3.2. Taxonomy
MycoBank: MB847441
Type species—Heterocorticium bambusicola S.H. He, T. Nie & Yue Li
Etymology—“Hetero-”: different, refers to the septate basidia; “Corticium”: a generic name of corticioid homobasidiomycetes, refers to the basidiomes similar to Corticium.
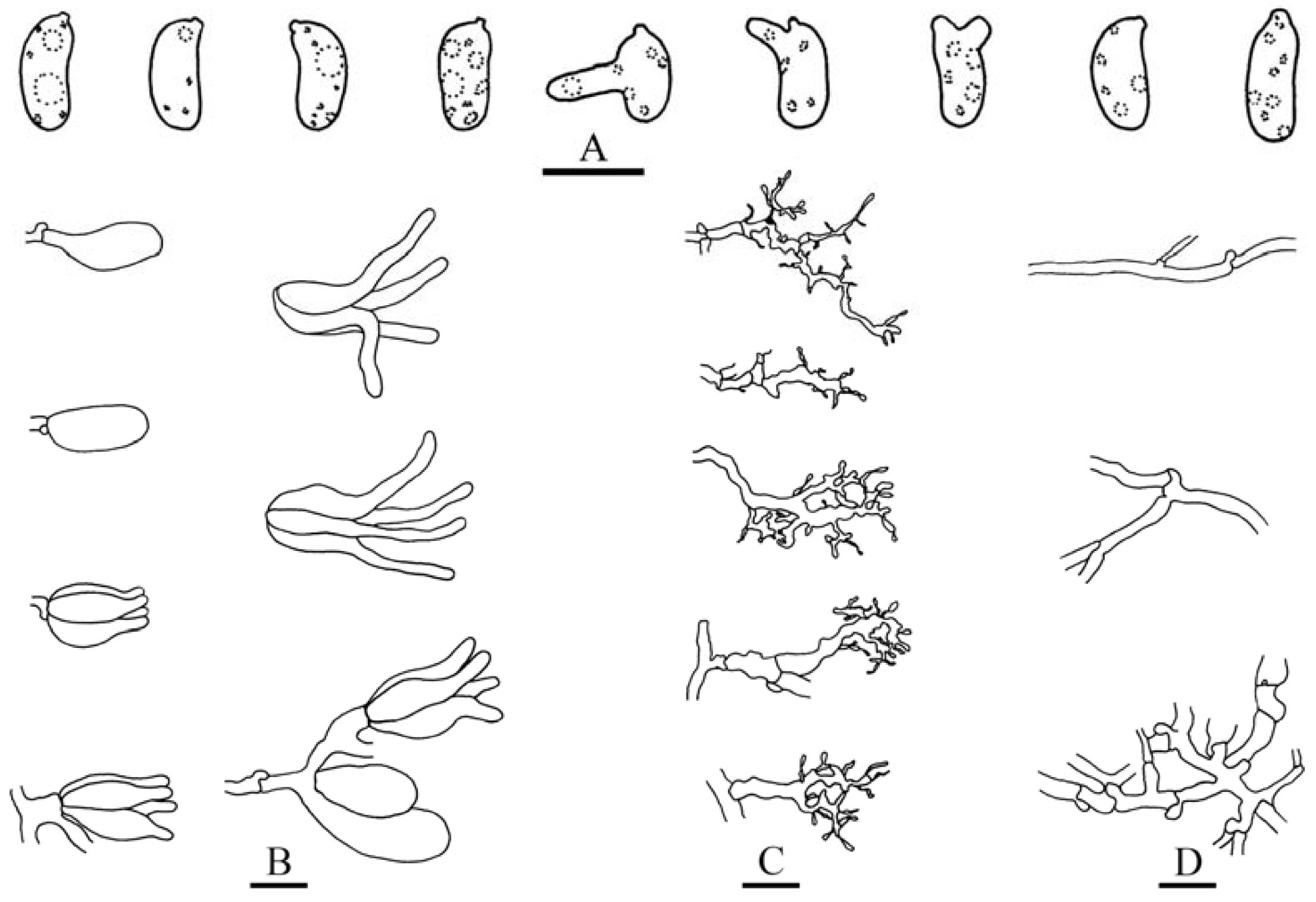
Basidiomes annual, resupinate, effused, closely adnate, inseparable from substrate, thin, coriaceous. Hymenophore smooth, grey, orange to brown; margin thinning out or abrupt, adnate. Hyphal system monomitic; generative hyphae clamped, colorless, thin- to thick-walled, frequently branched, septate. Subiculum indistinct, composed of densely interwoven hyphae. Cystidia absent or present. Dikaryophyses present, colorless, thin-walled, frequently branched. Basidia ovoid or subglobose, longitudinally septate, four-celled, embedded, without enucleate stalk. Basidiospores cylindrical or ellipsoid with an apiculus, colorless, thin-walled, smooth, IKI–, CB–, with oily contents, capable of germinating by repetition.
