yechshzhanov, T. First report of Nigrospora oryzae on wheat in Kazakhstan. Plant Dis. 2016, 100, 861. [Google Scholar] [CrossRef]
- The Fusarium Laboratory Manual; Leslie, J.F.; Summerell, B.A. (Eds.) Blackwell: Ames, IO, USA, 2006. [Google Scholar]
- Ellis, M.B. Dematiaceous Hyphomycetes; CAB: Surrey, UK, 1971. [Google Scholar]
- Sivanesan, A. Graminicolous species of Bipolaris, Curvularia, Drechslera, Exserohilum and their teleomorphs. Mycol. Papers 1987, 158, 1–126. [Google Scholar]
- Hudson, H.J. The perfect state of Nigrospora oryzae. Trans. Brit. Mycol. Soc. 1963, 46, 355–360. [Google Scholar] [CrossRef]
- Holliday, P.; Punithalingam, E. Macrophomina phaseolina. C.M.I. Descr. Pathog. Fungi Bact. 1970, 275, 1–2. [Google Scholar]
- O’Donnell, K.; Cigelnik, E.; Nirenberg, H.I. Molecular systematics and phylogeography of the Gibberella fujikuroi species complex. Mycologia 1998, 90, 465–493. [Google Scholar] [CrossRef] [Green Version]
- White, T.T.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Berbee, M.L.; Pirseyedi, M.; Hubbard, S. Cochliobolus phylogenetics and the origin of known, highly virulent pathogens, inferred from ITS and glyceraldehyde-3-phosphate dehydrogenase gene sequences. Mycologia 1999, 91, 964–977. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQTREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Duczek, L.J.; Verma, P.R.; Spurr, D.T. Effect of inoculum density of Cochliobolus sativus on common root rot of wheat and barley. Can. J. Plant Pathol. 1985, 7, 382–386. [Google Scholar] [CrossRef]
- Demirci, E. Rhizoctonia species and anastomosis groups isolated from barley and wheat in Erzurum, Turkey. Plant Pathol. 1998, 47, 10–15. [Google Scholar] [CrossRef]
- Nicol, J.M.; Rivoal, R.; Trethowan, R.M.; Van Ginkel, M.; Mergoum, M.; Singh, R.P. CIMMYT’s Approach to Identify And Use Resistance to Nematodes and Soilborne Fungi, in Developing Superior Wheat Germplasm. In Wheat in a Global Environment, Proceedings of the 6th International Wheat Conference, Budapest, Hungary, 5–9 June 2000; Bedo, Z., Lang, L., Eds.; Springer: Dordrecht, The Netherlands, 2001. [Google Scholar] [CrossRef]
- Wildermuth, G.B.; McNamara, R.B. Testing wheat seedlings for resistance to crown rot caused by Fusarium graminearum Group 1. Plant Dis. 1994, 78, 949–953. [Google Scholar] [CrossRef]
- Adhikari, S.; Kumari, J.; Jacob, S.R. Landraces-potential treasure for sustainable wheat improvement. Genet. Resour. Crop Evol. 2022, 69, 499–523. [Google Scholar] [CrossRef]
- Agrios, G.N. Plant Pathology, 4th ed.; Academic Press: London, UK, 1997; p. 635. [Google Scholar]
- Fernandez, M.R.; Basnyat, P.; Zentner, R.P. Response of common root rot in wheat to crop management in eastern Saskatchewan. Can. J. Plant Sci. 2007, 87, 953–963. [Google Scholar] [CrossRef] [Green Version]
