
The Penicillium brasilianum Histone Deacetylase Clr3 Regulates Secondary Metabolite Production and Tolerance to Oxidative Stress

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture Conditions
2.2. Genomic DNA Extraction
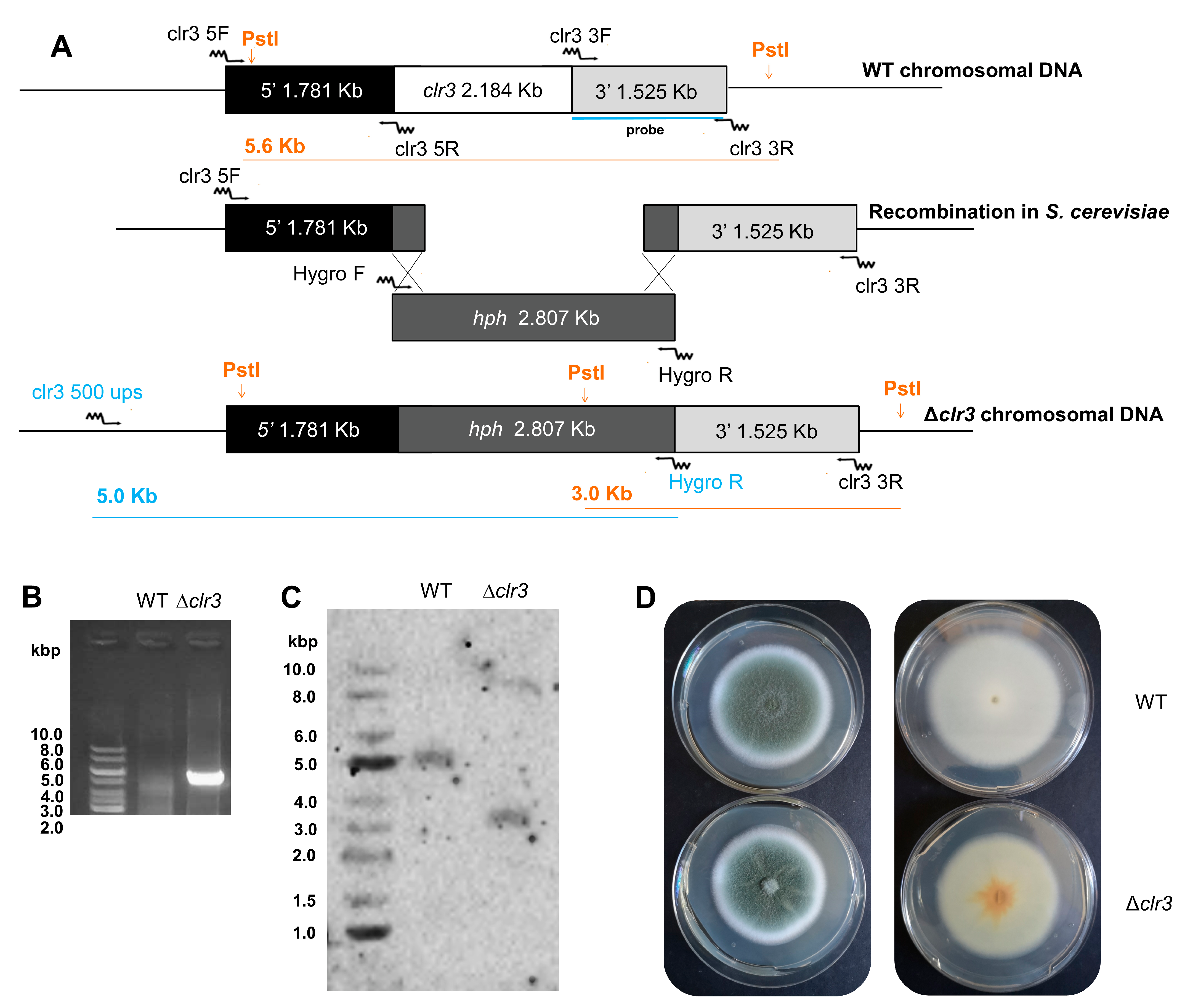
2.3. Construction of ∆clr3 Mutant
2.4. Southern Blot Analysis
2.5. Phenotypic Assays for Oxidative Stress Sensibility
2.6. RNA Extraction and RT-qPCR Analysis
2.7. In Silico Antioxidant Enzymes’ Subcellular Localization Prediction
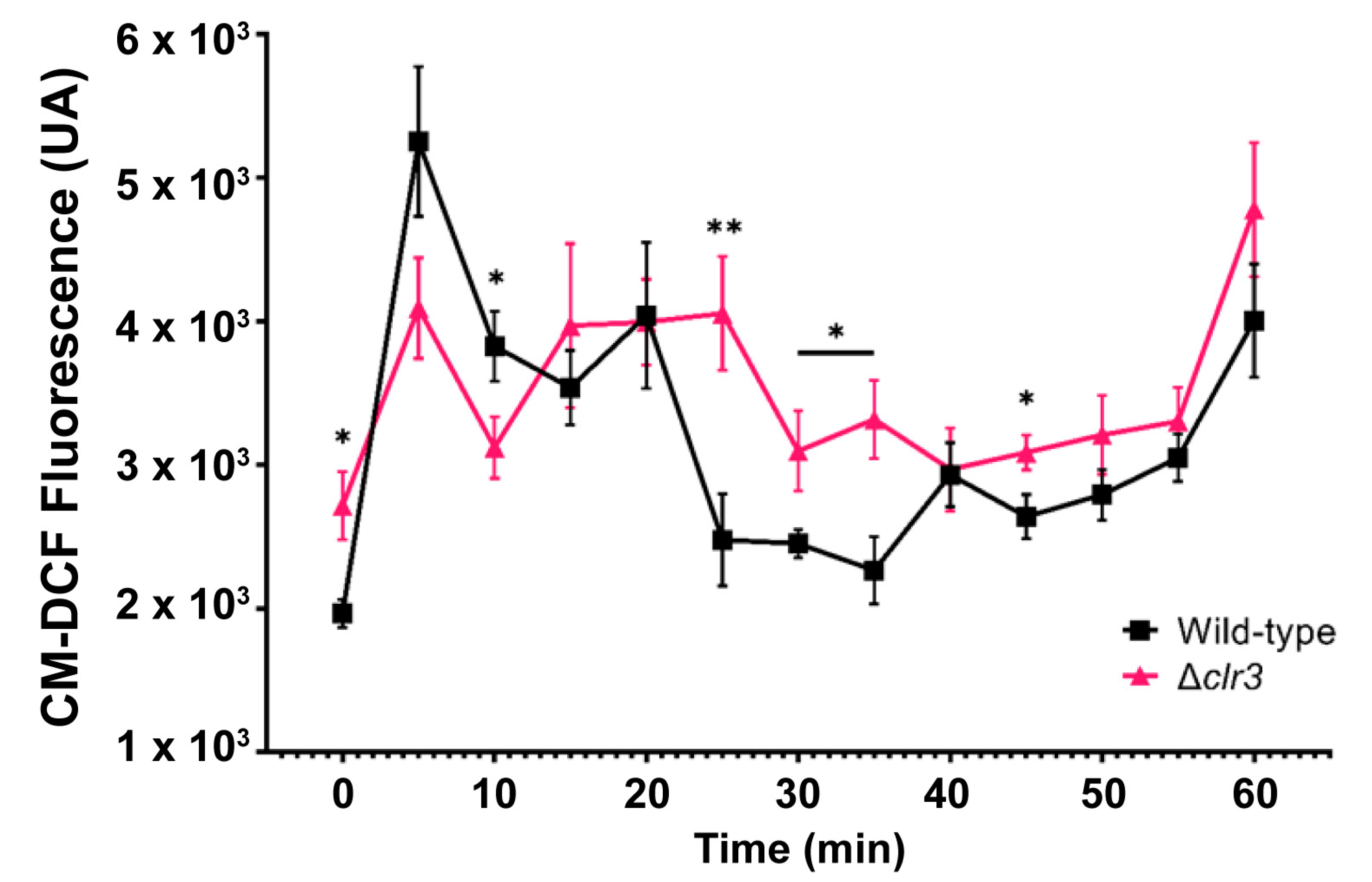
2.8. ROS Quantification
2.9. Secondary Metabolite Extraction
2.10. UPLC-DAD-MS Analyses
2.11. High-Resolution Mass Spectrometry Analyses
2.12. Chemical Epigenetic Modulation Experiments
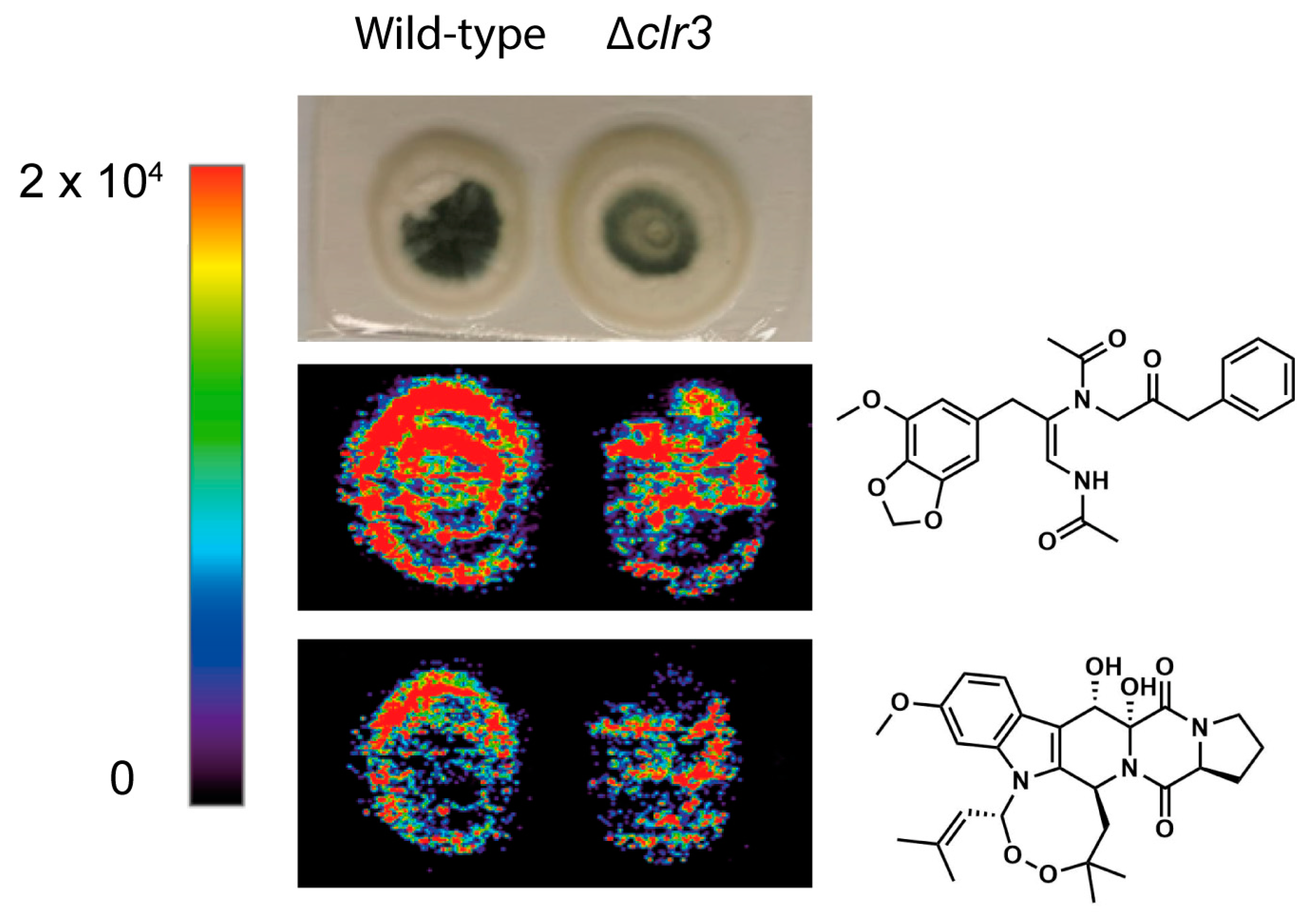
2.13. Mass Spectrometry Imaging (MSI)
3. Results and Discussion
3.1. Δclr3 Strain Construction and Phenotypic Analysis
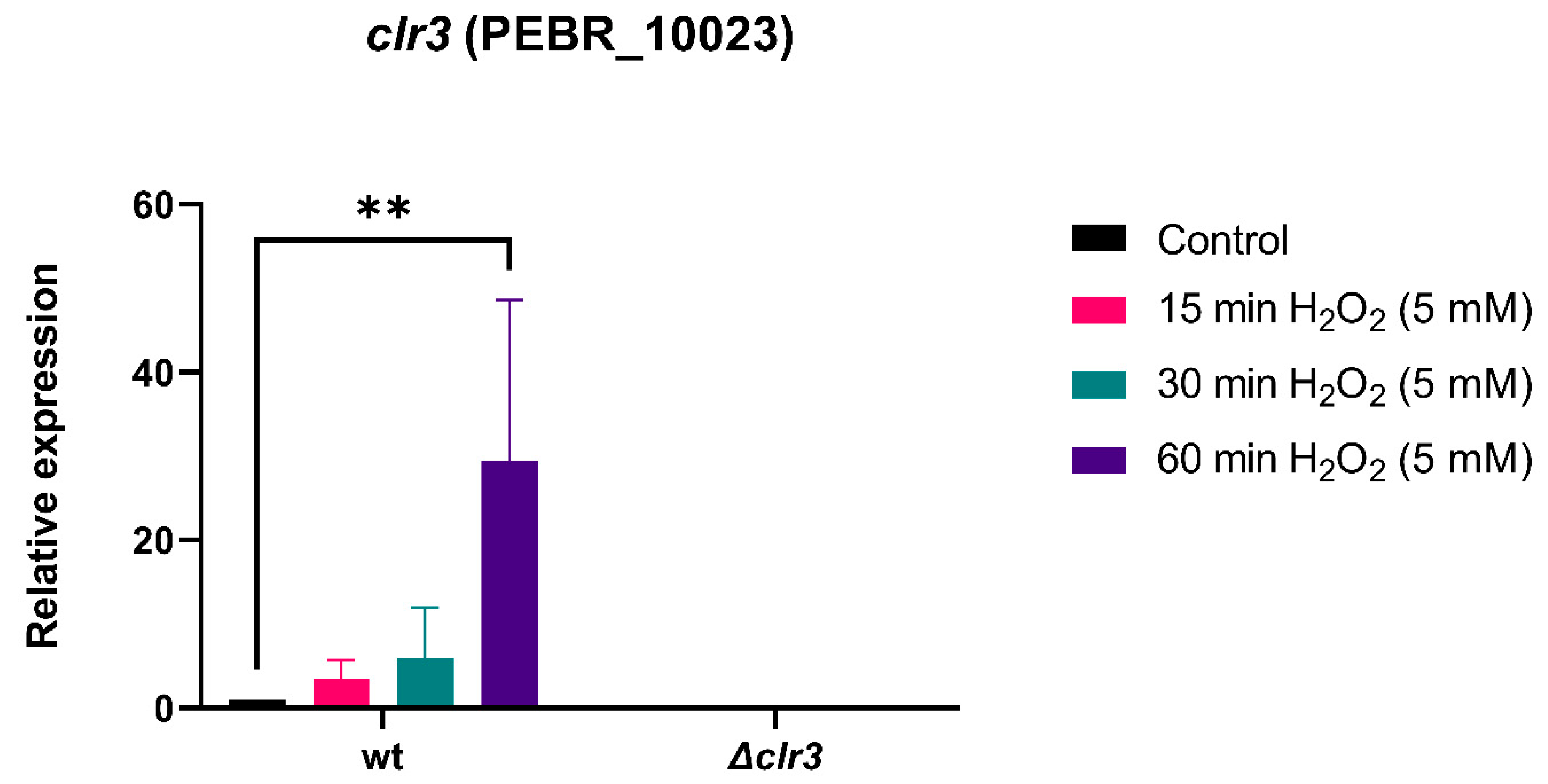
3.2. The Transcriptional Basis for P. brasilianum Δclr3 Sensitivity to Hydrogen Peroxide
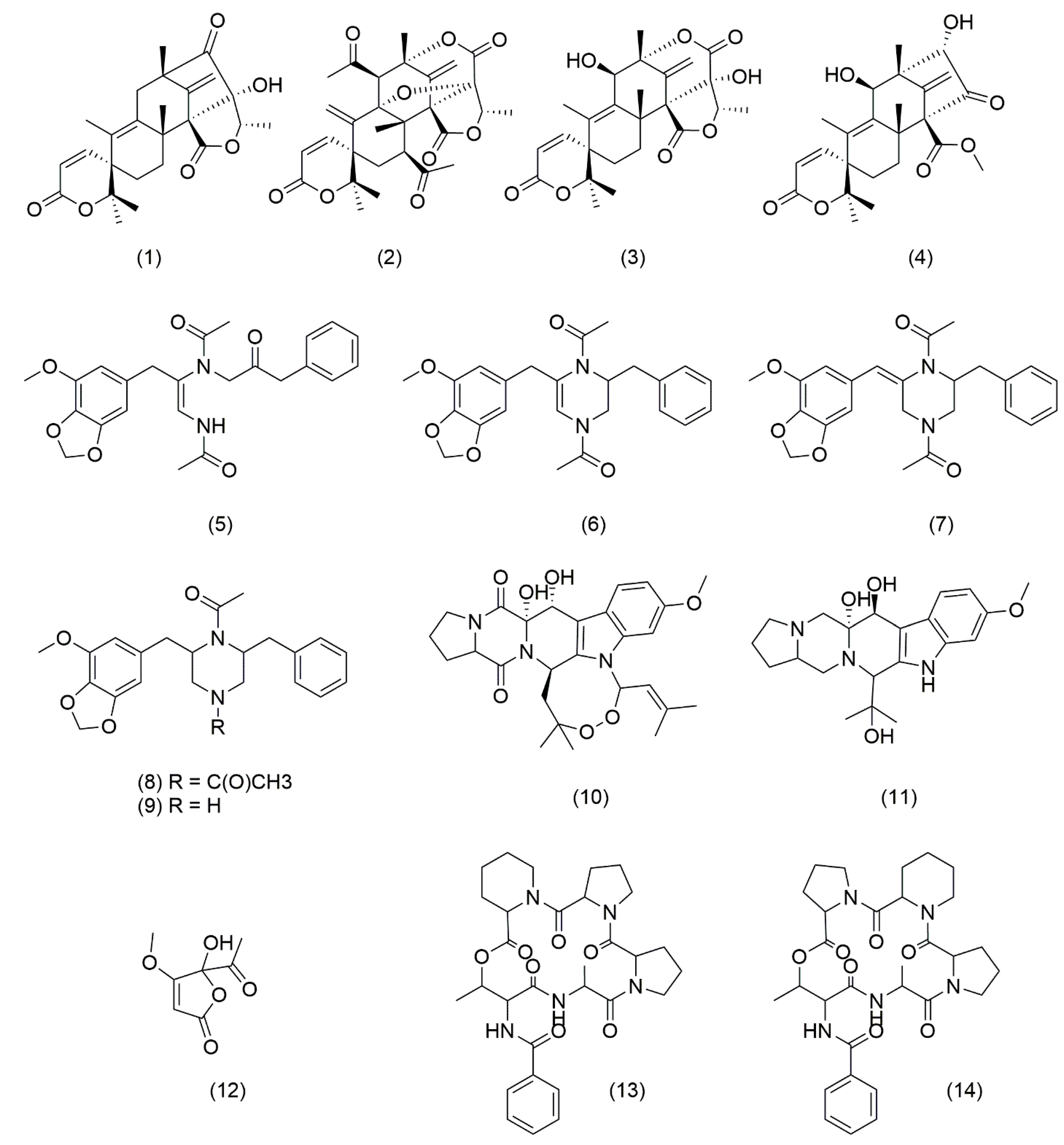
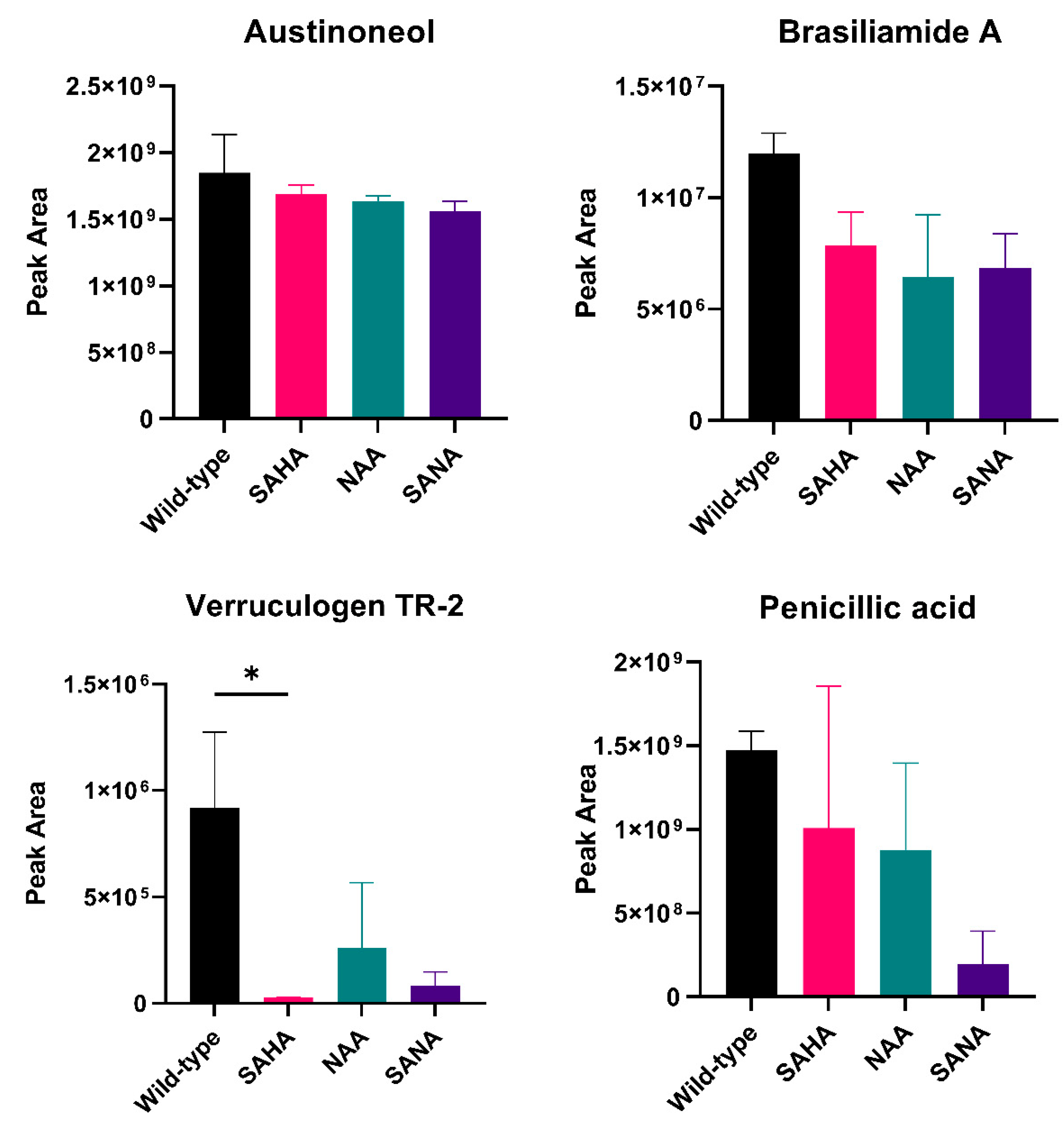
3.3. Natural Product Diversity in Δclr3 Strain
3.4. The Regulatory Role of HDACs in Penicillium brasilianum Secondary Metabolism
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, A.; Singh, D.K.; Kharwar, R.N.; White, J.F.; Gond, S.K. Fungal endophytes as efficient sources of plant-derived bioactive compounds and their prospective applications in natural product drug discovery: Insights, avenues, and challenges. Microorganisms 2021, 9, 197. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Li, S. Compartmentalized biosynthesis of fungal natural products. Curr. Opin. Biotechnol. 2021, 69, 128–135. [Google Scholar] [CrossRef]
- Bazioli, J.M.; Amaral, L.D.S.; Fill, T.P.; Rodrigues-Filho, E. Insights into Penicillium brasilianum Secondary Metabolism and Its Biotechnological Potential. Molecules 2017, 22, 858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fill, T.P.; da Silva, B.F.; Rodrigues-Fo, E. Biosynthesis of phenylpropanoid amides by an endophytic Penicillium brasilianum found in root bark of Melia azedarach. J. Microbiol. Biotechnol. 2010, 20, 622–629. [Google Scholar]