Notes—
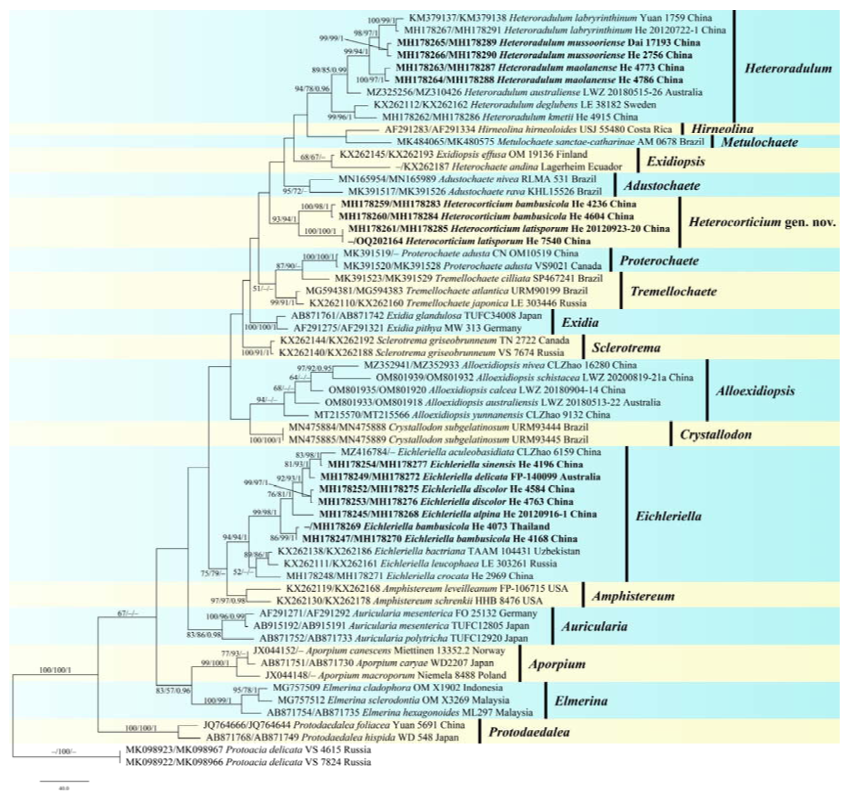
Heterocorticium is characterized by the resupinate coriaceous basidiomes with smooth, pigmented hymenophores, monomitic hyphal system with clamped generative hyphae, and cylindrical or ellipsoid basidiospores. Macroscopically,
Heterocorticium resembles a typical crust, but can be readily distinguished from homobasidiomycetes by its longitudinally septate basidia.
Heterocorticium is similar to
Exidiopsis and
Alloexidiopsis in that they differ by pale-colored basidiomes [
4,
17,
18]. The monotypic genus
Sclerotrema recently segregated from
Exidiopsis is similar to
Heterocorticium by sharing a pigmented hymenophore but differs by its distinctly curved allantoid basidiospores and occurrence mainly on
Alnus [
15,
18]. In the phylogenetic tree,
Heterocorticium formed a distinct lineage and did not show a close relationship with other genera (
Figure 2).
