
Variability in Nutrient Use by Orchid Mycorrhizal Fungi in Two Medium Types



Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolate Preparation
2.2. Phylogenetic Analyses
2.3. Experimental Media
2.3.1. Carbon Sources
2.3.2. Nitrogen Sources
2.3.3. Phosphorus Sources
2.3.4. Solid Cultures
2.3.5. Liquid Cultures
Determination of Exponential Growth Phase of the Isolates
Liquid Culture Experiment
2.4. Statistical Analysis
3. Results
3.1. Fungal Biomass Production in Solid and Liquid Media
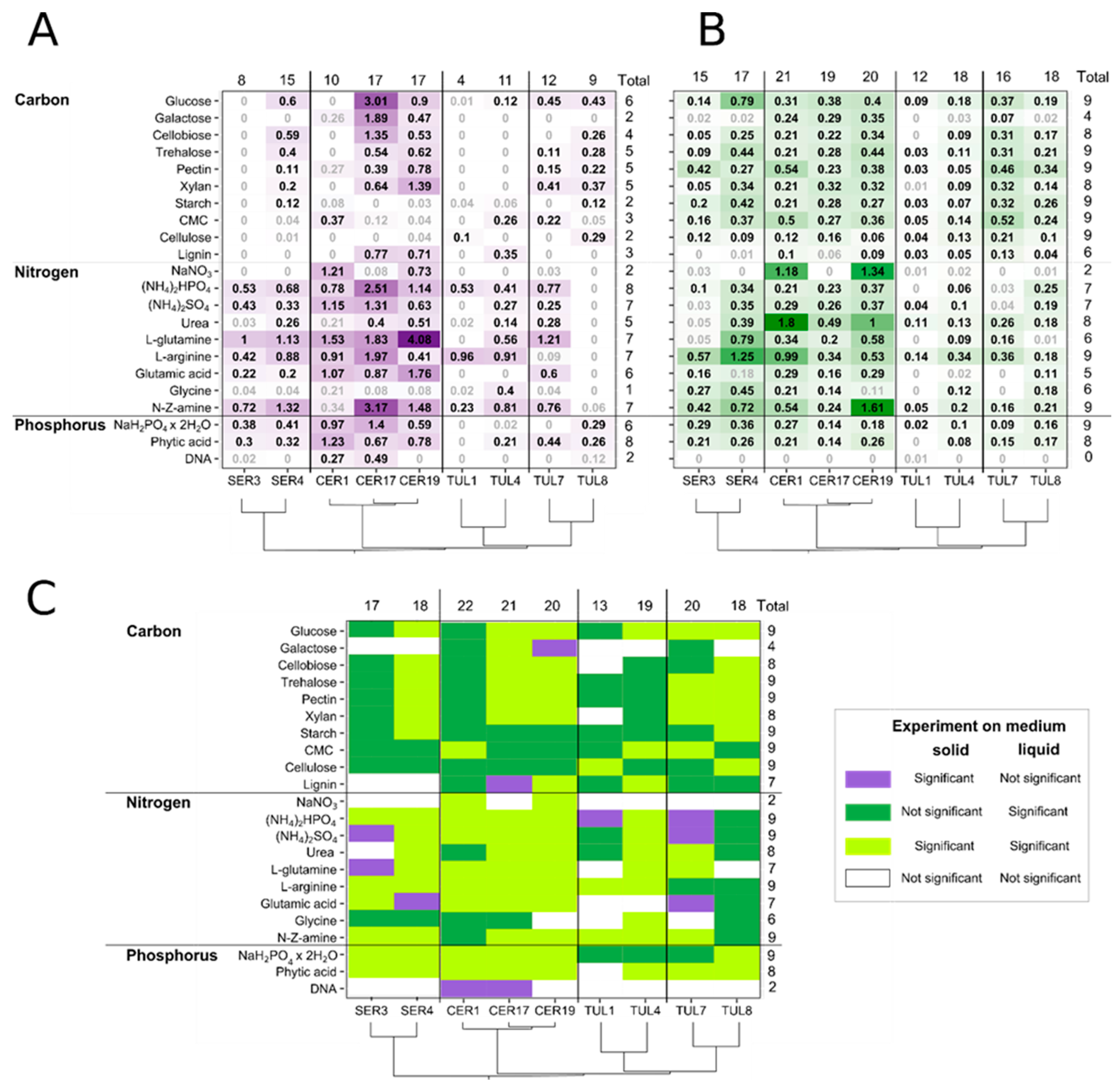
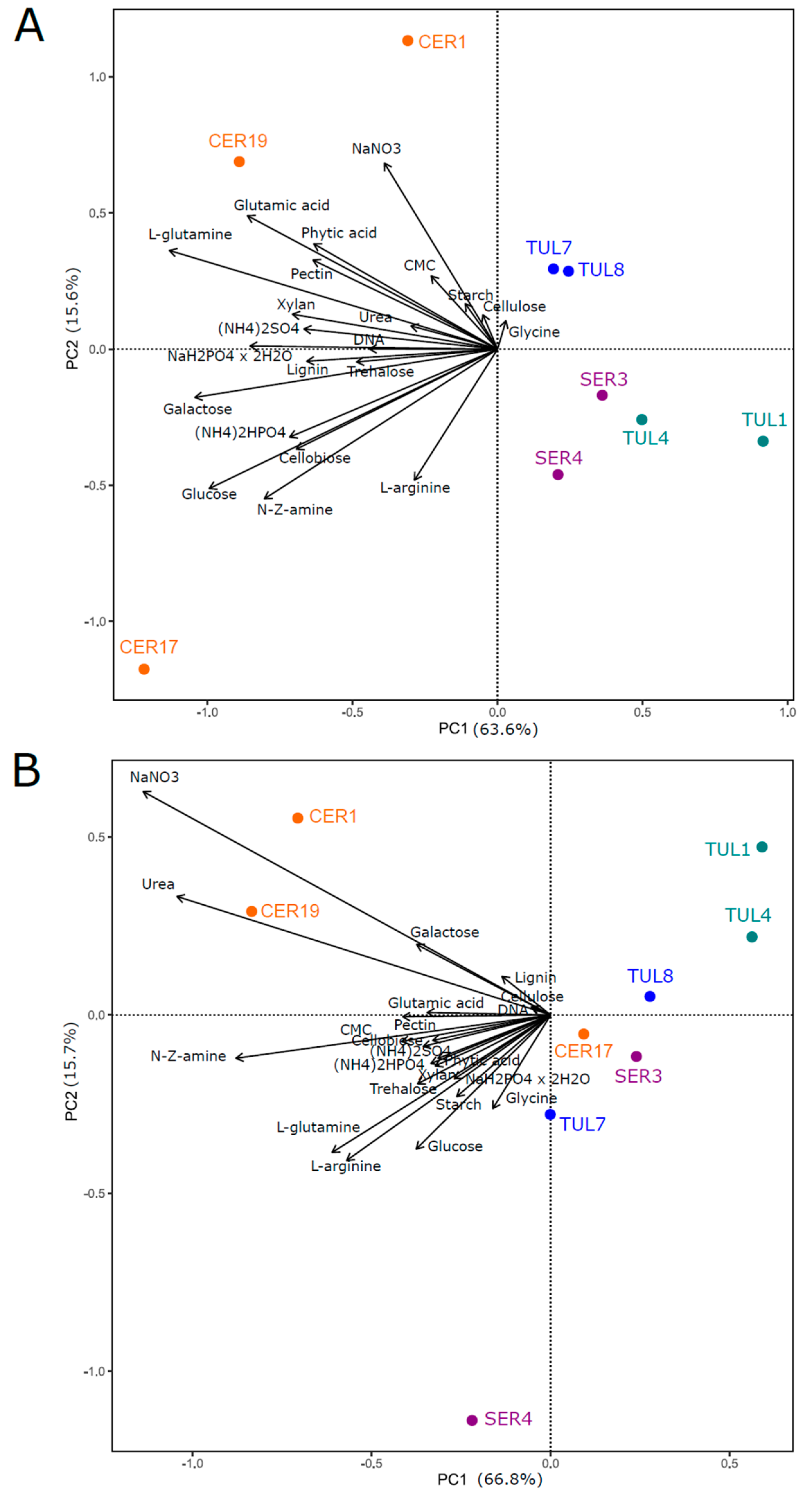
3.2. Carbon Sources
3.3. Nitrogen Sources
3.4. Phosphorus Sources
3.5. Comparison of Radial Growth Rates on Solid Medium
4. Discussion
4.1. Carbon Utilization
4.2. Nitrogen Utilization
4.3. Phosphorus Utilization
4.4. Disputable Use of Radial Growth Rate
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Kohler, A.; Kuo, A.; Nagy, L.G.; Morin, E.; Barry, K.W.; Buscot, F.; Canbäck, B.; Choi, C.; Cichocki, N.; Clum, A.; et al. Convergent losses of decay mechanisms and rapid turnover of symbiosis genes in mycorrhizal mutualists. Nat. Genet. 2015, 47, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, H.N. Terrestrial Orchids. From Seed to Mycotrophic Plant, 1st ed.; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Weiss, M.; Waller, F.; Zuccaro, A.; Selosse, M.A. Sebacinales—One thousand and one interactions with land plants. New Phytol. 2016, 211, 20–40. [Google Scholar] [CrossRef]
- Selosse, M.A.; Martos, F. Do chlorophyllous orchids heterotrophically use mycorrhizal fungal carbon? Trends Plant Sci. 2014, 19, 683–685. [Google Scholar] [CrossRef] [PubMed]
- Selosse, M.A.; Dubois, M.P.; Alvarez, N. Do Sebacinales commonly associate with plant roots as endophytes? Mycol. Res. 2009, 113, 1062–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veldre, V.; Abarenkov, K.; Bahram, M.; Martos, F.; Selosse, M.A.; Tamm, H.; Köljalg, U.; Tedersoo, L. Evolution of nutritional modes of Ceratobasidiaceae (Cantharellales, Basidiomycota) as revealed from publicly available ITS sequences. Fungal Ecol. 2013, 6, 256–268. [Google Scholar] [CrossRef]
- Bidartondo, M.I.; Bruns, T.D.; Weiß, M.; Sergio, S.; Read, D.J. Specialized cheating of the ectomycorrhizal symbiosis by an epiparasitic liverwort. Proc. R. Soc. Lond. B 2003, 270, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Ceresini, P.C.; Costa-Souza, E.; Zala, M.; Furtado, E.L.; Souza, N.L. Evidence that the Ceratobasidium-like white-thread blight and black rot fungal pathogens from persimmon and tea crops in the Brazilian Atlantic Forest agroecosystem are two distinct phylospecies. Genet. Mol. Biol. 2012, 35, 480–497. [Google Scholar] [CrossRef] [Green Version]