- Fujita, T.; Makishima, D.; Akiyama, K.; Hayashi, H. New convulsive compounds, brasiliamides A and B, from Penicillium brasilianum Batista JV-379. Biosci. Biotechnol. Biochem. 2002, 66, 1697–1705. [Google Scholar] [CrossRef] [Green Version]
- Fill, T.P.; Santos, R.M.G.d.; Barisson, A.; Rodrigues-Filho, E.; Souza, A.Q.L. Co-Production of Bisphenylpropanoid Amides and Meroterpenes by an Endophytic Penicillium brasilianum Found in the Root Bark of Melia azedarach. Z. Für Nat. C 2009, 64, 355. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.M.G.d.; Rodrigues-Filho, E. Structures of meroterpenes produced by Penicillium sp, an endophytic fungus found associated with Melia azedarach. J. Braz. Chem. Soc. 2003, 14, 722–727. [Google Scholar] [CrossRef] [Green Version]
- Geris dos Santos, R.M.; Rodrigues-Fo, E. Meroterpenes from Penicillium sp found in association with Melia azedarach. Phytochemistry 2002, 61, 907–912. [Google Scholar] [CrossRef]
- Inokoshi, J.; Nakamura, Y.; Hongbin, Z.; Uchida, R.; Nonaka, K.-I.; Masuma, R.; Tomoda, H. Spirohexalines, new inhibitors of bacterial undecaprenyl pyrophosphate synthase, produced by Penicillium brasilianum FKI-3368. J. Antibiot. 2013, 66, 37–41. [Google Scholar] [CrossRef]
- Hayashi, H.; Mukaihara, M.; Murao, S.; Arai, M.; Lee, A.Y.; Clardy, J. Acetoxydehydroaustin, a New Bioactive Compound, and Related Compound Neoaustin from Penicillium sp. MG–11. Biosci. Biotechnol. Biochem. 1994, 58, 334–338. [Google Scholar] [CrossRef]
- Fill, T.P.; Asenha, H.B.; Marques, A.S.; Ferreira, A.G.; Rodrigues-Fo, E. Time course production of indole alkaloids by an endophytic strain of Penicillium brasilianum cultivated in rice. Nat. Prod. Res. 2013, 27, 967–974. [Google Scholar] [CrossRef]
- Fill, T.P.; Baretta, J.F.; de Almeida, L.G.P.; Malavazi, I.; Cerdeira, L.T.; Samborskyy, M.; de Vasconcelos, A.T.R.; Leadlay, P.; Rodrigues-Filho, E. Draft Genome Sequence of the Fungus Penicillium brasilianum (Strain LaBioMMi 136), a Plant Endophyte from Melia azedarach. Microbiol. Resour. Announc. 2018, 7, e01235-18. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.-S.; Kim, E.-S. Recent advances in heterologous expression of natural product biosynthetic gene clusters in Streptomyces hosts. Curr. Opin. 2021, 69, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, Z.; Jia, J.; Du, T.; Zhang, N.; Tang, Y.; Fang, Y.; Fang, D. Overview of histone modification. In Histone Mutations and Cancer; Springer: Cham, Switzerland, 2021; pp. 1–16. [Google Scholar]
