
Host–Pathogen Interactions between Metarhizium spp. and Locusts


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
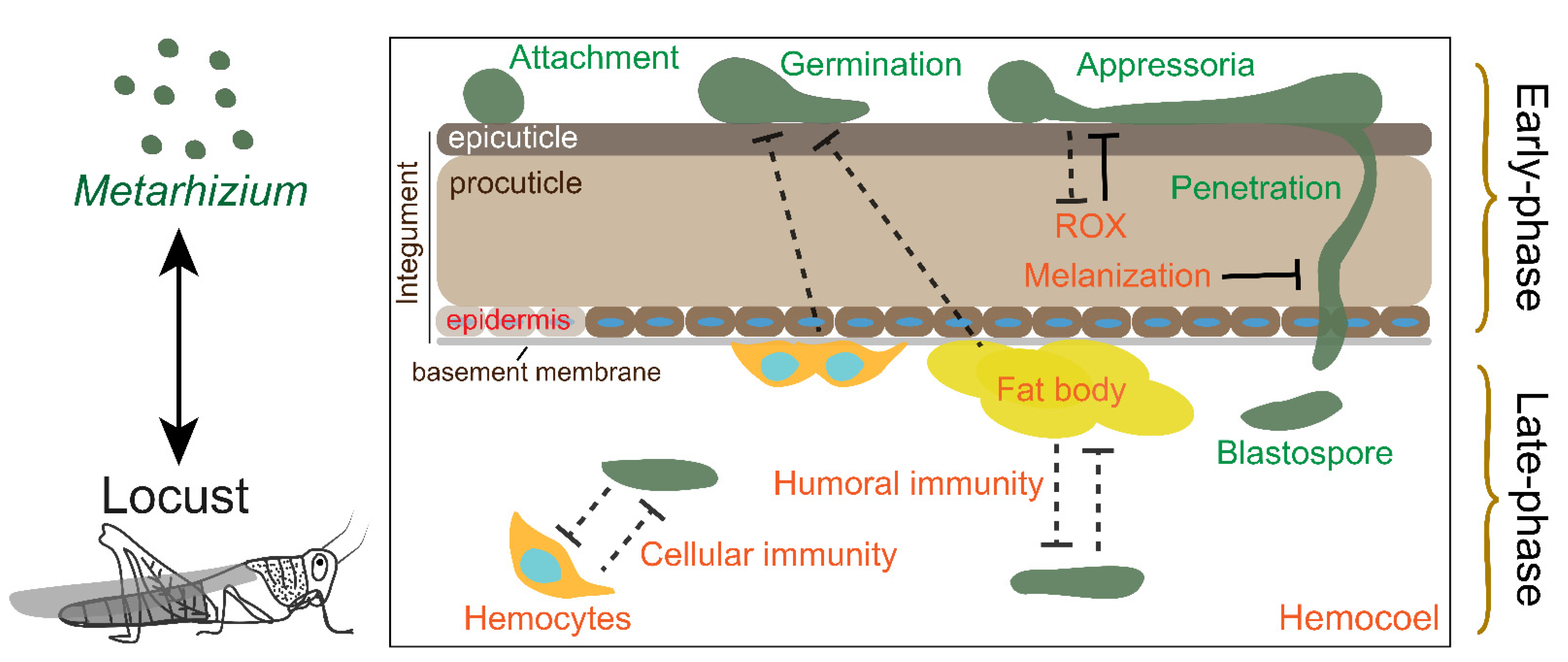
2. The Early Phase Interaction
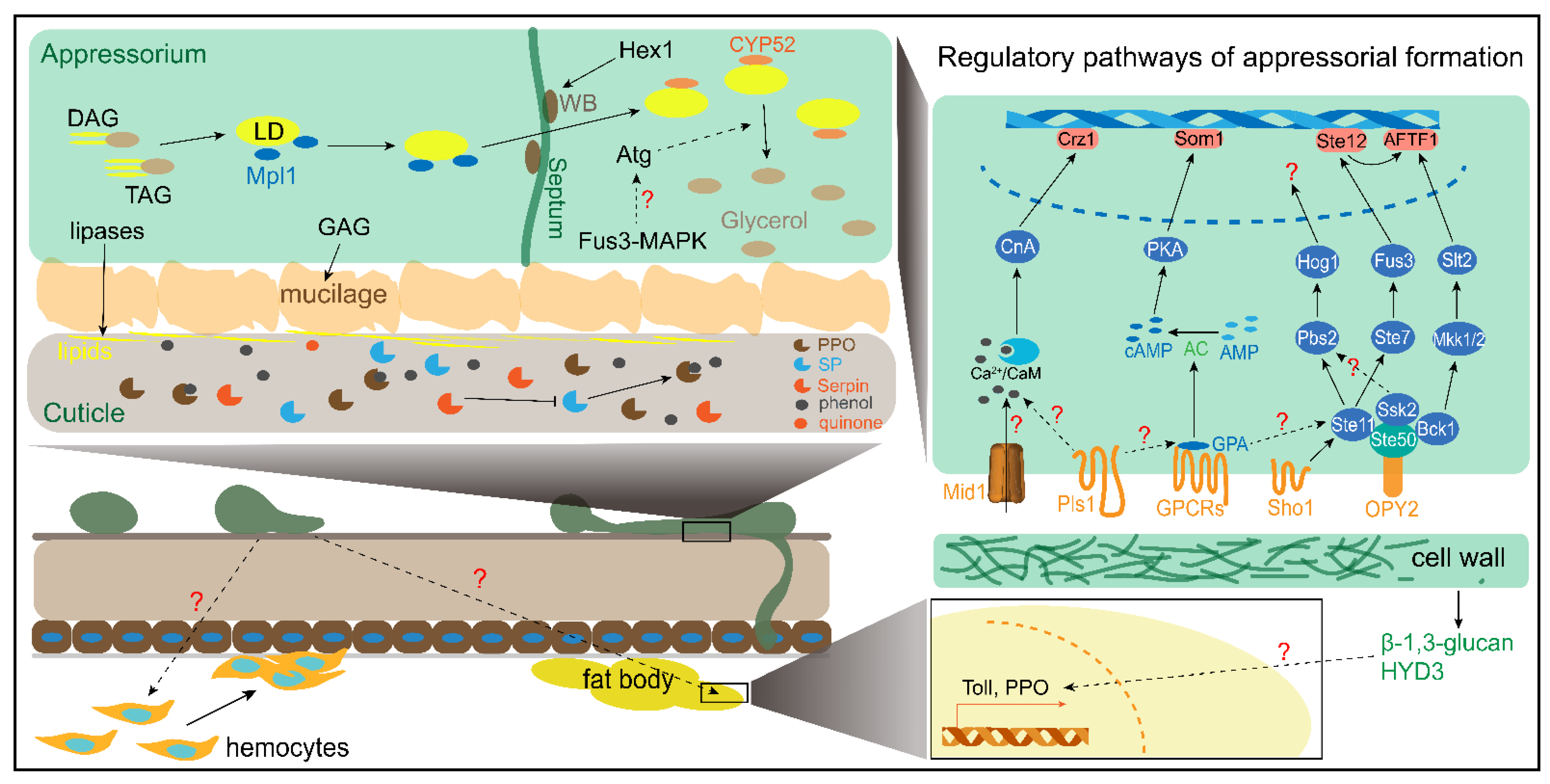
2.1. Attachment
2.2. Appressorial Formation
2.3. Penetration
2.4. The Locust Early Phase Immunity
3. The Late-Phase Interaction
3.1. Colonization of Metarhizium in Hemocoel
3.2. Humoral Response at Late-Phase Immunity
3.3. Cellular Response at Late-Phase Immunity
4. Locust Behavioral Fever and Density-Dependent Disease Resistance
4.1. Behavioral Fever
4.2. Density-Dependent Disease Resistance
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, C.; Wang, S. Insect Pathogenic Fungi: Genomics, Molecular Interactions, and Genetic Improvements. Annu. Rev. Entomol. 2017, 62, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Vega, F.E.; Meyling, N.V.; Luangsa-Ard, J.J.; Blackwell, M. Fungal Entomopathogens. In Insect Pathology, 2nd ed.; Vega, F., Kaya, H., Eds.; Academic: San Diego, CA, USA, 2012; pp. 171–220. [Google Scholar]
- Hu, X.; Xiao, G.; Zheng, P.; Shang, Y.; Su, Y.; Zhang, X.; Liu, X.; Zhan, S.; St Leger, R.J.; Wang, C. Trajectory and genomic determinants of fungal-pathogen speciation and host adaptation. Proc. Natl. Acad. Sci. USA 2014, 111, 16796–16801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St Leger, R.J.; Wang, J.B. Metarhizium: Jack of all trades, master of many. Open Biol. 2020, 10, 200307. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, X.; Xu, C.; Zhao, H.; Zhang, X.; Zeng, G.; Qian, Y.; Liu, R.; Guo, N.; Mi, W.; et al. Horizontal gene transfer allowed the emergence of broad host range entomopathogens. Proc. Natl. Acad. Sci. USA 2019, 116, 7982–7989. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Fang, W.; Wang, C.; Leger, R.J.S. Insertion of an Esterase Gene into a Specific Locust Pathogen (Metarhizium acridum) Enables It to Infect Caterpillars. PLoS Pathog. 2011, 7, e1002097. [Google Scholar] [CrossRef] [Green Version]
- Peng, G.X.; Xie, J.Q.; Guo, R.; Keyhani, N.O.; Zeng, D.Y.; Yang, P.Y.; Xia, Y.X. Long-term field evaluation and large-scale application of a Metarhizium anisopliae strain for controlling major rice pests. J. Pest. Sci. 2021, 94, 969–980. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Lopes, R.B.; Delalibera, I., Jr.; Fernandes, E.K.K.; Luz, C.; Faria, M. Current status and perspectives of fungal entomopathogens used for microbial control of arthropod pests in Brazil. J. Invertebr. Pathol. 2019, 165, 46–53. [Google Scholar] [CrossRef]
- Peng, G.X.; Wang, Z.K.; Yin, Y.P.; Zeng, D.Y.; Xia, Y.X. Field trials of Metarhizium anisopliae var. acridum (Ascomycota: Hypocreales) against oriental migratory locusts, Locusta migratoria manilensis (Meyen) in Northern China. Crop. Prot. 2008, 27, 1244–1250. [Google Scholar] [CrossRef]
- Hochkirch, A.; Bhaskar, D. Minimize collateral damage in locust control. Science 2021, 371, 1214–1215. [Google Scholar] [CrossRef]
- Guo, X.; Yu, Q.; Chen, D.; Wei, J.; Yang, P.; Yu, J.; Wang, X.; Kang, L. 4-Vinylanisole is an aggregation pheromone in locusts. Nature 2020, 584, 584–588. [Google Scholar] [CrossRef]
- Wang, X.; Kang, L. Molecular mechanisms of phase change in locusts. Annu. Rev. Entomol. 2014, 59, 225–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, K.; Thomas, M.B.; Blanford, S.; Doggett, M.; Simpson, S.J.; Moore, S.L. Coping with crowds: Density-dependent disease resistance in desert locusts. Proc. Natl. Acad. Sci. USA 2002, 99, 5471–5475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliot, S.L.; Blanford, S.; Horton, C.M.; Thomas, M.B. Fever and phenotype: Transgenerational effect of disease on desert locust phase state. Ecol. Lett 2003, 6, 830–836. [Google Scholar] [CrossRef]
- Barelli, L.; Padilla-Guerrero, I.E.; Bidochka, M.J. Differential expression of insect and plant specific adhesin genes, Mad1 and Mad2, in Metarhizium robertsii. Fungal Biol. 2011, 115, 1174–1185. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.B.; Szilvay, G.R.; Nakari-Setala, T.; Penttila, M.E. Hydrophobins: The protein-amphiphiles of filamentous fungi. FEMS Microbiol. Rev. 2005, 29, 877–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Xia, Y.X.; Kim, B.; Keyhani, N.O. Two hydrophobins are involved in fungal spore coat rodlet layer assembly and each play distinct roles in surface interactions, development and pathogenesis in the entomopathogenic fungus, Beauveria bassiana. Mol. Microbiol. 2011, 80, 811–826. [Google Scholar] [CrossRef]