MycoBank: MB847442
Type—China, Jiangxi Province, Yifeng County, Guanshan Nature Reserve, on culm of dead bamboo, 10 August 2016, He 4236 (BJFC 023678, holotype, CFMR, isotype).
Etymology—refers to its preferred substrate, bamboo.
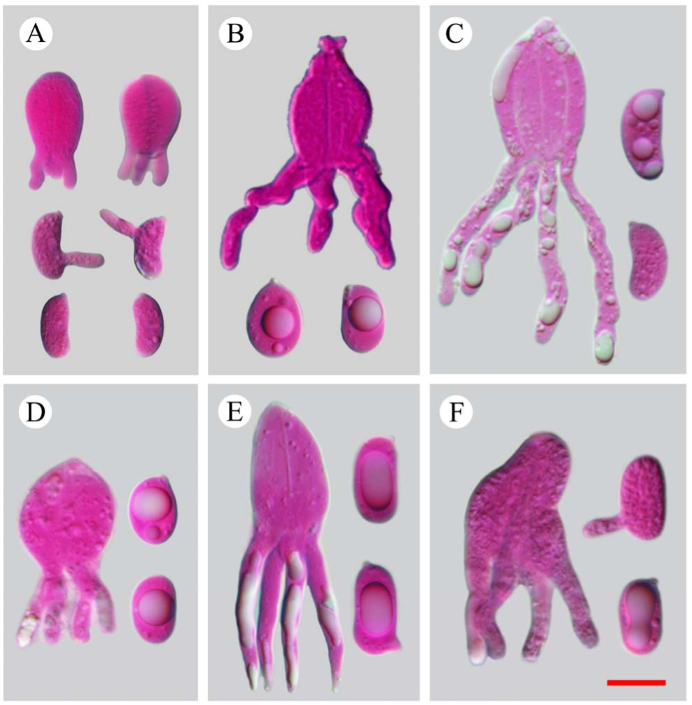
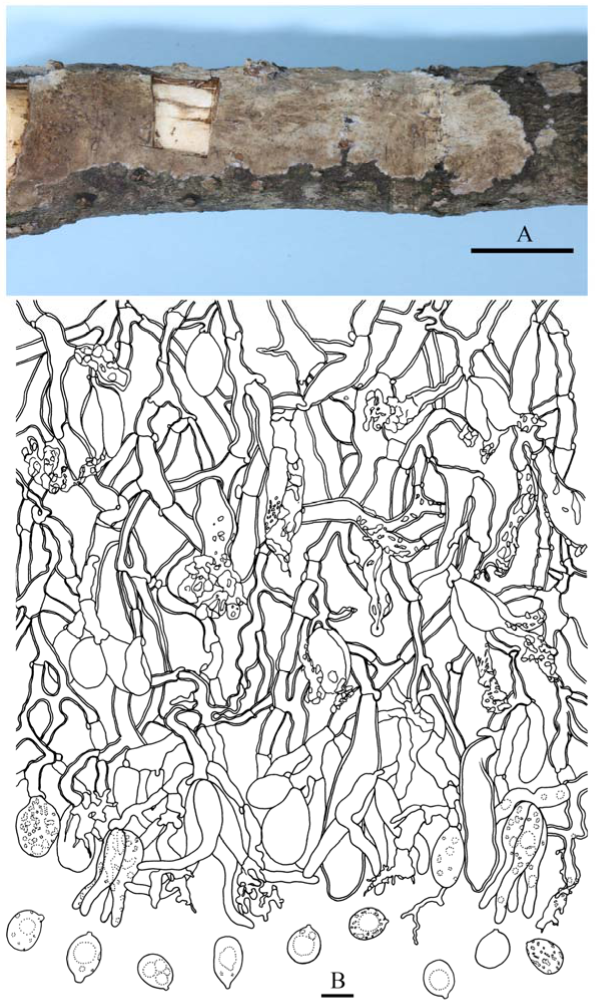
Fruiting body—Basidiomes annual, resupinate, effused, closely adnate, inseparable from substrate, coriaceous, first as small colonies, later confluent up to 20 × 5 cm, up to 120 µm thick in section. Hymenophore smooth, orange white (6A2), orang grey (6B2), greyish orange [6B(3–6)] to brownish orange [6C(3–6)], not cracked; margin thinning out or abrupt, distinct and white (6A1) when juvenile, indistinct and concolorous with hymenophore when mature.