- Kayihan, D.S.; Aksoy, E.; Kayihan, C. Identification and expression profiling of toxic boron-responsive microRNAs and their targets in sensitive and tolerant wheat cultivars. Turk. J. Agric. For. 2021, 45, 411–433. [Google Scholar] [CrossRef]
- Lin, C.; Cao, X.; Qu, Z.; Zhang, S.; Naqvi, N.I.; Deng, Y.Z. The histone deacetylases MoRpd3 and MoHst4 regulate growth, conidiation, and pathogenicity in the rice blast fungus Magnaporthe oryzae. Msphere 2021, 6, e00118-21. [Google Scholar] [CrossRef]
- Li, C.-Y.; Chung, Y.-M.; Wu, Y.-C.; Hunyadi, A.; Wang, C.C.; Chang, F.-R. Natural products development under epigenetic modulation in fungi. Phytochem. Rev. 2020, 19, 1323–1340. [Google Scholar] [CrossRef]
- Malavazi, I.; Goldman, G.H. Gene disruption in Aspergillus fumigatus using a PCR-based strategy and in vivo recombination in yeast. Methods Mol. Biol. 2012, 845, 99–118. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual/Joseph Sambrook; Cold Spring Harbor: Long Island, NY, USA, 2001. [Google Scholar]
- Rocha, M.C.; Fabri, J.H.T.M.; Franco de Godoy, K.; Alves de Castro, P.; Hori, J.I.; Ferreira da Cunha, A.; Arentshorst, M.; Ram, A.F.; van den Hondel, C.A.; Goldman, G.H. Aspergillus fumigatus MADS-box transcription factor rlmA is required for regulation of the cell wall integrity and virulence. G3 Genes Genomes Genet. 2016, 6, 2983–3002. [Google Scholar] [CrossRef] [Green Version]
- Rocha, M.C.; de Godoy, K.F.; Bannitz-Fernandes, R.; Fabri, J.; Barbosa, M.M.F.; de Castro, P.A.; Almeida, F.; Goldman, G.H.; da Cunha, A.F.; Netto, L.E.S.; et al. Analyses of the three 1-Cys Peroxiredoxins from Aspergillus fumigatus reveal that cytosolic Prx1 is central to H(2)O(2) metabolism and virulence. Sci. Rep. 2018, 8, 12314. [Google Scholar] [CrossRef]