- Fang, W.; Pei, Y.; Bidochka, M.J. A regulator of a G protein signalling (RGS) gene, cag8, from the insect-pathogenic fungus Metarhizium anisopliae is involved in conidiation, virulence and hydrophobin synthesis. Microbiology 2007, 153, 1017–1025. [Google Scholar] [CrossRef] [Green Version]
- Sevim, A.; Donzelli, B.G.; Wu, D.; Demirbag, Z.; Gibson, D.M.; Turgeon, B.G. Hydrophobin genes of the entomopathogenic fungus, Metarhizium brunneum, are differentially expressed and corresponding mutants are decreased in virulence. Curr. Genet. 2012, 58, 79–92. [Google Scholar] [CrossRef]
- Su, X.; Yan, X.; Chen, X.; Guo, M.; Xia, Y.; Cao, Y. Calcofluor white hypersensitive proteins contribute to stress tolerance and pathogenicity in entomopathogenic fungus, Metarhizium acridum. Pest. Manag. Sci. 2021, 77, 1915–1924. [Google Scholar] [CrossRef]
- Chen, X.; Liu, Y.Z.; Keyhani, N.O.; Xia, Y.X.; Cao, Y.Q. The regulatory role of the transcription factor Crz1 in stress tolerance, pathogenicity, and its target gene expression in Metarhizium acridum. Appl. Microbiol. Biot. 2017, 101, 5033–5043. [Google Scholar] [CrossRef]
- St Leger, R.J.; Joshi, L.; Roberts, D. Ambient pH is a major determinant in the expression of cuticle-degrading enzymes and hydrophobin by Metarhizium anisopliae. Appl. Environ. Microb. 1998, 64, 709–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, Y.; Cao, X.; Chen, J.; Wang, L.; Wei, G.; Wang, S. Coordinated regulation of infection-related morphogenesis by the KMT2-Cre1-Hyd4 regulatory pathway to facilitate fungal infection. Sci. Adv. 2020, 6, eaaz1659. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Jiang, H.; Du, Y.; Keyhani, N.O.; Xia, Y.; Jin, K. Members of chitin synthase family in Metarhizium acridum differentially affect fungal growth, stress tolerances, cell wall integrity and virulence. PLoS Pathog. 2019, 15, e1007964. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Zhou, X.; Xia, Y.; Cao, Y. Mid1 affects ion transport, cell wall integrity, and host penetration of the entomopathogenic fungus Metarhizium acridum. Appl. Microbiol. Biot. 2019, 103, 1801–1810. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wei, Q.; Xia, Y.; Jin, K. MaPacC, a pH-responsive transcription factor, negatively regulates thermotolerance and contributes to conidiation and virulence in Metarhizium acridum. Curr. Genet. 2020, 66, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wang, J.; Yin, Y.; Deng, C.; Yang, K.; Wang, Z. MripacC regulates blastosphere budding and influences virulence of the pathogenic fungus Metarhizium rileyi. Fungal Biol. 2021, 125, 596–608. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; St Leger, R.J. The MAD1 adhesin of Metarhizium anisopliae links adhesion with blastospore production and virulence to insects, and the MAD2 adhesin enables attachment to plants. Eukaryot. Cell 2007, 6, 808–816. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zhang, Q.; Xia, Y.; Jin, K. MaNmrA, a Negative Transcription Regulator in Nitrogen Catabolite Repression Pathway, Contributes to Nutrient Utilization, Stress Resistance, and Virulence in Entomopathogenic Fungus Metarhizium acridum. Biology 2021, 10, 1167. [Google Scholar] [CrossRef]
- Li, C.; Zhang, Q.; Xia, Y.; Jin, K. MaAreB, a GATA Transcription Factor, Is Involved in Nitrogen Source Utilization, Stress Tolerances and Virulence in Metarhizium acridum. J. Fungi. 2021, 7, 512. [Google Scholar] [CrossRef]
- Du, Y.; Jin, K.; Xia, Y. Involvement of MaSom1, a downstream transcriptional factor of cAMP/PKA pathway, in conidial yield, stress tolerances, and virulence in Metarhizium acridum. Appl. Microbiol. Biot. 2018, 102, 5611–5623. [Google Scholar] [CrossRef]