- Mosquera-Espinosa, A.T.; Bayman, P.; Prado, G.A.; Gómez-Carabalí, A.; Otero, J.T. The double life of Ceratobasidium: Orchid mycorrhizal fungi and their potential for biocontrol of Rhizoctonia solani sheath blight of rice. Mycologia 2012, 105, 141–150. [Google Scholar] [CrossRef]
- Adamo, M.; Chialva, M.; Calevo, J.; De Rose, S.; Girlanda, M.; Perotto, S.; Balestrini, R. The dark site of orchid symbiosis: Can Tulasnella calospora decompose host tissues? Int. J. Mol. Sci. 2020, 21, 3139. [Google Scholar] [CrossRef]
- Suetsugu, K.; Matsubayashi, J.; Tayasu, I. Some mycoheterotrophic orchids depends on carbon from dead wood: Novel evidence from a radiocarbon approach. New Phytol. 2020, 227, 1519–1529. [Google Scholar] [CrossRef]
- van der Kinderen, G. A method for the study of field germinated seeds of terrestrial orchids. Lindleyana 1995, 10, 68–73. [Google Scholar]
- Boddy, L.; Frankland, J.C.; van West, P. Ecology of Saptrotrophic Basidiomycetes, 1st ed.; Elsevier Science: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Baldrian, P. Enzymes of saprotrophic basidiomycetes. In Ecology of Satprotrophic Basidiomycetes, 1st ed.; Boody, L., Frankland, J.C., van West, P., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2008; Chapter 2; pp. 19–42. [Google Scholar]
- Berg, B.; Laskowski, R. Litter Decomposition: A Guide to Carbon and Nutrient Turnover; Elsevier Science Publishing Co Inc.: San Diego, CA, USA, 2006. [Google Scholar]
- Kallenbach, C.; Frey, S.; Grandy, A. Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls. Nat. Commun. 2016, 7, 13630. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, F.J. Organic forms of soil nitrogen. In Nitrogen in Agricultural Soils; Stevenson, F.J., Ed.; American Society of Agronomy: Madison, WI, USA, 1982; Volume 22, pp. 67–122. [Google Scholar]
- Richardson, A.E.; Simpson, R. Soil microorganisms mediating phosphorus availability update on microbial phosphorus. Plant Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muindi, E.M. Understanding soil phosphorus. Int. J. Plant Soil Sci. 2019, 31, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Midgley, D.J.; Jordan, L.A.; Saleeba, J.A.; McGee, P.A. Utilisation of carbon substrates by orchid and ericoid mycorrhizal fungi from Australian dry sclerophyll forests. Mycorrhiza 2006, 16, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Nurfadilah, S.; Swarts, N.D.; Dixon, K.W.; Lambers, H.; Merritt, D.J. Variation