- Rocha, M.C.; De Godoy, K.F.; De Castro, P.A.; Hori, J.I.; Bom, V.L.P.; Brown, N.A.; Da Cunha, A.F.; Goldman, G.H.; Malavazi, I. The Aspergillus fumigatus pkcA G579R mutant is defective in the activation of the cell wall integrity pathway but is dispensable for virulence in a neutropenic mouse infection model. PLoS ONE 2015, 10, e0135195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruder Nascimento, A.C.M.d.O.; dos Reis, T.F.; de Castro, P.A.; Hori, J.I.; Bom, V.L.P.; de Assis, L.J.; Ramalho, L.N.Z.; Rocha, M.C.; Malavazi, I.; Brown, N.A. Mitogen activated protein kinases SakAHOG1 and MpkC collaborate for Aspergillus fumigatus virulence. Mol. Microbiol. 2016, 100, 841–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Smedsgaard, J. Micro-scale extraction procedure for standardized screening of fungal metabolite production in cultures. J. Chromatogr. A 1997, 760, 264–270. [Google Scholar] [CrossRef]
- Angolini, C.F.F.; Vendramini, P.H.; Araújo, F.D.S.; Araújo, W.L.; Augusti, R.; Eberlin, M.N.; de Oliveira, L.G. Direct Protocol for Ambient Mass Spectrometry Imaging on Agar Culture. Anal. Chem. 2015, 87, 6925–6930. [Google Scholar] [CrossRef] [PubMed]
- Brakhage, A.A.; Schroeckh, V. Fungal secondary metabolites—Strategies to activate silent gene clusters. Fungal Genet. Biol. 2011, 48, 15–22. [Google Scholar] [CrossRef]
- Shwab, E.K.; Bok, J.W.; Tribus, M.; Galehr, J.; Graessle, S.; Keller, N.P. Histone deacetylase activity regulates chemical diversity in Aspergillus. Eukaryot. Cell 2007, 6, 1656–1664. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K.; Izawa, M.; Nakajima, Y.; Jin, Q.; Hirose, T.; Nakamura, T.; Koshino, H.; Kanamaru, K.; Ohsato, S.; Kamakura, T.; et al. Increased metabolite production by deletion of an HDA1-type histone deacetylase in the phytopathogenic fungi, Magnaporthe oryzae (Pyricularia oryzae) and Fusarium asiaticum. Lett. Appl. Microbiol. 2017, 65, 446–452. [Google Scholar] [CrossRef]
- Studt, L.; Schmidt, F.J.; Jahn, L.; Sieber, C.M.K.; Connolly, L.R.; Niehaus, E.M.; Freitag, M.; Humpf, H.U.; Tudzynski, B. Two histone deacetylases, FfHda1 and FfHda2, are important for Fusarium fujikuroi secondary metabolism and virulence. Appl. Environ. Microbiol. 2013, 79, 7719–7734. [Google Scholar] [CrossRef] [Green Version]