- Jin, K.; Han, L.; Xia, Y. MaMk1, a FUS3/KSS1-type mitogen-activated protein kinase gene, is required for appressorium formation, and insect cuticle penetration of the entomopathogenic fungus Metarhizium acridum. J. Invertebr. Pathol. 2014, 115, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Mei, L.; Wang, X.; Yin, Y.; Tang, G.; Wang, C. Conservative production of galactosaminogalactan in Metarhizium is responsible for appressorium mucilage production and topical infection of insect hosts. PLoS Pathog. 2021, 17, e1009656. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Xie, R.; Wang, Y.; Xie, T.; Xu, L.; Huang, B. The G-protein coupled receptor GPRK contributes to fungal development and full virulence in Metarhizium robertsii. J. Invertebr. Pathol. 2021, 183, 107627. [Google Scholar] [CrossRef] [PubMed]
- Latge, J.P. Tasting the fungal cell wall. Cell. Microbiol. 2010, 12, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Jarrold, S.L.; Moore, D.; Potter, U.; Charnley, A.K. The contribution of surface waxes to pre-penetration growth of an entomopathogenic fungus on host cuticle. Mycol. Res. 2007, 111, 240–249. [Google Scholar] [CrossRef]
- Wang, C.S.; Leger, R.J.S. Developmental and transcriptional responses to host and nonhost cuticles by the specific locust pathogen Metarhizium anisopliae var. acridum. Eukaryot. Cell 2005, 4, 937–947. [Google Scholar] [CrossRef] [Green Version]
- Ment, D.; Gindin, G.; Rot, A.; Eshel, D.; Teper-Bamnolker, P.; Ben-Ze’ev, I.; Glazer, I.; Samish, M. Role of cuticular lipids and water-soluble compounds in tick susceptibility to Metarhizium infection. Biocontrol Sci. Technol. 2013, 23, 956–967. [Google Scholar] [CrossRef]
- Saito, H. Regulation of cross-talk in yeast MAPK signaling pathways. Curr. Opin. Microbiol. 2010, 13, 677–683. [Google Scholar] [CrossRef]
- Wang, Z.; Feng, J.; Jiang, Y.; Xu, X.; Xu, L.; Zhou, Q.; Huang, B. MrPEX33 is involved in infection-related morphogenesis and pathogenicity of Metarhizium robertsii. Appl. Microbiol. Biot. 2021, 105, 1079–1090. [Google Scholar] [CrossRef]
- Chen, X.; Xu, C.; Qian, Y.; Liu, R.; Zhang, Q.; Zeng, G.; Zhang, X.; Zhao, H.; Fang, W. MAPK cascade-mediated regulation of pathogenicity, conidiation and tolerance to abiotic stresses in the entomopathogenic fungus Metarhizium robertsii. Environ. Microbiol. 2016, 18, 1048–1062. [Google Scholar] [CrossRef]
- Guo, N.; Qian, Y.; Zhang, Q.; Chen, X.; Zeng, G.; Zhang, X.; Mi, W.; Xu, C.; Leger, R.J.S.; Fang, W. Alternative transcription start site selection in Mr-OPY2 controls lifestyle transitions in the fungus Metarhizium robertsii. Nat. Commun. 2017, 8, 1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Q.; Du, Y.; Jin, K.; Xia, Y. The Ste12-like transcription factor MaSte12 is involved in pathogenicity by regulating the appressorium formation in the entomopathogenic fungus, Metarhizium acridum. Appl. Microbiol. Biot. 2017, 101, 8571–8584. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Wang, J.; Yang, K.; Fan, L.; Wang, Z.; Yin, Y. Regulation of conidiation, polarity growth, and pathogenicity by MrSte12 transcription factor in entomopathogenic fungus, Metarhizium rileyi. Fungal Genet. Biol. 2021, 155, 103612. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Zhang, X.; Guo, N.; Fang, W. MrSt12 implicated in the regulation of transcription factor AFTF1 by Fus3-MAPK during cuticle penetration by the entomopathogenic fungus Metarhizium robertsii. Fungal Genet. Biol. 2019, 131, 103244. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Wen, Z.; Xia, Y.; Jin, K. The transmembrane protein MaSho1 negatively regulates conidial yield by shifting the conidiation pattern in Metarhizium acridum. Appl. Microbiol. Biot. 2020, 104, 4005–4015. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Duan, Z.; Leger, R.J.S. MOS1 osmosensor of Metarhizium anisopliae is required for adaptation to insect host hemolymph. Eukaryot. Cell 2008, 7, 302–309. [Google Scholar] [CrossRef] [Green Version]
- Shang, J.; Shang, Y.; Tang, G.; Wang, C. Identificatio.on of a key G-protein coupled receptor in mediating appressorium formation and fungal virulence against insects. Sci. China-Life Sci. 2021, 64, 466–477. [Google Scholar] [CrossRef]
- Tong, Y.; Wu, H.; Liu, Z.; Wang, Z.; Huang, B. G-Protein Subunit Galphai in Mitochondria, MrGPA1, Affects Conidiation, Stress Resistance, and Virulence of Entomopathogenic Fungus Metarhizium robertsii. Front. Microbiol. 2020, 11, 1251. [Google Scholar] [CrossRef]