- Moya-Elizondo, E.; Arismendi, N.; Castro, M.P.; Doussoulin, H. Distribution and prevalence of crown rot pathogens affecting wheat crops in southern Chile. Chil. J. Agric. Res. 2015, 75, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Alahmad, S.; Simpfendorfer, S.; Bentley, A.R. Crown rot of wheat in Australia: Fusarium pseudograminearum taxonomy, population biology and disease management. Australas. Plant Pathol. 2018, 47, 285–299. [Google Scholar] [CrossRef]
- Paulitz, T.C.; Smiley, R.W.; Cook, R.J. Insights into the prevalence and management of soilborne cereal pathogens under direct seeding in the Pacific Northwest, USA. Can. J. Plant Pathol. 2002, 24, 416–428. [Google Scholar] [CrossRef]
- Tunali, B.; Nicol, J.M.; Hodson, D.; Uçkun, Z.; Büyük, O.; Erdurmuş, D. Root and crown rot fungi associated with spring, facultative, and winter wheat in Turkey. Plant Dis. 2008, 92, 1299–1306. [Google Scholar] [CrossRef] [Green Version]
- Shikur Gebremariam, E.; Sharma-Poudyal, D.; Paulitz, T.C. Identity and pathogenicity of Fusarium species associated with crown rot on wheat (Triticum spp.) in Turkey. Eur. J. Plant Pathol. 2018, 150, 387–399. [Google Scholar] [CrossRef]
- Zhou, H.; He, X.; Wang, S.; Ma, Q.; Sun, B.; Ding, S.; Chen, L.; Zhang, M.; Li, H. Diversity of the Fusarium pathogens associated with crown rot in the Huanghuai wheat-growing region of China. Environ. Microbiol. 2019, 21, 2740–2754. [Google Scholar] [CrossRef]
- Mathieson, J.T.; Rush, C.M.; Bordovsky, D.; Clark, L.E.; OR, J. Effects of tillage on common root rot of wheat in Texas. Plant Dis. 1990, 74, 1006–1008. [Google Scholar] [CrossRef]
- Acharya, K.; Dutta, A.K.; Pradhan, P. Bipolaris sorokiniana (Sacc.) Shoem.: The most destructive wheat fungal pathogen in the warmer areas. Aust. J. Crop Sci. 2011, 5, 1064–1071. [Google Scholar] [CrossRef]
- Hill, J.P.; Fernandez, J.A.; McShane, M.S. Fungi associated with common root rot of winter wheat in Colorado and Wyoming. Plant Dis. 1983, 67, 795–797. [Google Scholar] [CrossRef]
- Fernandez, J.A.; Wofford, D.S.; Horton, J.L. Augmentation of wheat common root rot by Fusarium acuminatum. Mycopathologia 1985, 90, 177–179. [Google Scholar] [CrossRef]
- Fernandez, M.R.; Fox, S.L.; Hucl, P.; Singh, A.K.; Stevenson, F.C. Root rot severity and fungal populations in spring common, durum and spelt wheat, and Kamut grown under organic management in western Canada. Can. J. Plant Sci. 2014, 94, 937–946. [Google Scholar] [CrossRef]
- Gonzalez, M.S.; Trevathan, L.E. Identity and pathogenicity of fungi associated with root and crown rot of soft red winter wheat grown on the upper coastal plain land resource area of Mississippi. J. Phytopathol 2000, 148, 77–85. [Google Scholar] [CrossRef]