- Guzman-Chavez, F.; Salo, O.; Samol, M.; Ries, M.; Kuipers, J.; Bovenberg, R.A.L.; Vreeken, R.J.; Driessen, A.J.M. Deregulation of secondary metabolism in a histone deacetylase mutant of Penicillium chrysogenum. Microbiologyopen 2018, 7, e00598. [Google Scholar] [CrossRef] [Green Version]
- Khochbin, S.; Verdel, A.; Lemercier, C.; Seigneurin-Berny, D. Functional significance of histone deacetylase diversity. Curr. Opin. Genet. Dev. 2001, 11, 162–166. [Google Scholar] [CrossRef] [Green Version]
- Tribus, M.; Galehr, J.; Trojer, P.; Brosch, G.; Loidl, P.; Marx, F.; Haas, H.; Graessle, S. HdaA, a major class 2 histone deacetylase of Aspergillus nidulans, affects growth under conditions of oxidative stress. Eukaryot. Cell 2005, 4, 1736–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.; Oh, J.-H.; Shwab, E.K.; Dagenais, T.R.T.; Andes, D.; Keller, N.P. HdaA, a class 2 histone deacetylase of Aspergillus fumigatus, affects germination and secondary metabolite production. Fungal Genet. Biol. 2009, 46, 782–790. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, L.; Wysong, D.; Diamond, R.; Aguirre, J. Two divergent catalase genes are differentially regulated during Aspergillus nidulans development and oxidative stress. J. Bacteriol. 1997, 179, 3284–3292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dicker, E.; Cederbaum, A.I. NADH-dependent generation of reactive oxygen species by microsomes in the presence of iron and redox cycling agents. Biochem. Pharmacol. 1991, 42, 529–535. [Google Scholar] [CrossRef]
- Moye-Rowley, W.S. Regulation of the transcriptional response to oxidative stress in fungi: Similarities and differences. Eukaryot. Cell 2003, 2, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Sang, M.K.; Han, G.D.; Oh, J.Y.; Chun, S.-C.; Kim, K.D. Penicillium brasilianum as a novel pathogen of onion (Allium cepa L.) and other fungi predominant on market onion in Korea. Crop Prot. 2014, 65, 138–142. [Google Scholar] [CrossRef]
- Osorio-Concepción, M.; Cristóbal-Mondragón, G.R.; Gutiérrez-Medina, B.; Casas-Flores, S. Histone Deacetylase HDA-2 Regulates Trichoderma atroviride Growth, Conidiation, Blue Light Perception, and Oxidative Stress Responses. Appl. Environ. Microbiol. 2017, 83, e02922-16. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.G. Overview on peroxiredoxin. Mol. Cells 2016, 39, 1–5. [Google Scholar]