- Liu, S.; Peng, G.; Xia, Y. The adenylate cyclase gene MaAC is required for virulence and multi-stress tolerance of Metarhizium acridum. BMC Microbiol. 2012, 12, 163. [Google Scholar] [CrossRef] [Green Version]
- St Leger, R.J.; Butt, T.M.; Staples, R.C.; Roberts, D.W. Second messenger involvement in differentiation of the entomopathogenic fungus Metarhizium anisopliae. J. Gen. Microbiol. 1990, 136, 1779–1789. [Google Scholar] [CrossRef] [Green Version]
- Fang, W.; Pava-ripoll, M.; Wang, S.; St Leger, R. Protein kinase A regulates production of virulence determinants by the entomopathogenic fungus, Metarhizium anisopliae. Fungal Genet. Biol. 2009, 46, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Du, M.; Luo, S.; Xia, Y. Calcineurin modulates growth, stress tolerance, and virulence in Metarhizium acridum and its regulatory network. Appl. Microbiol. Biot. 2014, 98, 8253–8265. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; He, M.; Cao, Y.; Xia, Y. The tetraspanin gene MaPls1 contributes to virulence by affecting germination, appressorial function and enzymes for cuticle degradation in the entomopathogenic fungus, Metarhizium acridum. Environ. Microbiol. 2013, 15, 2966–2979. [Google Scholar] [PubMed]
- Zhao, T.; Tian, H.; Xia, Y.; Jin, K. MaPmt4, a protein O-mannosyltransferase, contributes to cell wall integrity, stress tolerance and virulence in Metarhizium acridum. Curr. Genet. 2019, 65, 1025–1040. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Tian, H.; Xia, Y.; Jin, K. O-mannosyltransferase MaPmt2 contributes to stress tolerance, cell wall integrity and virulence in Metarhizium acridum. J Invertebr. Pathol. 2021, 184, 107649. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jin, K.; Xia, Y. Contributions of beta-tubulin to cellular morphology, sporulation and virulence in the insect-fungal pathogen, Metarhizium acridum. Fungal Genet. Biol. 2017, 103, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Ming, Y.; Wei, Q.; Jin, K.; Xia, Y. MaSnf1, a sucrose non-fermenting protein kinase gene, is involved in carbon source utilization, stress tolerance, and virulence in Metarhizium acridum. Appl. Microbiol. Biot. 2014, 98, 10153–10164. [Google Scholar] [CrossRef]
- Li, G.; Fan, A.; Peng, G.; Keyhani, N.O.; Xin, J.; Cao, Y.; Xia, Y. A bifunctional catalase-peroxidase, MakatG1, contributes to virulence of Metarhizium acridum by overcoming oxidative stress on the host insect cuticle. Environ. Microbiol. 2017, 19, 4365–4378. [Google Scholar] [CrossRef]
- Tang, G.; Shang, Y.; Li, S.; Wang, C. MrHex1 is Required for Woronin Body Formation, Fungal Development and Virulence in Metarhizium robertsii. J. Fungi. 2020, 6, 172. [Google Scholar] [CrossRef]
- Zeng, G.; Zhang, P.; Zhang, Q.; Zhao, H.; Li, Z.; Zhang, X.; Wang, C.; Yin, W.-B.; Fang, W. Duplication of a Pks gene cluster and subsequent functional diversification facilitate environmental adaptation in Metarhizium species. PLoS Genet. 2018, 14, e1007472. [Google Scholar] [CrossRef]
- Shang, Y.; Chen, P.; Chen, Y.; Lu, Y.; Wang, C. MrSkn7 Controls Sporulation, Cell Wall Integrity, Autolysis, and Virulence in Metarhizium robertsii. Eukaryot. Cell 2015, 14, 396–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enrique Padilla-Guerrero, I.; Barelli, L.; Angelica Gonzalez-Hernandez, G.; Carlos Torres-Guzman, J.; Bidochka, M.J. Flexible metabolism in Metarhizium anisopliae and Beauveria bassiana: Role of the glyoxylate cycle during insect pathogenesis. Microbiology 2011, 157, 199–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Wang, H.; Keyhani, N.O.; Xia, Y.; Peng, G.G. Disruption of an adenylate-forming reductase required for conidiation, increases virulence of the insect pathogenic fungus Metarhizium acridum by enhancing cuticle invasion. Pest. Manag. Sci. 2020, 76, 758–768. [Google Scholar] [CrossRef]
- St. Leger, R.J.; Roberts, D.W.; Staples, R.C. A model to explain differentiation of appressoria by germlings of Metarhizium anisopliae. J. Invertebr. Pathol. 1991, 57, 299–310. [Google Scholar] [CrossRef]
- Abu Sadat, M.; Jeon, J.; Mir, A.A.; Choi, J.; Choi, J.; Lee, Y.-H. Regulation of Cellular Diacylglycerol through Lipid Phosphate Phosphatases Is Required for Pathogenesis of the Rice Blast Fungus, Magnaporthe oryzae. PLoS ONE 2014, 9, e100726. [Google Scholar]