Microscopic structures—Hyphal system monomitic; generative hyphae with clamp connections. Subiculum thin, with many crystals adjacent to substrate; hyphae in this layer colorless, thin-walled, frequently branched and septate, slightly agglutinated, densely interwoven, 1–3 µm in diam. Cystidia absent. Dikaryophyses numerous, forming a light brown layer in hymenium, colorless, thin-walled, frequently branched. Basidia embedded, ovoid to subglobose, longitudinally septate, four-celled, without enucleate stalk, 13–20 × 9–11 µm. Basidiospores cylindrical with a distinct apiculus, slightly curved, colorless, thin-walled, smooth, IKI–, CB–, with oily contents, capable of germinating by repetition, 10–14 × 5–6 µm, L = 11.5 µm, W = 5.5 µm, Q = 1.8–2.4 (n = 60/2).
Additional specimens examined—China, Fujian Province, Jian’ou County, Wanmulin Nature Reserve, on culm of dead bamboo, 19 August 2016, He 4545 (BJFC 023986); Nanping County, Wuyishan Forest Park, on dead bamboo, 3 October 2018, He 5691 (BJFC 026753); Guangxi Autonomous Region, Xing’an County, Mao’ershan Nature Reserve, on culm of dead bamboo, 13 July 2017, He 4799 (BJFC 024318, CFMR); Jiangxi Province, Yifeng County, Guanshan Nature Reserve, on culm of dead bamboo, 10 August 2016, He 4234 (BJFC023676) and He 4260 (BJFC 023702); Anyuan County, Sanbaishan Forest Park, on culm of dead bamboo, 15 August 2016, He 4415 (BJFC 023856); Taiwan, Nantou County, Jenai Township, Entrance of Southern Tungyenshan, alt. 1700 m, on culm of dead bamboo, 7 December 2016, He 4604 (BJFC 024046, CFMR) and He 4607 (BJFC 024049); Xitou, on culm of rotten bamboo, 11 December 2016, He 4638 (BJFC 024081); Yunnan Province, Lushui County, Gaoligongshan Nature Reserve, on culm of dead bamboo, 29 November 2015, He 3341 (BJFC 021736, CFMR), He 3344 (BJFC 021739) and He 3359 (BJFC 021754); Xichou County, Xiaoqiaogou Forest Farm, on culm of dead bamboo, 16 November 2019, He 6293 (BJFC 033237); Malaysia, Kuala Lumpur, Forest Eco-Park, on culm of dead bamboo, 8 December 2019, He 6430 (BJFC 033374) and He 6435 (BJFC 033379).
Notes—Heterocorticium bambusicola is widely distributed in southern China on bamboo and was also found in Malaysia. It is characterized by thin, resupinate, coriaceous basidiomes, smooth hymenophore, and well-developed dikaryophyses. The other species in the genus, H. latisporum, differs from H. bambusicola by having grayish brown hymenophore and wider basidiospores.
Heterocorticium latisporum S.H. He, T. Nie & Yue Li,
sp. nov. Figure 5B and
Figure 8
MycoBank: MB847443
Type—China, Sichuan Province, Qionglai County, Tiantaishan Forest Park, on fallen angiosperm branch, 23 September 2012, He 20120923-20 (BJFC 014667, holotype, CFMR, isotype).
Etymology—refers to wide basidiospores.