- Lessing, F.; Kniemeyer, O.; Wozniok, I.; Loeffler, J.; Kurzai, O.; Haertl, A.; Brakhage, A.A. The Aspergillus fumigatus transcriptional regulator AfYap1 represents the major regulator for defense against reactive oxygen intermediates but is dispensable for pathogenicity in an intranasal mouse infection model. Eukaryot. Cell 2007, 6, 2290–2302. [Google Scholar] [CrossRef] [Green Version]
- Lambou, K.; Lamarre, C.; Beau, R.; Dufour, N.; Latge, J.P. Functional analysis of the superoxide dismutase family in Aspergillus fumigatus. Mol. Microbiol. 2010, 75, 910–923. [Google Scholar] [CrossRef]
- Paris, S.; Wysong, D.; Debeaupuis, J.-P.; Shibuya, K.; Philippe, B.; Diamond, R.D.; Latgé, J.-P. Catalases of Aspergillus fumigatus. Infect. Immun. 2003, 71, 3551–3562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, R.L.; Williams, J.A.; Stadtman, E.P.; Shacter, E. [37] Carbonyl assays for determination of oxidatively modified proteins. Methods Enzymol. 1994, 233, 346–357. [Google Scholar] [PubMed]
- Chexal, K.K.; Spinger, J.P.; Clardy, J.; Cole, R.J.; Kirksey, J.W.; Dorner, J.W.; Cutler, H.G.; Strawter, B.J. Austin, a novel polyisoprenoid mycotoxin from Aspergillus ustus. J. Am. Chem. Soc. 1976, 98, 6748. [Google Scholar] [CrossRef]
- Geris, R.; Rodrigues-Fo, E.; Garcia da Silva, H.H.; Garcia da Silva, I. Larvicidal Effects of Fungal Meroterpenoids in the Control of Aedes aegypti L., the Main Vector of Dengue and Yellow Fever. Chem. Biodivers. 2008, 5, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T.; Hayashi, H. New brasiliamide congeners, brasiliamides C, D and E, from Penicillium brasilianum Batista JV-379. Biosci. Biotechnol. Biochem. 2004, 68, 820–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knaus, H.-G.; McManus, O.B.; Lee, S.H.; Schmalhofer, W.A.; Garcia-Calvo, M.; Helms, L.M.H.; Sanchez, M.; Giangiacomo, K.; Reuben, J.P. Tremorgenic Indole Alkaloids Potently Inhibit Smooth Muscle High-Conductance Calcium-Activated Potassium Channels. Biochemistry 1994, 33, 5819–5828. [Google Scholar] [CrossRef]