- Jevtić, R.; Stošić, N.; Župunski, V.; Lalošević, M.; Orbović, B. Variability of stem-base infestation and coexistence of Fusarium spp. causing crown rot of winter wheat in Serbia. J. Plant Pathol. 2019, 35, 553–563. [Google Scholar] [CrossRef]
- Özer, G.; İmren, M.; Paulitz, T.C.; Bayraktar, H.; Muminjanov, H.; Dababat, A.A. First Report of Crown Rot Caused by Fusarium algeriense on Wheat in Azerbaijan. Plant Dis. 2020, 104, 582. [Google Scholar] [CrossRef]
- Akinsanmi, O.A.; Mitter, V.; Simpfendorfer, S.; Backhouse, D.; Chakraborty, S. Identity and pathogenicity of Fusarium spp. isolated from wheat fields in Queensland and northern New South Wales. Aust. J. Agric. Res. 2004, 55, 97–107. [Google Scholar] [CrossRef]
- Backhouse, D.; Burgess, L.W. Climatic analysis of the distribution of Fusarium graminearum, F. pseudograminearum and F. culmorum on cereals in Australia. Australas. Plant Pathol. 2002, 31, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Özer, G.; İmren, M.; Alkan, M.; Paulitz, T.C.; Bayraktar, H.; PALACIOĞLU, G.; Dababat, A.A. Molecular and pathogenic characterization of Cochliobolus anamorphs associated with common root rot of wheat in Azerbaijan. Phytopathol. Mediterr. 2020, 59, 147–158. [Google Scholar] [CrossRef]
- Fernandez, M.R.; Chen, Y. Pathogenicity of Fusarium species on different plant parts of spring wheat under controlled conditions. Plant Dis. 2005, 89, 164–169. [Google Scholar] [CrossRef] [Green Version]
- Broders, K.D.; Parker, M.L.; Melzer, M.S.; Boland, G.J. Phylogenetic diversity of Rhizoctonia solani associated with canola and wheat in Alberta, Manitoba, and Saskatchewan. Plant Dis. 2014, 98, 1695–1701. [Google Scholar] [CrossRef] [Green Version]
- Jaaffar, A.K.M.; Paulitz, T.C.; Schroeder, K.L.; Thomashow, L.S.; Weller, D.M. Molecular characterization, morphological characteristics, virulence and geographic distribution of Rhizoctonia spp. in Washington State, USA. Phytopathology 2016, 106, 459–473. [Google Scholar] [CrossRef] [Green Version]
- Özer, G.; Göre, M.E.; İmren, M.; Khalilova, S.; Muminjanov, H.; Dababat, A.A. First report of Rhizoctonia solani AG-4 HGII and AG-2-1 causing root rot of wheat in Azerbaijan. Plant Dis. 2019, 103, 2132. [Google Scholar] [CrossRef]
- Melzer, M.S.; Yu, H.; Labun, T.; Dickson, A.; Boland, G.J. Characterization and pathogenicity of Rhizoctonia spp. from field crops in Canada. Can. J. Plant Pathol. 2016, 38, 367–374. [Google Scholar] [CrossRef]
- Taheri, A.E.; Hamel, C.; Gan, Y.; Vujanovic, V. First report of Fusarium redolens from Saskatchewan and its comparative pathogenicity. Can. J. Plant Pathol. 2011, 33, 559–564. [Google Scholar] [CrossRef]
