
Development of a Vector Set for High or Inducible Gene Expression and Protein Secretion in the Yeast Genus Blastobotrys


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Media
2.2. Genome Sequencing of A. siamense Strain L8in5 and Identification of GH43 AXH1 Gene
2.3. Accession Numbers
2.4. Promoters Cloning
2.5. Construction of Epitope Tagging and Secretion Plasmids
2.6. Heterologous Enzyme Cloning and Expression
2.7. Protocol for Blastobotrys spp. Transformation
2.8. Flow Cytometry
2.9. Microscopy
2.10. Protein Analysis
3. Results
3.1. B. raffinosifermentans Is Partially Equipped to Degrade Xylan
3.2. Growth Capacities of B. raffinosifermentans CBS 8335
3.3. Characterisation of the New Promoters on YEA Medium at 30 °C
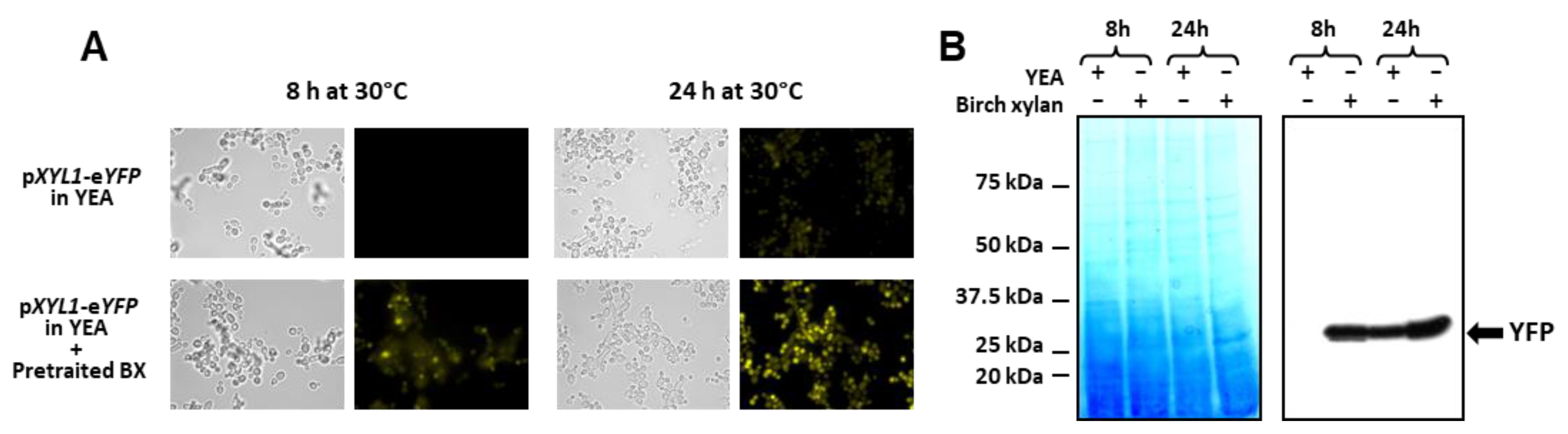
3.4. Characterisation of Inducible Promoters
3.5. Expression of Endogenous Epitope-Tagged Secretory Proteins
3.6. Expression of a Heterologous Glycosyl Hydrolase of the GH43 AXH-like Subgroup
3.7. Versatility of the Transformation/Expression Tools
3.8. A Xylanase Activity in B. yvelinesensis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muller, S.; Sandal, T.; Kamp-Hansen, P.; Dalboge, H. Comparison of expression systems in the yeasts Saccharomyces cerevisiae, Hansenula polymorpha, Klyveromyces lactis, Schizosaccharomyces pombe and Yarrowia lipolytica. Cloning of two novel promoters from Yarrowia lipolytica. Yeast 1998, 14, 1267–1283. [Google Scholar] [CrossRef]
- Gellissen, G.; Kunze, G.; Gaillardin, C.; Cregg, J.M.; Berardi, E.; Veenhuis, M.; van der Klei, I. New yeast expression platforms based on methylotrophic Hansenula polymorpha and Pichia pastoris and on dimorphic Arxula adeninivorans and Yarrowia lipolytica–a comparison. FEMS Yeast Res. 2005, 5, 1079–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boer, E.; Steinborn, G.; Matros, A.; Mock, H.P.; Gellissen, G.; Kunze, G. Production of interleukin-6 in Arxula adeninivorans, Hansenula polymorpha and Saccharomyces cerevisiae by applying the wide-range yeast vector (CoMed) system to simultaneous comparative assessment. FEMS Yeast Res. 2007, 7, 1181–1187. [Google Scholar] [CrossRef] [Green Version]
- Madhavan, A.; Arun, K.B.; Sindhu, R.; Krishnamoorthy, J.; Reshmy, R.; Sirohi, R.; Pugazhendi, A.; Awasthi, M.K.; Szakacs, G.; Binod, P. Customized yeast cell factories for biopharmaceuticals: From cell engineering to process scale up. Microb. Cell Fact 2021, 20, 124. [Google Scholar] [CrossRef]
- Wartmann, T.; Kruger, A.; Adler, K.; Duc, B.M.; Kunze, I.; Kunze, G. Temperature-dependent dimorphism of the yeast Arxula adeninivorans Ls3. Antonie Van Leeuwenhoek 1995, 68, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.X.; Wartmann, T.; Stoltenburg, R.; Kunze, G. Halotolerance of the yeast Arxula adeninivorans LS3. Antonie Van Leeuwenhoek 2000, 77, 303–311. [Google Scholar] [CrossRef]
- Gienow, U.; Kunze, G.; Schauer, F.; Bode, R.; Hofemeister, J. The yeast genus Trichosporon spec. LS3; molecular characterization of genomic complexity. Zentr. Mikrobiol. 1990, 145, 3–12. [Google Scholar] [CrossRef]