- Schürmann, B.T.M.; Sallum, W.S.T.; Takahashi, J.A. Austin, dehydroaustin and other metabolites from Penicillium brasilianum. Química Nova 2010, 33, 1044–1046. [Google Scholar] [CrossRef] [Green Version]
- Fill, T.P.; Pallini, H.F.; Amaral, L.d.S.; Silva, J.V.d.; Bidóia, D.L.; Peron, F.; Garcia, F.P.; Nakamura, C.V.; Rodrigues-Filho, E. Copper and Manganese Cations Alter Secondary Metabolism in the Fungus Penicillium brasilianum. J. Braz. Chem. Soc. 2016, 27, 1444–1451. [Google Scholar]
- Albright, J.C.; Henke, M.T.; Soukup, A.A.; McClure, R.A.; Thomson, R.J.; Keller, N.P.; Kelleher, N.L. Large-scale metabolomics reveals a complex response of Aspergillus nidulans to epigenetic perturbation. ACS Chem. Biol. 2015, 10, 1535–1541. [Google Scholar] [CrossRef] [Green Version]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef]
- Henrikson, J.C.; Hoover, A.R.; Joyner, P.M.; Cichewicz, R.H. A chemical epigenetics approach for engineering the in situ biosynthesis of a cryptic natural product from Aspergillus niger. Org. Biomol. Chem. 2009, 7, 435–438. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Fang, H.; Yin, C.; Wei, C.; Hu, J.; Zhang, Y. Antimicrobial Metabolites Produced by Penicillium mallochii CCH01 Isolated From the Gut of Ectropis oblique, Cultivated in the Presence of a Histone Deacetylase Inhibitor. Front. Microbiol. 2019, 10, 2186. [Google Scholar] [CrossRef] [PubMed]
- Pfannenstiel, B.T.; Greco, C.; Sukowaty, A.T.; Keller, N.P. The epigenetic reader SntB regulates secondary metabolism, development and global histone modifications in Aspergillus flavus. Fungal Genet. Biol. 2018, 120, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.P. Translating biosynthetic gene clusters into fungal armor and weaponry. Nat. Chem. Biol. 2015, 11, 671–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° | Molecule | Ion Formula ([M+H]+) | Calculated m/z ([M+H]+) | Experimental m/z ([M+H]+) | Error (ppm) | Class |
|---|---|---|---|---|---|---|
| 1 | Isoaustinone | C25H31O6 | 427.2115 | 427.2116 | 0.15 | Meroterpenoid |
| 2 | Acetoxydehydroaustin | C29H33O11 | 557.2017 | 557.2017 | −0.03 | Meroterpenoid |
| 3 | Austinol | C25H31O7 | 443.2064 | 443.2064 | −0.09 | Meroterpenoid |
| 4 | Austinoneol | C24H31O6 | 415.2115 | 415.2115 | 0.08 | Meroterpenoid |