in nutrient-acquisition patterns by mycorrhizal fungi of rare and common orchids explains diversification in a global biodiversity hotspot. Ann. Bot. 2013, 111, 1233–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehra, S.; Morrison, P.D.; Coates, F.; Lawrie, A.C. Differences in carbon source utilisation by orchid mycorrhizal fungi from common and endangered species of Caladenia (Orchidaceae). Mycorrhiza 2017, 27, 95–108. [Google Scholar] [CrossRef]
- Oktalira, F.T. Diversity of Serendipitaceae Mycorrhizal Associations of Australian Terrestrial Orchids. Ph.D. Thesis, The Australian National University, Canberra, Australia, 2021. [Google Scholar]
- Fochi, V.; Chitarra, V.; Kohler, A.; Voyron, S.; Singan, V.R.; Lindquist, E.A.; Barry, K.W.; Girlanda, M.; Grigoriev, I.V.; Martin, F.; et al. Fungal and plant gene expression in the Tulasnella calospora–Serapias vomeracea symbiosis provides clues about nitrogen pathways in orchid mycorrhizas. New Phytol. 2017, 213, 365–379. [Google Scholar] [CrossRef] [Green Version]
- Freestone, M.; Linde, C.; Swarts, N.; Reiter, N. Ceratobasidium orchid mycorrhizal fungi reveal intraspecific variation and interaction with different nutrient media in symbiotic germination of Prasophyllum (Orchidaceae). Symbiosis 2022, 87, 255–268. [Google Scholar] [CrossRef]
- Cruz, D.; Suárez, J.P.; Kottke, I.; Piepenbring, M.; Oberwinkler, F. Defining species in Tulasnella by correlating morphology and nrDNA ITS-5.8S sequence data of basidiomata from a tropical Andean forest. Mycol. Prog. 2011, 10, 229–238. [Google Scholar] [CrossRef]
- Oberwinkler, F.; Riess, K.; Bauer, R.; Kirschner, R.; Garnica, S. Taxonomic re-evaluation of the Ceratobasidium-Rhizoctonia complex and Rhizoctonia butinii, a new species attacking spruce. Mycol. Prog. 2013, 12, 763–776. [Google Scholar] [CrossRef]
- González, D.; Rodriguez-Carres, M.; Boekhout, T.; Stalpers, J.; Kuramae, E.E.; Nakatani, A.K.; Vilgalys, R.; Cubeta, M.A. Phylogenetic relationships of Rhizoctonia fungi within the Cantharellales. Fungal Biol. 2016, 120, 603–619. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, P.A.; Seviour, R.J.; Schmid, F. Growth of filamentous fungi in submerged culture: Problems and possible solutions. Crit. Rev. Biotechnol. 2000, 20, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Pradeep, F.S.; Begam, M.S.; Palaniswamy, M.; Pradeep, B.V. Influence of culture media on growth and pigment production by Fusarium moniliforme KUMBF1201 isolated from Paddy Field Soil. World Appl. Sci. J. 2013, 22, 70–77. [Google Scholar]