- Thomas, S.; Sanya, D.R.A.; Fouchard, F.; Nguyen, H.V.; Kunze, G.; Neuveglise, C.; Crutz-Le Coq, A.M. Blastobotrys adeninivorans and B. raffinosifermentans, two sibling yeast species which accumulate lipids at elevated temperatures and from diverse sugars. Biotechnol. Biofuels 2019, 12, 154. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Robnett, C.J. Multigene phylogenetic analysis of the Trichomonascus, Wickerhamiella and Zygoascus yeast clades, and the proposal of Sugiyamaella gen. nov. and 14 new species combinations. FEMS Yeast Res. 2007, 7, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Kunze, G.; Kunze, I. Arxula adeninivorans. In Non Conventional Yeasts in Biotechnology; Wolf, K., Ed.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 389–409. [Google Scholar]
- Wartmann, T.; Kunze, G. Genetic transformation and biotechnological application of the yeast Arxula adeninivorans. Appl. Microbiol. Biotechnol. 2000, 54, 619–624. [Google Scholar] [CrossRef]
- Rosel, H.; Kunze, G. Integrative transformation of the dimorphic yeast arxula adeninivorans LS3 based on hygromycin B resistance. Curr. Genet. 1998, 33, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Wartmann, T.; Rosel, H.; Kunze, I.; Bode, R.; Kunze, G. AILV1 gene from the yeast Arxula adeninivorans LS3—A new selective transformation marker. Yeast 1998, 14, 1017–1025. [Google Scholar] [CrossRef]
- Terentiev, Y.; Pico, A.H.; Boer, E.; Wartmann, T.; Klabunde, J.; Breuer, U.; Babel, W.; Suckow, M.; Gellissen, G.; Kunze, G. A wide-range integrative yeast expression vector system based on Arxula adeninivorans-derived elements. J. Ind. Microbiol. Biotechnol. 2004, 31, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Boer, E.; Piontek, M.; Kunze, G. Xplor 2-an optimized transformation/expression system for recombinant protein production in the yeast Arxula adeninivorans. Appl. Microbiol. Biotechnol. 2009, 84, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Boer, E.; Wartmann, T.; Luther, B.; Manteuffel, R.; Bode, R.; Gellissen, G.; Kunze, G. Characterization of the AINV gene and the encoded invertase from the dimorphic yeast Arxula adeninivorans. Antonie Van Leeuwenhoek 2004, 86, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Boer, E.; Breuer, F.S.; Weniger, M.; Denter, S.; Piontek, M.; Kunze, G. Large-scale production of tannase using the yeast Arxula adeninivorans. Appl. Microbiol. Biotechnol. 2011, 92, 105–114. [Google Scholar] [CrossRef]
- Bischoff, F.; Litwinska, K.; Cordes, A.; Baronian, K.; Bode, R.; Schauer, F.; Kunze, G. Three New Cutinases from the Yeast Arxula adeninivorans That Are Suitable for Biotechnological Applications. Appl. Environ. Microbiol. 2015, 81, 5497–5510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvaro-Benito, M.; Fernandez-Lobato, M.; Baronian, K.; Kunze, G. Assessment of Schwanniomyces occidentalis as a host for protein production using the wide-range Xplor2 expression platform. Appl. Microbiol. Biotechnol. 2013, 97, 4443–4456. [Google Scholar] [CrossRef] [Green Version]