- Foster, A.J.; Ryder, L.S.; Kershaw, M.J.; Talbot, N.J. The role of glycerol in the pathogenic lifestyle of the rice blast fungus Magnaporthe oryzae. Environ. Microbiol. 2017, 19, 1008–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, R.A.; Talbot, N.J. Under pressure: Investigating the biology of plant infection by Magnaporthe oryzae. Nat. Rev. Microbiol. 2009, 7, 185–195. [Google Scholar] [CrossRef]
- Wang, C.; Leger, R.J.S. The Metarhizium anisopliae perilipin homolog MPL1 regulates lipid metabolism, appressorial turgor pressure, and virulence. J. Biol. Chem. 2007, 282, 21110–21115. [Google Scholar] [CrossRef] [Green Version]
- Keyhani, N.O. Lipid biology in fungal stress and virulence: Entomopathogenic fungi. Fungal Biol. 2018, 122, 420–429. [Google Scholar] [CrossRef]
- Gao, Q.; Shang, Y.; Huang, W.; Wang, C. Glycerol-3-Phosphate Acyltransferase Contributes to Triacylglycerol Biosynthesis, Lipid Droplet Formation, and Host Invasion in Metarhizium robertsii. Appl. Environ. Microb. 2013, 79, 7646–7653. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Jiang, Y.; Li, Y.; Feng, J.; Huang, B. MrArk1, an actin-regulating kinase gene, is required for endocytosis and involved in sustaining conidiation capacity and virulence in Metarhizium robertsii. Appl. Microbiol. Biot. 2019, 103, 4859–4868. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Tian, H.; Xia, Y.; Jin, K. MaPmt1, a protein O-mannosyltransferase, contributes to virulence through governing the appressorium turgor pressure in Metarhizium acridum. Fungal Genet. Biol. 2020, 145, 103480. [Google Scholar] [CrossRef] [PubMed]
- Akhouayri, I.; Turc, C.; Royet, J.; Charroux, B. Toll-8/Tollo negatively regulates antimicrobial response in the Drosophila respiratory epithelium. PLoS Pathog. 2011, 7, e1002319. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.; Chen, Y.; Huang, W.; Shang, Y.; Chen, P.; Wang, C. Linkage of autophagy to fungal development, lipid storage and virulence in Metarhizium robertsii. Autophagy 2013, 9, 538–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Hong, S.; Tang, G.; Lu, Y.; Wang, C. Unveiling the function and regulation control of the DUF3129 family proteins in fungal infection of hosts. Philos. Trans. R. Soc. B 2019, 374, 20180321. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Jin, K.; Ying, S.-H.; Zhang, Y.; Xiao, G.; Shang, Y.; Duan, Z.; Hu, X.; Xie, X.-Q.; Zhou, G.; et al. Genome Sequencing and Comparative Transcriptomics of the Model Entomopathogenic Fungi Metarhizium anisopliae and M. acridum. PLoS Genet. 2011, 7, e1001264. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, J.P.; Bateman, R.; Charnley, A.K. Role of cuticle-degrading proteases in the virulence of Metarhizium spp. for the desert locust, Schistocerca gregaria. J. Invertebr. Pathol. 1998, 71, 128–137. [Google Scholar] [CrossRef]
- Fan, Y.; Fang, W.; Guo, S.; Pei, X.; Zhang, Y.; Xiao, Y.; Li, D.; Jin, K.; Bidochka, M.J.; Pei, Y. Increased insect virulence in Beauveria bassiana strains overexpressing an engineered chitinase. Appl. Environ. Microb. 2007, 73, 295–302. [Google Scholar] [CrossRef] [Green Version]
- Leger, R.J.S.; Joshi, L.; Bidochka, M.J.; Rizzo, N.W.; Roberts, D.W. Characterization and ultrastructural localization of chitinases from Metarhizium anisopliae, M. flavoviride, and Beauveria bassiana during fungal invasion of host (Manduca sexta) cuticle. Appl. Environ. Microb. 1996, 62, 907–912. [Google Scholar]
- Hackman, R.H. Structure and function in tick cuticle. Ann. Rev. Entomol. 1982, 27, 75–95. [Google Scholar] [CrossRef]
- Malek, S.R.A. Polyphenols and their quinone derivatives in the cuticle of the desert locust, Schistocerca gregaria (Forskål). Comp. Biochem. Phys. 1961, 2, 35–50. [Google Scholar] [CrossRef]
- Soderhall, K.; Hall, L.; Unestam, T.; Nyhlen, L. Attachment of phenoloxidase to fungal cell walls in arthropod immunity. J. Invertebr. Pathol. 1979, 34, 285–294. [Google Scholar] [CrossRef]
- Gonzalez-Santoyo, I.; Cordoba-Aguilar, A. Phenoloxidase: A key component of the insect immune system. Entomol. Exp. Appl. 2012, 142, 1–16. [Google Scholar] [CrossRef]