- Viniegra-González, G.; Favela-Torres, E.; Aguilar, C.N.; Rómero-Gomez, S.J.; Díaz-Godínez, G.; Augur, C. Advantages of fungal enzyme production in solid state over liquid fermentation systems. Biochem. Eng. J. 2003, 13, 157–167. [Google Scholar] [CrossRef]
- Marx, D.H.; Bryan, W.C. Growth and ectomycorrhizal development of loblolly pine seedlings in fumigated soil infested with the fungal symbiont Pisolithus tinctorius. For. Sci. 1975, 21, 245–254. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Taylor, D.L.; McCormick, M.K. Internal transcribed spacer primers and sequences for improved characterization of basidiomycetous orchid mycorrhizas. New Phytol. 2008, 177, 1020–1033. [Google Scholar] [CrossRef]
- Těšitelová, T.; Jersáková, J.; Roy, M.; Kubátová, B.; Těšitel, J.; Urfus, T.; Trávníček, P.; Suda, J. Ploidy-specific symbiotic interactions: Divergence of mycorrhizal fungi between cytotypes of the Gymnadenia conopsea group (Orchidaceae). New Phytol. 2013, 199, 1022–1033. [Google Scholar] [CrossRef]
- Milne, I.; Wright, F.; Rowe, G.; Marshall, D.F.; Husmeier, D.; McGuire, G. TOPALi: Software for automatic identification of recombinant sequences within DNA multiple alignments. Bioinformatics 2004, 20, 1806–1807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 1 February 2022).
- O’Hara, R.B.; Simpson, G.L.; Solymos, P.M.; Stevens, H.H.; Szoecs, E.; Wagne, H. Package “Vegan”: Community Ecology Package. R Package Version 2.5-1. 2018. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 February 2022).
- Grothendieck, G. LME4: Linear Mixed-Effects Models using ‘Eigen’ and S4. R Package Version 1.1-10. 2015. Available online: http://lme4.r-forge.r-project.org (accessed on 1 February 2022).
- Mehra, S. Nutritional and Genetic Diversity in Orchid Mycorrhizal Fungi from Caladenia Species. PhD Thesis, RMIT University, Victoria, Australia, December 2014. [Google Scholar]
- Gessner, M.O. Ergosterol as a measure of fungal biomass. In Methods to Study Litter Decomposition, 2nd ed.; Graça, M.A., Bärlocher, F., Gessner, M.O., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 189–195. [Google Scholar]
- Wright, M.; Cross, R.; Cousens, R.D.; May, T.W.; McLean, C.B. The functional significance for the orchid Caladenia tentaculata of genetic and geographic variation in the mycorrhizal fungus Sebacina vermifera s. l. complex. Muelleria 2011, 29, 130–140. [Google Scholar] [CrossRef]
- Dearnaley, J.D.W.; Perotto, S.; Selosse, M.A. Structure and development of orchid mycorrhizas. In Molecular Mycorrhizal Symbiosis, 1st ed.; Martin, F., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 63–86. [Google Scholar]