| 5 | Brasiliamide A | C24H27N2O6 | 439.1864 | 439.1865 | −0.05 | Bisphenylpropanoid amides |
| 6 | Brasiliamide B | C24H27N2O5 | 423.1914 | 423.1914 | −0.09 | Bisphenylpropanoid amides |
| 7 | Brasiliamide C | C24H27N2O5 | 423.1914 | 423.1915 | 0.06 | Bisphenylpropanoid amides |
| 8 | Brasiliamide D | C24H29N2O5 | 425.2071 | 425.2070 | −0.16 | Bisphenylpropanoid amides |
| 9 | Brasiliamide E | C22H27N2O4 | 383.1965 | 383.1966 | 0.24 | Bisphenylpropanoid amides |
| 10 | Verruculogen * | C27H32N3O6 | 494.2286 | 494.2287 | 0.28 | Diketopiperazines |
| 11 | Verruculogen TR-2 | C22H28N3O6 | 430.1973 | 430.1969 | −0.84 | Diketopiperazines |
| 12 | Penicillic acid | C8H11O4 | 171.0652 | 171.0652 | 0.20 | Polyketide |
| 13 | JBIR 114 | C30H40N5O7 | 582.2922 | 582.2922 | −0.08 | Cyclodepsipeptides |
| 14 | JBIR 115 | C30H40N5O7 | 582.2922 | 582.2922 | −0.08 | Cyclodepsipeptides |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akiyama, D.Y.; Rocha, M.C.; Costa, J.H.; Teles, C.B.; da Silva Zuccoli, G.; Malavazi, I.; Fill, T.P. The Penicillium brasilianum Histone Deacetylase Clr3 Regulates Secondary Metabolite Production and Tolerance to Oxidative Stress. J. Fungi 2022, 8, 514. https://doi.org/10.3390/jof8050514
Akiyama DY, Rocha MC, Costa JH, Teles CB, da Silva Zuccoli G, Malavazi I, Fill TP. The Penicillium brasilianum Histone Deacetylase Clr3 Regulates Secondary Metabolite Production and Tolerance to Oxidative Stress. Journal of Fungi. 2022; 8(5):514. https://doi.org/10.3390/jof8050514
Chicago/Turabian StyleAkiyama, Daniel Yuri, Marina Campos Rocha, Jonas Henrique Costa, Caroline Brandão Teles, Giuliana da Silva Zuccoli, Iran Malavazi, and Taicia Pacheco Fill. 2022. "The Penicillium brasilianum Histone Deacetylase Clr3 Regulates Secondary Metabolite Production and Tolerance to Oxidative Stress" Journal of Fungi 8, no. 5: 514. https://doi.org/10.3390/jof8050514
APA StyleAkiyama, D. Y., Rocha, M. C., Costa, J. H., Teles, C. B., da Silva Zuccoli, G., Malavazi, I., & Fill, T. P. (2022). The Penicillium brasilianum Histone Deacetylase Clr3 Regulates Secondary Metabolite Production and Tolerance to Oxidative Stress. Journal of Fungi, 8(5), 514. https://doi.org/10.3390/jof8050514