- Kumari, A.; Baronian, K.; Kunze, G.; Gupta, R. Extracellular expression of YlLip11 with a native signal peptide from Yarrowia lipolytica MSR80 in three different yeast hosts. Protein Expr. Purif. 2015, 110, 138–144. [Google Scholar] [CrossRef]
- Pham, H.T.M.; Kunath, K.; Gehrmann, L.; Giersberg, M.; Tuerk, J.; Uhlig, S.; Hanke, G.; Simon, K.; Baronian, K.; Kunze, G. Application of modified Arxula adeninivorans yeast cells in an online biosensor for the detection of estrogenic compounds in wastewater samples. Sens. Actuators B-Chem. 2013, 185, 628–637. [Google Scholar] [CrossRef]
- Chamas, A.; Nieter, A.; Pham, H.T.; Giersberg, M.; Hettwer, K.; Uhlig, S.; Simon, K.; Baronian, K.; Kunze, G. Development of a recombinant Arxula adeninivorans cell bioassay for the detection of molecules with progesterone activity in wastewater. Anal. Bioanal. Chem. 2015, 407, 8109–8120. [Google Scholar] [CrossRef] [PubMed]
- Chamas, A.; Giersberg, M.; Friedrich, K.; Sonntag, F.; Kunze, D.; Uhlig, S.; Simon, K.; Baronian, K.; Kunze, G. Purification and immunodetection of the complete recombinant HER-2[neu] receptor produced in yeast. Protein Expr. Purif. 2015, 105, 61–70. [Google Scholar] [CrossRef]
- Buttner, R.; Bode, R.; Birnbaum, D. Alcoholic Fermentation of Starch by Arxula-Adeninivorans. Zbl. Mikrobiol. 1992, 147, 225–230. [Google Scholar] [CrossRef]
- Kunze, G.; Hähnel, U. Production of Butanol by Fermentation in Arxula sp. European Patent Application No. EP,2,508,597,A1, 10 October 2012. [Google Scholar]
- Rhee, M.S.; Wei, L.; Sawhney, N.; Kim, Y.S.; Rice, J.D.; Preston, J.F. Metabolic potential of Bacillus subtilis 168 for the direct conversion of xylans to fermentation products. Appl. Microbiol. Biotechnol. 2016, 100, 1501–1510. [Google Scholar] [CrossRef]
- Andlar, M.; Rezic, T.; Mardetko, N.; Kracher, D.; Ludwig, R.; Santek, B. Lignocellulose degradation: An overview of fungi and fungal enzymes involved in lignocellulose degradation. Eng. Life Sci. 2018, 18, 768–778. [Google Scholar] [CrossRef] [PubMed]
- Tabanag, I.D.F.; Tsai, S.-L. Hemicellulose degradation and utilization by a synthetic Saccharomyces cerevisiae consortium. bioRxiv 2018, 244. [Google Scholar] [CrossRef] [Green Version]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Ann. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef]
- van den Brink, J.; de Vries, R.P. Fungal enzyme sets for plant polysaccharide degradation. Appl. Microbiol. Biotechnol. 2011, 91, 1477–1492. [Google Scholar] [CrossRef] [Green Version]