- Andersen, S.O. Characterization of a trypsin-solubilized phenoloxidase from locust cuticle. Insect. Biochem. 1978, 8, 6. [Google Scholar] [CrossRef]
- Li, B.; Li, H.; Tian, Y.; Abro, N.A.; Nong, X.; Zhang, Z.; Wang, G. Molecular Identification and Immunity Functional Characterization of Lmserpin1 in Locusta migratoria manilensis. Insects 2021, 12, 178. [Google Scholar] [CrossRef]
- Zhang, W.; Zheng, X.; Chen, J.; Keyhani, N.O.; Cai, K.; Xia, Y. Spatial and temporal transcriptomic analyses reveal locust initiation of immune responses to Metarhizium acridum at the pre-penetration stage. Dev. Comp. Immunol. 2020, 104, 103524. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Li, S.; Si, Y.; Hu, J.; Xia, Y. Locust can detect beta-1, 3-glucan of the fungal pathogen before penetration and defend infection via the Toll signaling pathway. Dev. Comp. Immunol. 2020, 106, 103636. [Google Scholar] [CrossRef]
- Jiang, Z.-Y.; Ligoxygakis, P.; Xia, Y.-X. HYD3, a conidial hydrophobin of the fungal entomopathogen Metarhizium acridum induces the immunity of its specialist host locust. Int. J. Biol. Macromol. 2020, 165, 1303–1311. [Google Scholar] [CrossRef]
- Yu, Y.; Cao, Y.; Xia, Y.; Liu, F. Wright-Giemsa staining to observe phagocytes in Locusta migratoria infected with Metarhizium acridum. J. Invertebr. Pathol. 2016, 139, 19–24. [Google Scholar] [CrossRef]
- Gunnarsson, S.G.S. Infection of Schistocerca gregaria by the fungus Metarhizium anisopliae cellular reactions in the integument studied by scanning electron and light microscopy. J. Invertebr. Pathol. 1988, 52, 9–17. [Google Scholar] [CrossRef]
- Brey, P.T.; Lee, W.J.; Yamakawa, M.; Koizumi, Y.; Perrot, S.; Francois, M.; Ashida, M. Role of the integument in insect immunity—Epicuticular abrasion and induction of cecropin synthesis in cuticular epithelial-cells. Proc. Natl. Acad. Sci. USA 1993, 90, 6275–6279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meister, M.; Braun, A.; Kappler, C.; Reichhart, J.M.; Hoffmann, J.A. Insect immunity—A trransgenic analysis in drosophila defines several functional domains in the diptericin promoter. Embo J. 1994, 13, 5958–5966. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jiang, H.B. Interaction of beta-1,3-glucan with its recognition protein activates hemolymph proteinase 14, an initiation enzyme of the prophenoloxidase activation system in Manduca sexta. J. Biol. Chem. 2006, 281, 9271–9278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Xu, C.; Lu, H.L.; Chen, X.; St Leger, R.J.; Fang, W. Host-to-pathogen gene transfer facilitated infection of insects by a pathogenic fungus. PLoS Pathog. 2014, 10, e1004009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y.X.; Gao, M.Y.; Clarkson, J.M.; Charnley, A.K. Molecular cloning, characterisation, and expression of a neutral trehalase from the insect pathogenic fungus Metarhizium anisopliae. J. Invertebr. Pathol. 2002, 80, 127–137. [Google Scholar] [CrossRef]
- Jin, K.; Peng, G.; Liu, Y.; Xia, Y. The acid trehalase, ATM1, contributes to the in vivo growth and virulence of the entomopathogenic fungus, Metarhizium acridum. Fungal Genet. Biol. 2015, 77, 61–67. [Google Scholar] [CrossRef]
- Peng, G.; Jin, K.; Liu, Y.; Xia, Y. Enhancing the utilization of host trehalose by fungal trehalase improves the virulence of fungal insecticide. Appl. Microbiol. Biot. 2015, 99, 8611–8618. [Google Scholar] [CrossRef]
- Sohda, M.; Misumi, Y.; Ogata, S.; Sakisaka, S.; Hirose, S.; Ikehara, Y.; Oda, K. Trans.-Golgi protein p230/golgin-245 is involved in phagophore formation. Biochem. Biophys. Res. Commun. 2015, 456, 275–281. [Google Scholar] [CrossRef]
- Li, Z.; Wang, C.; Xia, Y. Isolation of two Locust protein targets of a protein tyrosine phosphatase from Metarhizium anisopliae strain CQMa102. J. Invertebr. Pathol. 2008, 99, 151–155. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, S.; Li, X.; Zhu, P.; Xie, E.; Li, Z. Expression, purification, and characterization of a novel acid phosphatase that displays protein tyrosine phosphatases activity from Metarhizium anisopliae strain CQMa102. Biosci. Biotechnol. Biochem. 2017, 81, 2292–2300. [Google Scholar] [CrossRef] [Green Version]