- Freestone, M.W.; Swarts, N.D.; Reiter, N.; Tomlinson, S.; Sussmilch, F.C.; Wright, M.M.; Holmes, G.D.; Phillips, R.D.; Linde, C.C. Continental-scale distribution and diversity of Ceratobasidium orchid mycorrhizal fungi in Australia. Ann. Bot. 2021, 128, 329–343. [Google Scholar] [CrossRef]
- Di Lonardo, D.P.; Van der Wal, A.; Harkes, P.; de Boer, W. Effect of nitrogen on fungal growth efficiency. Plant Biosyst. 2020, 154, 433–437. [Google Scholar] [CrossRef]
- Meti, R.S.; Ambarish, S.; Khajure, P.V. Enzymes of ammonia assimilation in fungi: An overview. Recent Res. Sci. Technol. 2011, 2, 28–38. [Google Scholar]
- Schweiger, J.M.I.; Bidartondo, M.I.; Gebauer, G. Stable isotope signatures of underground seedlings reveal the organic matter gained by adult orchids from mycorrhizal fungi. Fungal Ecol. 2017, 32, 870–881. [Google Scholar] [CrossRef]
- Cameron, D.D.; Leake, J.R.; Read, D.J. Mutualistic mycorrhiza in orchids: Evidence from plant-fungus carbon and nitrogen transfers in the green-leaved terrestrial orchid Goodyera repens. New Phytol. 2006, 171, 405–416. [Google Scholar] [CrossRef]
- Ray, P.; Abraham, P.E.; Guo, Y.; Giannone, R.J.; Engle, N.L.; Yang, Z.K.; Jacobson, D.; Hettich, R.L.; Tschaplinski, T.J.; Craven, K.D. Scavenging organic nitrogen and remodelling lipid metabolism are key survival strategies adopted by the endophytic fungi, Serendipita vermifera and Serendipita bescii to alleviate nitrogen and phosphorous starvation in vitro. Environ. Microbiol. Rep. 2019, 11, 548–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadley, G.; Ong, S.H. Nutritional requirements of orchid endophytes. New Phytol. 1978, 81, 561–569. [Google Scholar] [CrossRef]
- Tudzynski, B. Nitrogen regulation of fungal secondary metabolism in fungi. Front. Microbiol. 2014, 5, 656. [Google Scholar] [CrossRef] [Green Version]
- Dijk, E.; Eck, N. Effects of mycorrhizal fungi on in vitro nitrogen response of some Dutch indigenous orchid species. Canad. J. Bot. 1995, 73, 1203–1211. [Google Scholar] [CrossRef]
- Vogt-Schilb, H.; Těšitelová, T.; Kotilínek, M.; Sucháček, P.; Kohout, P.; Jersáková, J. Altered rhizoctonia assemblages in grasslands on ex-arable land support germination of mycorrhizal generalist, not specialist orchids. New Phytol. 2020, 227, 1200–1212. [Google Scholar] [CrossRef]
- Figura, T.; Tylová, E.; Jersáková, J.; Vohník, M.; Ponert, J. Fungal symbionts may modulate nitrate inhibitory effect on orchid seed germination. Mycorrhiza 2021, 31, 231–241. [Google Scholar] [CrossRef]