- Polizeli, M.L.; Rizzatti, A.C.; Monti, R.; Terenzi, H.F.; Jorge, J.A.; Amorim, D.S. Xylanases from fungi: Properties and industrial applications. Appl. Microbiol. Biotechnol. 2005, 67, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Ogura, A.; Inui, M.; Tokuda, S.; Hosokawa, S.; Ihara, H.; Kasai, N. Identification of a GH62 alpha-L-arabinofuranosidase specific for arabinoxylan produced by Penicillium chrysogenum. Appl. Microbiol. Biotechnol. 2011, 90, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Chong, S.L.; Battaglia, E.; Coutinho, P.M.; Henrissat, B.; Tenkanen, M.; de Vries, R.P. The alpha-glucuronidase Agu1 from Schizophyllum commune is a member of a novel glycoside hydrolase family (GH115). Appl. Microbiol. Biotechnol. 2011, 90, 1323–1332. [Google Scholar] [CrossRef] [Green Version]
- Biely, P.; Mastihubova, M.; Tenkanen, M.; Eyzaguirre, J.; Li, X.L.; Vrsanska, M. Action of xylan deacetylating enzymes on monoacetyl derivatives of 4-nitrophenyl glycosides of beta-D-xylopyranose and alpha-L-arabinofuranose. J. Biotechnol. 2011, 151, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Jeffries, T.W.; Grigoriev, I.V.; Grimwood, J.; Laplaza, J.M.; Aerts, A.; Salamov, A.; Schmutz, J.; Lindquist, E.; Dehal, P.; Shapiro, H.; et al. Genome sequence of the lignocellulose-bioconverting and xylose-fermenting yeast Pichia stipitis. Nat. Biotechnol. 2007, 25, 319–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassa-Barbosa, L.A.; Procopio, R.E.; Matos, I.T.; Filho, S.A. Isolation and characterization of yeasts capable of efficient utilization of hemicellulosic hydrolyzate as the carbon source. Genet. Mol. Res. 2015, 14, 11605–11612. [Google Scholar] [CrossRef] [PubMed]
- Ravn, J.L.; Engqvist, M.K.M.; Larsbrink, J.; Geijer, C. CAZyme prediction in ascomycetous yeast genomes guides discovery of novel xylanolytic species with diverse capacities for hemicellulose hydrolysis. Biotechnol. Biofuels 2021, 14, 150. [Google Scholar] [CrossRef]
- Wartmann, T.; Bellebna, C.; Boer, E.; Bartelsen, O.; Gellissen, G.; Kunze, G. The constitutive AHSB4 promoter--a novel component of the Arxula adeninivorans-based expression platform. Appl. Microbiol. Biotechnol. 2003, 62, 528–535. [Google Scholar] [CrossRef]
- Partow, S.; Siewers, V.; Bjorn, S.; Nielsen, J.; Maury, J. Characterization of different promoters for designing a new expression vector in Saccharomyces cerevisiae. Yeast 2010, 27, 955–964. [Google Scholar] [CrossRef]
- Haslbeck, M.; Walke, S.; Stromer, T.; Ehrnsperger, M.; White, H.E.; Chen, S.; Saibil, H.R.; Buchner, J. Hsp26: A temperature-regulated chaperone. EMBO J. 1999, 18, 6744–6751. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Zhao, X.Q.; Chang, A.K.; Zhang, Q.M.; Bai, F.W. Ethanol-induced yeast flocculation directed by the promoter of TPS1 encoding trehalose-6-phosphate synthase 1 for efficient ethanol production. Metab. Eng. 2012, 14, 1–8. [Google Scholar] [CrossRef]
- Mhetras, N.; Liddell, S.; Gokhale, D. Purification and characterization of an extracellular beta-xylosidase from Pseudozyma hubeiensis NCIM 3574 (PhXyl), an unexplored yeast. Amb. Express 2016, 6, 73. [Google Scholar] [CrossRef] [Green Version]