- Gleeson, P.A.; Anderson, T.J.; Stow, J.L.; Griffiths, G.; Toh, B.H.; Matheson, F. p230 is associated with vesicles budding from the trans-Golgi network. J. Cell Sci. 1996, 109, 2811–2821. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Wang, Y.; Li, J.; Hu, S.; Yang, P.; Kang, L. Transformation of glycerate kinase (GLYK) into Metarhizium acridum increases virulence to locust. Pest. Manag. Sci. 2021, 77, 1465–1475. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Sanchez, E.; Lange, A.B.; Orchard, I. Effects of the mycotoxin destruxin A on Locusta migratoria visceral muscles. Toxicon 2010, 56, 1043–1051. [Google Scholar] [CrossRef] [PubMed]
- Sbaraini, N.; Guedes, R.L.; Andreis, F.C.; Junges, A.; de Morais, G.L.; Vainstein, M.H.; de Vasconcelos, A.T.; Schrank, A. Secondary metabolite gene clusters in the entomopathogen fungus Metarhizium anisopliae: Genome identification and patterns of expression in a cuticle infection model. BMC Genom. 2016, 17, 736. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, Y.; Narwani, T.; Subramanian, S. Genome sequence and comparative analysis of clavicipitaceous insect-pathogenic fungus Aschersonia badia with Metarhizium spp. BMC Genom. 2016, 17, 367. [Google Scholar] [CrossRef] [Green Version]
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Chung, H.; Shin, Y.P.; Kim, I.-W.; Natarajan, S.; Veerappan, K.; Seo, M.; Park, J.; Hwang, J.S. In silico strategic curation, retrieval and prediction of novel antimicrobial peptide from Locusta migratoria transcriptome. J. Asia-Pac. Entomol. 2021, 24, 237–242. [Google Scholar] [CrossRef]
- Lv, M.; Mohamed, A.A.; Zhang, L.; Zhang, P.; Zhang, L. A Family of CSalphabeta Defensins and Defensin-Like Peptides from the Migratory Locust, Locusta migratoria, and Their Expression Dynamics during Mycosis and Nosemosis. PLoS ONE 2016, 11, e0161585. [Google Scholar] [CrossRef]
- Zheng, X.; Xia, Y. beta-1,3-Glucan recognition protein (beta GRP) is essential for resistance against fungal pathogen and opportunistic pathogenic gut bacteria in Locusta migratoria manilensis. Dev. Comp. Immunol. 2012, 36, 602–609. [Google Scholar] [CrossRef]
- Han, P.; Han, J.; Zhang, M.; Fan, J.; Gong, Q.; Ma, E.; Zhang, J. 20-Hydroxyecdysone enhances Immulectin-1 mediated immune response against entomogenous fungus in Locusta migratoria. Pest. Manag. Sci. 2020, 76, 304–313. [Google Scholar] [CrossRef]
- Han, P.; Han, J.; Fan, J.; Zhang, M.; Ma, E.; Li, S.; Fan, R.; Zhang, J. 20-Hydroxyecdysone activates PGRP-SA mediated immune response in Locusta migratoria. Dev. Comp. Immunol. 2017, 72, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.F. G protein-coupled receptors function as cell membrane receptors for the steroid hormone 20-hydroxyecdysone. Cell Commun. Signal. 2020, 18, 146. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Keyhani, N.O.; Zhang, H.; Cai, K.; Xia, Y. Inhibitor of apoptosis-1 gene as a potential target for pest control and its involvement in immune regulation during fungal infection. Pest. Manag. Sci. 2020, 76, 1831–1840. [Google Scholar] [CrossRef] [PubMed]
- Cerenius, L.; Lee, B.L.; Soderhall, K. The proPO-system: Pros and cons for its role in invertebrate immunity. Trends Immunol. 2008, 29, 263–271. [Google Scholar] [CrossRef]
- Goldsworthy, G.; Mullen, L.; Opoku-Ware, K.; Chandrakant, S. Interactions between the endocrine and immune systems in locusts. Physiol. Entomol. 2003, 28, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Goldsworthy, G.; Opoku-Ware, K.; Mullen, L. Adipokinetic hormone enhances laminarin and bacterial lipopolysaccharide-induced activation of the prophenoloxidase cascade in the African migratory locust, Locusta migratoria. J. Insect Physiol. 2002, 48, 601–608. [Google Scholar] [CrossRef] [Green Version]
- Goldsworthy, G.J.; Opoku-Ware, K.; Mullen, L.M. Adipokinetic hormone and the immune responses of locusts to infection. Ann. N. Y. Acad. Sci. 2005, 1040, 106–113. [Google Scholar] [CrossRef]
- Abdelatef, G.M.; El-Maghraby, M.M.A.; Gomma, E.A.; Metaweh, H.H. Haemolymph Picture and Its Chemical Components in Schistocerca gregaria (Forskal) as Affected by Two Entomopathogenic Fungi. Egypt. J. Biol. Pest Control 2009, 19, 119–128. [Google Scholar]