- Zuccaro, A.; Lahrmann, U.; Güldener, U.; Langen, G.; Pfiffi, S.; Biedenkopf, D.; Wong, P.; Samans, B.; Grimm, C.; Basiewicz, M.; et al. Endophytic life strategies decoded by genome and transcriptome analyses of the mutualistic root symbiont Piriformospora indica. PLoS Pathog. 2011, 7, e1002290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephen, R.C.; Fung, K.K. Nitrogen requirements of the fungal endophytes of Arundina chinensis. Canad. J. Bot. 1971, 49, 407–410. [Google Scholar] [CrossRef]
- Hobbie, E.A.; Sánchez, F.S.; Rygiewicz, P.T. Carbon use, nitrogen use, and isotopic fractionation of ectomycorrhizal and saprotrophic fungi in natural abundance and 13C-labelled cultures. Mycol. Res. 2004, 108, 725–736. [Google Scholar] [CrossRef]
- Johri, A.K.; Oelmüller, R.; Dua, M.; Yadav, V.; Kumar, M.; Tuteja, N.; Varma, A.; Bonfante, P.; Persoon, B.L.; Stroud, R.M. Fungal association and utilization of phosphate by plants: Success, limitations, and future prospects. Front. Microbiol. 2015, 6, 984. [Google Scholar] [CrossRef] [Green Version]
- Jansa, J.; Finlay, R.; Wallander, H.; Smith, A.; Smith, S.E. Role of Mycorrhizal Symbioses in Phosphorus Cycling. In Phosphorus in Action: Soil Biology; Bünemann, E., Oberson, A., Frossard, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; Volume 26. [Google Scholar]
- Harrison, M.J.; van Buuren, M.L. A phosphate transporter from the mycorrhizal fungus Glomus versiforme. Nature 1995, 378, 626–629. [Google Scholar] [CrossRef] [PubMed]
- Tatry, M.V.; Kassis, E.E.; Lambilliotte, R.; Corratgé, C.; Van Aarle, I.; Amenc, L.K.; Alary, R.; Zimmermann, S.; Sentenac, H.; Plassard, C. Two differentially regulated phosphate transporters from the symbiotic fungus Hebeloma cylindrosporum and phosphorus acquisition by ectomycorrhizal Pinus pinaster. Plant J. 2009, 57, 1092–1102. [Google Scholar] [CrossRef] [PubMed]
- Plassard, C.; Becquer, A.; Garcia, K. Phosphorus transport in mycorrhiza: How far are we? Trends Plant Sci. 2019, 24, 794–801. [Google Scholar] [CrossRef]
- Mujica, M.I.; Cisternas, M.; Claro, A.; Simunovic, M.; Pérez, F. Nutrients and fungal identity affect the outcome of symbiotic germination in Bipinnula fimbriata (Orchidaceae). Symbiosis 2020, 83, 91–101. [Google Scholar] [CrossRef]
- Jarosh, K.A.; Kandeler, E.; Frossard, E.; Bünemann, E.K. Is the enzymatic hydrolysis of soil organic phosphorus compounds limited by enzyme or substrate availability? Soil Biol. Biochem. 2019, 139, 107628. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; Bahram, M. Mycorrhizal types differ in ecophysiology and alter plant nutrition and soil processes. Biol. Rev. 2019, 94, 1857–1880. [Google Scholar] [CrossRef]