- Valdes-Hevia, M.D.; de la Guerra, R.; Gancedo, C. Isolation and characterization of the gene encoding phosphoenolpyruvate carboxykinase from Saccharomyces cerevisiae. FEBS Lett. 1989, 258, 313–316. [Google Scholar] [CrossRef]
- Saubin, M.; Devillers, H.; Proust, L.; Brier, C.; Grondin, C.; Pradal, M.; Legras, J.L.; Neuveglise, C. Investigation of Genetic Relationships Between Hanseniaspora Species Found in Grape Musts Revealed Interspecific Hybrids with Dynamic Genome Structures. Front. Microbiol. 2019, 10, 2960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Kunze, G.; Gaillardin, C.; Czernicka, M.; Durrens, P.; Martin, T.; Boer, E.; Gabaldon, T.; Cruz, J.A.; Talla, E.; Marck, C.; et al. The complete genome of Blastobotrys (Arxula) adeninivorans LS3—A yeast of biotechnological interest. Biotechnol. Biofuels 2014, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Leplat, C.; Nicaud, J.M.; Rossignol, T. High-throughput transformation method for Yarrowia lipolytica mutant library screening. FEMS Yeast Res. 2015, 15, fov052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanya, D.R.A.; Onesime, D.; Kunze, G.; Neuveglise, C.; Crutz-Le Coq, A.M. The native acyltransferase-coding genes DGA1 and DGA2 affect lipid accumulation in Blastobotrys raffinosifermentans differently when overexpressed. FEMS Yeast Res. 2020, 20, foaa060. [Google Scholar] [CrossRef]
- Madzak, C.; Treton, B.; Blanchin-Roland, S. Strong hybrid promoters and integrative expression/secretion vectors for quasi-constitutive expression of heterologous proteins in the yeast Yarrowia lipolytica. J. Mol. Microbiol. Biotechnol. 2000, 2, 207–216. [Google Scholar]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sonderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Gupta, R.; Brunak, S. Prediction of glycosylation across the human proteome and the correlation to protein function. Pac. Symp. Biocomput. 2002, 7, 310–322. [Google Scholar]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F.; et al. MycoCosm portal: Gearing up for 1000 fungal genomes. Nucleic. Acids. Res. 2014, 42, D699–D704. [Google Scholar] [CrossRef]
- Boer, E.; Bode, R.; Mock, H.P.; Piontek, M.; Kunze, G. Atan1p-an extracellular tannase from the dimorphic yeast Arxula adeninivorans: Molecular cloning of the ATAN1 gene and characterization of the recombinant enzyme. Yeast 2009, 26, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Pons, T.; Naumoff, D.G.; Martinez-Fleites, C.; Hernandez, L. Three acidic residues are at the active site of a beta-propeller architecture in glycoside hydrolase families 32, 43, 62, and 68. Proteins 2004, 54, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Zeng, Y.; Tang, R.Q.; Alper, H.S.; Bai, F.W.; Zhao, X.Q. Condition-specific promoter activities in Saccharomyces cerevisiae. Microb. Cell Fact. 2018, 17, 58. [Google Scholar] [CrossRef] [PubMed]
- Nicaud, J.M.; Fabre, E.; Gaillardin, C. Expression of invertase activity in Yarrowia lipolytica and its use as a selective marker. Curr. Genet. 1989, 16, 253–260. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | LS3 Gene | Size (pb) 1 | Predicted Regulation |