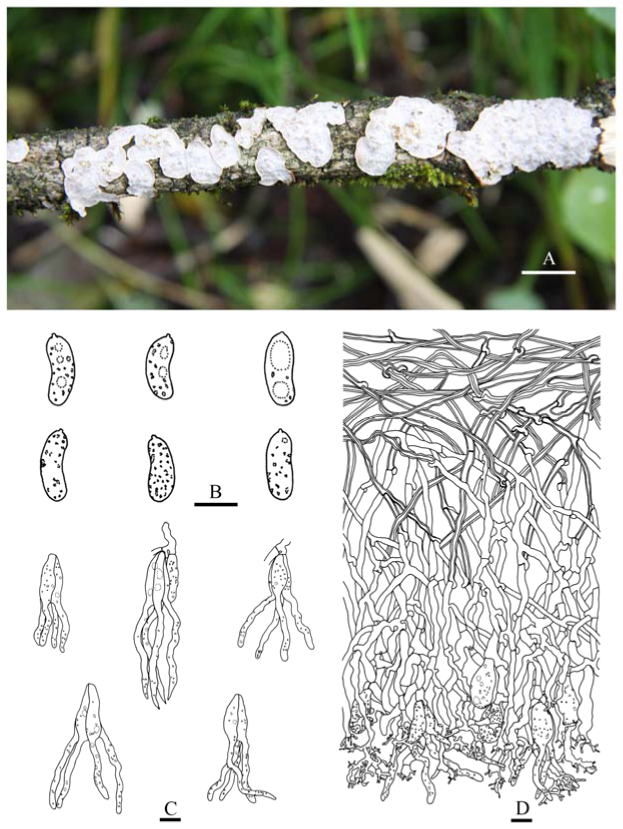
Fruiting body—Basidiomes annual, resupinate, effused, closely adnate, inseparable from substrate, coriaceous; first as small, scattered colonies, later confluent up to 10 × 2.5 cm, up to 150 µm thick in section. Hymenophore smooth, brownish orange [6C(3–4)], greyish brown [6(D–F)3], light brown [6D(4–6)] to brown [6E(4–6)], not cracked or locally cracked; margin thinning out, white (6A1) when juvenile, becoming concolorous with hymenophore when mature.
Microscopic structures—Hyphal system monomitic; generative hyphae with clamp connections. Subiculum thin, with scattered crystals; hyphae colorless, thin- to slightly thick-walled, frequently branched, septate, slightly agglutinated, densely interwoven, 2–3 µm in diam. Cystidia clavate, colorless, with a basal clamp connection, 20–60 × 6–13 µm. Dikaryophyses numerous, colorless, thin-walled, slightly branched. Basidia embedded, ovoid to subglobose, longitudinally septate, four-celled, without enucleate stalk, 16–22 × 11–15 µm. Basidiospores broadly ellipsoid-to-ovoid with a distinct apiculus, colorless, thin-walled, smooth, IKI–, CB–, with oily contents, capable of germinating by repetition, 11–13 (–14) × 8–9 µm, L = 12 µm, W = 8.5 µm, Q = 1.4 (n = 30/1).
Additional specimen examined—China, Anhui Province, Jingde County, Majiaxi Forest Park, on dead angiosperm branch, 27 July 2022, He 7540 (BJFC 038676).
Notes—Heterocorticium latisporum is characterized by resupinate coriaceous basidiomes with a grayish brown, smooth hymenophore, and broadly ellipsoid-to-ovoid basidiospores.
MycoBank: MB847444
Type—China, Sichuan Province, Xiaojin County, Siguniangshan Nature Reserve, on dead angiosperm branch, 16 September 2012, He 20120916-1 (BJFC 014581, holotype).
Etymology—refers to growing on mountains of high altitude.
Fruiting body—Basidiomes annual, discoid, resupinate to slightly effused-reflexed, adnate, separable from substrate, coriaceous, brittle; first as small patches, later confluent up to 5.5 cm long, 1.5 cm wide, up to 400 µm thick in section. Hymenophore smooth to slightly tuberculate, white (6A1) when fresh, becoming greyish green [1D(2–5)], yellowish grey (2C2) to olive grey (2D2) upon drying, not cracked or partly cracked when dry; margin abrupt, slightly elevated, concolorous or slightly darker than hymenophore.