- Linden, B.R. Comparison of radial growth rate of mycorrhizal fungi isolated from 43 species of northern orchids. Karstenia 1988, 28, 19–25. [Google Scholar] [CrossRef]
- Prosser, J.I. Kinetics of filamentous growth and branching. In The Growing Fungus; Gow, N.A.R., Gadd, G.M., Eds.; Chapman & Hall: London, UK, 1995; pp. 301–318. [Google Scholar]
- Zheng, W.; Lehmann, A.; Ryo, M.; Vályi, K.K.; Rilling, M.C. Growth rate trades off with enzymatic investment in soil filamentous fungi. Sci. Rep. 2020, 10, 11013. [Google Scholar] [CrossRef]
- Ivarsson, M.; Drake, H.; Bengtson, S.; Rasmussen, B. A cryptic alternative for the evolution of hyphae. Bioessays 2020, 42, 1900183. [Google Scholar] [CrossRef] [Green Version]
- Jansa, J.; Mozafar, A.; Frossard, E. Phosphorus acquisition strategies within arbuscular mycorrhizal fungal community of a single field site. Plant Soil 2005, 276, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Bukovská, P.; Rozmoš, M.; Kotianová, M.; Gančarčíková, K.; Dudáš, M.; Hršelová, H.; Jansa, J. Arbuscular mycorrhiza mediates efficient recycling from soil to plants of nitrogen bound in chitin. Front. Microbiol. 2021, 12, 574060. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OTU Code | Fungal Group Code | Orchid Host | Site | Genbank Accession Number | Genetic Similarity with the Nearest Taxonomically Informative Blast |
|---|---|---|---|---|---|
| CER1 | CER | Neotinea ustulata | F | MZ502998 | 96.9% AF354063.1 Thanatephorus cucumeris |
| CER17 | CER | Anacamptis morio | F | MZ502999 | 89.0% AJ427402.1 Ceratobasisium anceps |
| CER19 | CER | Ophrys sphegodes subsp. passionis | F | MZ503000 | 91.2% DQ097889.1 Ceratobasidium sp. |
| SER3 | SER | Neottia ovata | CZ | MZ503001 | 97.1% EU625992.1 Sebacina vermifera |
| SER4 | SER | Neottia ovata | CZ | MZ503002 | 95.8% NR144842.1 Serendipita herbamans |
| TUL1 | TUL-A | Anacamptis pyramidalis | F | MZ503003 | 97.9% AB369932.1 Epulorhiza sp. |
| TUL4 | TUL-A | Orchis mascula | F | MZ503004 | 97.9% MT489316.1 Tulasnella helicospora |
| TUL7 | TUL-B | Anacamptis morio | F | MZ503005 | 96.3% AY373298.1 Tulasnella calospora |
| TUL8 | TUL-B | Anacamptis morio | F | MZ503006 | 76.4% GU166413.1 Tulasnella irregularis |
| Source Type | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Carbon | Nitrogen | Phosphorus | ||||||||
| Media Type | Multiple Comparison Fungal Group vs. Source Type | Estimate (±SE) | z-Value | p-Value | Estimate (±SE) | z-Value | p-Value | Estimate (±SE) | z-Value | p-Value |