|---|---|---|---|
| HSP26 | ARAD1D12166g | 460 | Induced by heat shock |
| PCK1 | ARAD1D32010g | 1107 | Repressed by glucose Induced by gluconeogenic substrate |
| TDH3 | ARAD1D16896g | 1153 | Constitutive |
| TEF1 | ARAD1C01892g | 300 | Constitutive |
| TPS1 | ARAD1C43846g | 636 | Induced by osmotic stress |
| XYL1 | ARAD1D50644g | 879 | Induced by xylobiose |
| GH Family/CE | Name (EC Number) | LS3 Gene | Conserved Domains | Yeast Homologues (% Identity with LS3) 1 |
|---|---|---|---|---|
| GH10/GH11 | XYN1 (EC 3.2.1.8) | - | pfam00331 | S. stipitis KAG2732737.1 (GH10) Su. lignohabitans XP_018736116.1 (GH10) Su. lignohabitans XP_018734939.1 (GH10) T. ciferrii KAA8917054.1 (GH10) |
| GH43 | ABN1 (EC 3.2.1.99) | ARAD1D18216g | cd18831 | T. ciferrii KAA8914121.1 (78%) |
| GH43 | AXH1 (EC 3.2.1.55) | - | cd18619 | - |
| GH67 | AGU1 (EC 3.2.1.139) | ARAD1D23848g | pfam07488 | T. ciferrii KAA8917114.1 (61%) and T. ciferrii KAA8917113.1 (67%) |
| GH3 | XYL1 (EC 3.2.1.37) | ARAD1D50644g | PLN03080 | Su. lignohabitans XP_018735137.1 (58%) |
| CUT08920.1 | T. ciferrii KAA8906613.1 (50%) | |||
| ARAD1C05676g | PLN03080 | Su. lignohabitans XP_018735137.1 (55%) | ||
| CUT08919.1 | T. ciferrii KAA8906613.1 (60%) | |||
| CE1 | FAE (EC 3.2.1.73) | ARAD1A06094g | pfam07519 | - |
| ARAD1A19822g | - |
| Name | CDS Used for BlastP | LS3 Homologue | Conserved Domains | Best Homologues to LS3 Protein (% Identity) |
|---|---|---|---|---|
| XR (Xylose reductase) | S. stipitis PICST_89614 (XP_001385181.1) | ARAD1C28094g | cd19115 | T. ciferrii KAA8897533.1 (79%) Su. lignohabitans XP_018737989.1 (74%) |
| XDH (Xylitol dehydrogenase) | S. stipitis PICST_86924 (XP_001386982.1) | ARAD1D37840g (CAG34729.1) | - | T. ciferrii KAA8903833.1 (64%) Su. lignohabitans XP_018736469.1 (60%) |
| XK (Xylulo kinase) | S. stipitis PICST_68734 (XP_001387325.2) | ARAD1C08800g | cd07776 | T. ciferrii KAA8897400.1 (59%) Su. lignohabitans XP_018737264.1 (59%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boisramé, A.; Neuvéglise, C. Development of a Vector Set for High or Inducible Gene Expression and Protein Secretion in the Yeast Genus Blastobotrys. J. Fungi 2022, 8, 418. https://doi.org/10.3390/jof8050418
Boisramé A, Neuvéglise C. Development of a Vector Set for High or Inducible Gene Expression and Protein Secretion in the Yeast Genus Blastobotrys. Journal of Fungi. 2022; 8(5):418. https://doi.org/10.3390/jof8050418
Chicago/Turabian StyleBoisramé, Anita, and Cécile Neuvéglise. 2022. "Development of a Vector Set for High or Inducible Gene Expression and Protein Secretion in the Yeast Genus Blastobotrys" Journal of Fungi 8, no. 5: 418. https://doi.org/10.3390/jof8050418
APA StyleBoisramé, A., & Neuvéglise, C. (2022). Development of a Vector Set for High or Inducible Gene Expression and Protein Secretion in the Yeast Genus Blastobotrys. Journal of Fungi, 8(5), 418. https://doi.org/10.3390/jof8050418