- Shikur Gebremariam, E.; Karakaya, A.; Erginbas-Orakci, G.; Dababat, A.A.; Sharma-Poudyal, D.; Paulitz, T.C. First report of Fusarium redolens causing crown rot of wheat (Triticum spp.) in Turkey. Plant Dis. 2015, 99, 1280. [Google Scholar] [CrossRef]
- Specht, L.P.; Rush, C.M. Fungi associated with root and foot rot of winter wheat and populations of Cochliobolus sativus in the Texas Panhandle. Plant Dis. 1988, 72, 959–963. [Google Scholar] [CrossRef]
- Smiley, R.W.; Patterson, L.M. Pathogenic fungi associated with Fusarium foot rot of winter wheat in the semiarid Pacific Northwest. Plant Dis. 1996, 80, 944–949. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Karagandy | East Kazakhstan | Almaty | Total | ||||
|---|---|---|---|---|---|---|---|---|
| N | IF | N | IF | N | IF | N | IF | |
| Bipolaris sorokiniana | 222 | 43.61 | 136 | 43.31 | 189 | 47.49 | 547 * | 44.80 |
| Fusarium acuminatum | 158 | 31.04 | 39 | 12.42 | 52 | 13.07 | 249 | 20.39 |
| Fusarium equiseti | 26 | 5.11 | 48 | 15.29 | 50 | 12.56 | 124 | 10.16 |
| Curvularia spicifera | 62 | 12.18 | 17 | 5.41 | 14 | 3.52 | 93 | 7.62 |
| Fusarium culmorum | 14 | 2.75 | 22 | 7.01 | 22 | 5.53 | 58 | 4.75 |
| Fusarium oxysporum | 5 | 0.98 | 21 | 6.69 | 24 | 6.03 | 50 | 4.10 |
| Fusarium redolens | 4 | 0.79 | 19 | 6.05 | 6 | 1.51 | 29 | 2.38 |
| Rhizoctonia solani AG2-1 | 1 | 0.20 | 4 | 1.27 | 8 | 2.01 | 13 | 1.06 |
| Nigrospora oryzae | 3 | 0.59 | 3 | 0.96 | 6 | 1.51 | 12 | 0.98 |
| Curvularia inaequalis | 6 | 1.18 | 2 | 0.64 | 3 | 0.75 | 11 | 0.90 |
| Fusarium pseudograminearum | 2 | 0.39 | 2 | 0.64 | 5 | 1.26 | 9 | 0.74 |
| Fusarium flocciferum | 0 | 0.00 | 0 | 0.00 | 9 | 2.26 | 9 | 0.74 |
| Macrophomina phaseolina | 0 | 0.00 | 0 | 0.00 | 8 | 2.01 | 8 | 0.66 |
| Fusarium cf. incarnatum | 4 | 0.79 | 0 | 0.00 | 0 | 0.00 | 4 | 0.33 |
| Fusarium sp. | 1 | 0.20 | 0 | 0.00 | 2 | 0.50 | 3 | 0.25 |
| Fusarium torulosum | 1 | 0.20 | 1 | 0.32 | 0 | 0.00 | 2 | 0.16 |
| Total | 509 | 100 | 314 | 100 | 398 | 100 | 1221 | 100 |
| Species | Karagandy | East Kazakhstan | Almaty | Total | ||||
|---|---|---|---|---|---|---|---|---|
| N | FI | N | FI | N | FI | N | FI | |
| Bipolaris sorokiniana | 25 * | 96.15 | 13 | 72.22 | 18 | 85.71 | 56 | 86.15 |
| Fusarium acuminatum | 24 | 92.31 | 7 | 38.89 | 12 | 57.14 | 43 | 66.15 |
| Fusarium oxysporum | 5 | 19.23 | 10 | 55.56 | 12 | 57.14 | 27 | 41.54 |
| Fusarium equiseti | 7 | 26.92 | 8 | 44.44 | 8 | 38.10 | 23 | 35.38 |
| Curvularia spicifera | 11 | 42.31 | 4 | 22.22 | 3 | 14.29 | 18 | 27.69 |
| Fusarium culmorum | 4 | 15.38 | 4 | 22.22 | 4 | 19.05 | 12 | 18.46 |
| Rhizoctonia solani AG2-1 | 1 | 3.85 | 3 | 16.67 | 4 | 19.05 | 8 | 12.31 |
| Fusarium redolens | 2 | 7.69 | 2 | 11.11 | 2 | 9.52 | 6 | 9.23 |
| Nigrospora oryzae | 2 | 7.69 | 2 | 11.11 | 2 | 9.52 | 6 | 9.23 |
| Fusarium pseudograminearum | 1 | 3.85 | 1 | 5.56 | 3 | 14.29 | 5 | 7.69 |