| Solid | CER vs. SER | 0.37 (0.17) | 2.10 | 0.15 | 0.26 (0.17) | 1.47 | 0.45 | 0.11 (0.18) | 0.62 | 0.92 |
| CER vs. TUL−A | 0.78 (0.17) | 4.38 | <0.001 *** | 1.01 (0.17) | 5.64 | <0.001 *** | 0.55 (0.18) | 2.98 | 0.01 * | |
| CER vs. TUL−B | 0.14 (0.17) | 0.82 | 0.84 | 0.44 (0.17) | 2.49 | 0.060 | −0.09 (0.18) | −0.49 | 0.96 | |
| SER vs. TUL−A | 0.41 (0.19) | 2.09 | 0.15 | 0.74 (0.19) | 3.80 | <0.001 *** | 0.44 (0.20) | 2.15 | 0.14 | |
| SER vs. TUL−B | −0.22 (0.19) | −1.15 | 0.65 | 0.18 (0.19) | 0.93 | 0.79 | −0.21 (0.20) | −1.02 | 0.74 | |
| TUL−A vs. TUL−B | −0.63 (0.19) | −3.24 | 0.006 ** | −0.56 (0.19) | −2.87 | 0.02 * | −0.65 (0.20) | −3.17 | 0.008 ** | |
| Liquid | CER vs. SER | 0.95 (0.28) | 3.38 | 0.003 ** | 0.88 (0.28) | 3.09 | 0.01 * | 1.38 (0.31) | 4.36 | <0.001 *** |
| CER vs. TUL−A | 1.35 (0.28) | 4.80 | <0.001 *** | 1.36 (0.28) | 4.74 | <0.001 *** | 2.90 (0.32) | 8.95 | <0.001 *** | |
| CER vs. TUL−B | 0.77 (0.28) | 2.74 | 0.030 * | 0.15 (0.28) | 0.54 | 0.95 | 1.13 (0.31) | 3.59 | 0.002 ** | |
| SER vs. TUL−A | 0.39 (0.30) | 1.27 | 0.58 | 0.48 (0.31) | 1.54 | 0.41 | 1.51 (0.35) | 4.30 | <0.001 *** | |
| SER vs. TUL−B | −0.18 (0.30) | −0.59 | 0.93 | −0.72 (0.31) | −2.31 | 0.09 | −0.25 (0.34) | −0.73 | 0.88 | |
| TUL−A vs. TUL−B | −0.57 (0.30) | −1.87 | 0.24 | −1.20 (0.31) | −3.83 | <0.001 *** | −1.76 (0.34) | −5.07 | <0.001 *** | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Novotná, A.; Mennicken, S.; de Paula, C.C.P.; Vogt-Schilb, H.; Kotilínek, M.; Těšitelová, T.; Šmilauer, P.; Jersáková, J. Variability in Nutrient Use by Orchid Mycorrhizal Fungi in Two Medium Types. J. Fungi 2023, 9, 88. https://doi.org/10.3390/jof9010088
Novotná A, Mennicken S, de Paula CCP, Vogt-Schilb H, Kotilínek M, Těšitelová T, Šmilauer P, Jersáková J. Variability in Nutrient Use by Orchid Mycorrhizal Fungi in Two Medium Types. Journal of Fungi. 2023; 9(1):88. https://doi.org/10.3390/jof9010088
Chicago/Turabian StyleNovotná, Alžběta, Sophie Mennicken, Caio C. Pires de Paula, Hélène Vogt-Schilb, Milan Kotilínek, Tamara Těšitelová, Petr Šmilauer, and Jana Jersáková. 2023. "Variability in Nutrient Use by Orchid Mycorrhizal Fungi in Two Medium Types" Journal of Fungi 9, no. 1: 88. https://doi.org/10.3390/jof9010088
APA StyleNovotná, A., Mennicken, S., de Paula, C. C. P., Vogt-Schilb, H., Kotilínek, M., Těšitelová, T., Šmilauer, P., & Jersáková, J. (2023). Variability in Nutrient Use by Orchid Mycorrhizal Fungi in Two Medium Types. Journal of Fungi, 9(1), 88. https://doi.org/10.3390/jof9010088