- Han, P.; Gong, Q.; Fan, J.; Zhang, M.; Abbas, M.; Zhu, W.; Deng, S.; Xing, S.; Zhang, J. 20-Hydroxyecdysone regulates the prophenoloxidase cascade to immunize Metarhizium anisopliae in Locusta migratoria. Pest Manag. Sci. 2020, 76, 3149–3158. [Google Scholar] [CrossRef]
- Gillespie, J.P.; Burnett, C.; Charnley, A.K. The immune response of the desert locust Schistocerca gregaria during mycosis of the entomopathogenic fungus, Metarhizium anisopliae var acridum. J. Insect Physiol. 2000, 46, 429–437. [Google Scholar] [CrossRef]
- Smith, R.C.; Barillas-Mury, C.; Jacobs-Lorena, M. Hemocyte differentiation mediates the mosquito late-phase immune response against Plasmodium in Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2015, 112, e3412–e3420. [Google Scholar] [CrossRef] [Green Version]
- Eleftherianos, I.; Heryanto, C.; Bassal, T.; Zhang, W.; Tettamanti, G.; Mohamed, A. Haemocyte-mediated immunity in insects: Cells, processes and associated components in the fight against pathogens and parasites. Immunology 2021, 164, 401–432. [Google Scholar] [CrossRef] [PubMed]
- Boltana, S.; Rey, S.; Roher, N.; Vargas, R.; Huerta, M.; Huntingford, F.A.; Goetz, F.W.; Moore, J.; Garcia-Valtanen, P.; Estepa, A.; et al. Behavioural fever is a synergic signal amplifying the innate immune response. Proc. Biol. Sci. 2013, 280, 20131381. [Google Scholar] [CrossRef] [PubMed]
- Hunt, V.L.; Lock, G.D.; Pickering, S.G.; Charnley, A.K. Application of infrared thermography to the study of behavioural fever in the desert locust. J. Therm. Biol. 2011, 36, 443–451. [Google Scholar] [CrossRef]
- Bundey, S.; Raymond, S.; Dean, P.; Roberts, S.K.; Dillon, R.J.; Charnley, A.K. Eicosanoid involvement in the regulation of behavioral fever in the desert locust, Schistocerca gregaria. Arch. Insect Biochem. Physiol. 2003, 52, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.C.; Kim, Y. Toll signal pathway activating eicosanoid biosynthesis shares its conserved upstream recognition components in a lepidopteran Spodoptera exigua upon infection by Metarhizium rileyi, an entomopathogenic fungus. J. Invertebr. Pathol. 2022, 188, 107707. [Google Scholar] [CrossRef]
- Hunt, V.L.; Charnley, A.K. The inhibitory effect of the fungal toxin, destruxin A, on behavioural fever in the desert locust. J. Insect Physiol. 2011, 57, 1341–1346. [Google Scholar] [CrossRef]
- Ouedraogo, R.M.; Goettel, M.S.; Brodeur, J. Behavioral thermoregulation in the migratory locust: A therapy to overcome fungal infection. Oecologia 2004, 138, 312–319. [Google Scholar] [CrossRef]
- Springate, S.; Thomas, M.B. Thermal biology of the meadow grasshopper, Chorthippus parallelus, and the implications for resistance to disease. Ecol. Entomol. 2005, 30, 724–732. [Google Scholar] [CrossRef]
- Wilson, K.; Reeson, A.F. Density-dependent prophylaxis: Evidence from Lepidoptera–baculovirus interactions? Ecol. Entomol. 1998, 23, 100–101. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.A.; Pell, J.K.; Simpson, S.J. Crowded locusts produce hatchlings vulnerable to fungal attack. Biol. Lett. 2009, 5, 845–848. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, P.; Cui, F.; Kang, L. Altered immunity in crowded locust reduced fungal (Metarhizium anisopliae) pathogenesis. PLoS Pathog. 2013, 9, e1003102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Tong, X.; Yuan, S.; Yang, P.; Li, L.; Zhao, Y.; Kang, L. Variation of TNF modulates cellular immunity of gregarious and solitary locusts against fungal pathogen Metarhizium anisopliae. Proc. Natl. Acad. Sci. USA 2022, 119, e2120835119. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Xia, Y. Host–Pathogen Interactions between Metarhizium spp. and Locusts. J. Fungi 2022, 8, 602. https://doi.org/10.3390/jof8060602
Li J, Xia Y. Host–Pathogen Interactions between Metarhizium spp. and Locusts. Journal of Fungi. 2022; 8(6):602. https://doi.org/10.3390/jof8060602
Chicago/Turabian StyleLi, Jun, and Yuxian Xia. 2022. "Host–Pathogen Interactions between Metarhizium spp. and Locusts" Journal of Fungi 8, no. 6: 602. https://doi.org/10.3390/jof8060602
APA StyleLi, J., & Xia, Y. (2022). Host–Pathogen Interactions between Metarhizium spp. and Locusts. Journal of Fungi, 8(6), 602. https://doi.org/10.3390/jof8060602