| Curvularia inaequalis | 3 | 11.54 | 1 | 5.56 | 1 | 4.76 | 5 | 7.69 |
| Fusarium flocciferum | 0 | 0.00 | 0 | 0.00 | 3 | 14.29 | 3 | 4.62 |
| Fusarium torulosum | 1 | 3.85 | 1 | 5.56 | 0 | 0.00 | 2 | 3.08 |
| Fusarium sp. | 1 | 3.85 | 0 | 0.00 | 1 | 4.76 | 2 | 3.08 |
| Fusarium cf. incarnatum | 2 | 7.69 | 0 | 0.00 | 0 | 0.00 | 2 | 3.08 |
| Macrophomina phaseolina | 0 | 0.00 | 0 | 0.00 | 2 | 9.52 | 2 | 3.08 |
| Species | Number of Isolates | Average Severity Index * | Average Disease Severity (%) *** | Virulence Category **** |
|---|---|---|---|---|
| Fusarium pseudograminearum | 5 | 3.55 ± 0.60a ** | 71.09 | HV |
| Fusarium culmorum | 5 | 3.42 ± 0.64a | 68.37 | HV |
| Bipolaris sorokiniana | 5 | 2.83 ± 0.50b | 56.60 | MV |
| Fusarium sp. | 3 | 2.25 ± 0.61bc | 44.98 | MV |
| Rhizoctonia solani AG2-1 | 3 | 2.21 ± 0.54c | 44.12 | MV |
| Fusarium redolens | 4 | 2.20 ± 0.41c | 44.05 | MV |
| Curvularia spicifera | 5 | 1.94 ± 0.56cd | 38.87 | MiV |
| Curvularia ineaqualis | 5 | 1.60 ± 0.47de | 32.04 | MiV |
| Nigrospora oryzae | 5 | 1.56 ± 0.50de | 31.24 | MiV |
| Fusarium acuminatum | 5 | 1.37 ± 0.48e | – | NP |
| Fusarium oxysporum | 5 | 1.36 ± 0.52e | – | NP |
| Fusarium flocciferum | 5 | 1.29 ± 0.46e | – | NP |
| Control | 5 | 1.28 ± 0.41e | – | NP |
| Macrophomina phaseolina | 5 | 1.26 ± 0.43e | – | NP |
| Fusarium torulosum | 2 | 1.19 ± 0.29e | – | NP |
| Fusarium cf. incarnatum | 2 | 1.19 ± 0.29e | – | NP |
| Fusarium equiseti | 5 | 1.18 ± 0.37e | – | NP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bozoğlu, T.; Derviş, S.; Imren, M.; Amer, M.; Özdemir, F.; Paulitz, T.C.; Morgounov, A.; Dababat, A.A.; Özer, G. Fungal Pathogens Associated with Crown and Root Rot of Wheat in Central, Eastern, and Southeastern Kazakhstan. J. Fungi 2022, 8, 417. https://doi.org/10.3390/jof8050417
Bozoğlu T, Derviş S, Imren M, Amer M, Özdemir F, Paulitz TC, Morgounov A, Dababat AA, Özer G. Fungal Pathogens Associated with Crown and Root Rot of Wheat in Central, Eastern, and Southeastern Kazakhstan. Journal of Fungi. 2022; 8(5):417. https://doi.org/10.3390/jof8050417
Chicago/Turabian StyleBozoğlu, Tuğba, Sibel Derviş, Mustafa Imren, Mohammed Amer, Fatih Özdemir, Timothy C. Paulitz, Alexey Morgounov, Abdelfattah A. Dababat, and Göksel Özer. 2022. "Fungal Pathogens Associated with Crown and Root Rot of Wheat in Central, Eastern, and Southeastern Kazakhstan" Journal of Fungi 8, no. 5: 417. https://doi.org/10.3390/jof8050417
APA StyleBozoğlu, T., Derviş, S., Imren, M., Amer, M., Özdemir, F., Paulitz, T. C., Morgounov, A., Dababat, A. A., & Özer, G. (2022). Fungal Pathogens Associated with Crown and Root Rot of Wheat in Central, Eastern, and Southeastern Kazakhstan. Journal of Fungi, 8(5), 417. https://doi.org/10.3390/jof8050417