Microscopic structures—Hyphal system monomitic, generative hyphae with clamp connections. Basal layer present, yellow to yellowish brown; hyphae yellow, distinctly thick-walled, densely agglutinated, unbranched, aseptate, 3–5 µm in diam. Subiculum thick; hyphae in this layer colorless, slightly-to-distinctly thick-walled, rarely branched, moderately septate, interwoven, 2–4 µm in diam. Hymenium composed of dikaryophyses, basidia and immature basidia. Cystidia absent. Dikaryophyses numerous, colorless, thin-walled, frequently branched. Basidia clavate to subcylindrical, colorless, longitudinally septate, four-celled, 20–26 × 10–15 µm. Immature basidia abundant, subglobose to ellipsoid. Basidiospores cylindrical with a small apiculus, slightly curved, colorless, thin-walled, smooth, IKI–, CB–, with oily contents, capable of germinating by repetition, 15–17 × 6–7 µm, L = 16 µm, W = 6.5 µm, Q = 2.5 (n = 30/1).
Notes—
Eichleriella alpina is characterized by having discoid-to-slightly effused-reflexed basidiomes, a distinct basal layer, relatively large basidiospores and a distribution in temperate areas of southwestern China.
Eichleriella macrospora (Ellis and Everh.) G.W. Martin is similar but differs in having slightly shorter basidiospores (10–15 × 5–7 µm) and a distribution in the north central USA [
33]. In the phylogenetic tree,
E. alpina formed a distinct lineage in the
Eichlerilla clade (
Figure 3).
MycoBank: MB847445
Type—Thailand, Chiang Rai Province, Doi Mae Salong, on culm of dead bamboo, 22 July 2016, He 4073 (BJFC 023514, holotype, CFMR, isotype).
Etymology—refers to growing on bamboo.
Fruiting body—Basidiomes annual, resupinate, adnate, separable from substrate, soft, membranous to coriaceous, first as small colonies, later confluent up to 10 cm long, 4 cm wide, up to 500 µm thick in section, new basidiomes are usually grown from dead parts. Hymenophore smooth, greyish orange [5B(3–5)] to brownish orange [5C(3–5)] in fertile parts, light brown [5D(3–5)] in sterile or dead areas, not cracked; margin slightly thinning out, white (5A1), fimbriate when juvenile, becoming concolorous or darker than hymenophore with age.
Microscopic structures—Hyphal system dimitic, generative hyphae with clamp connections. Subiculum thick, yellow. Skeletal hyphae setae-like, predominant, yellow to yellowish brown, distinctly thick-walled to subsolid, frequently branched, usually bifurcated, up to 7 µm in diam., slightly dextrinoid, branches short or long with acute tips. Generative hyphae rare to abundant, colorless, thin- to slightly thick-walled, rarely branched, frequently septate, scattered among skeletal hyphae, 1.5–4 µm in diam. Hymenium composed of dikaryophyses, basidia and immature basidia. Cystidia absent. Dikaryophyses numerous, colorless, thin-walled, frequently branched, usually stalked, swollen in middle part. Basidia ovoid to subglobose, colorless, longitudinally septate, four-celled, 15–20 × 9–13 µm. Basidiospores ellipsoid-to-broadly ellipsoid, colorless, thin-walled, smooth, IKI–, CB–, with oily contents, capable of germinating by repetition, 10–13 × 6–7.5 µm, L = 11 µm, W = 7 µm, Q = 1.6–1.7 (n = 60/2).
Additional specimens examined—China, Hunan Province, Dong’an County, Shunhuangshan Nature Reserve, on culm of dead bamoo, 3 July 2015, He 2365 (BJFC 020819, CFMR) and He 2371 (BJFC 020825, CFMR); Jiangxi Province, Yifeng County, Guanshan Nature Reserve, on culm of dead bamboo, 9 August 2016, He 4168 (BJFC 023610, CFMR); Zhejiang Province, Lin’an County, Tianmushan Nature Reserve, on culm of dead Phyllostachys, 15 October 2004, Dai 6391 (BJFC 016628, CFMR); Thailand, Chiang Rai Province, Doi Pui, on culm of dead bamboo, 23 July 2016, He 4088 (BJFC 023529).
Notes—
Eichleriella bambusicola is characterized by having resupinate basidiomes, a dimitic hyphal system with yellow setae-like skeletal hyphae, ellipsoid-to-broadly ellipsoid basidiospores, and fruiting on bamboo in subtropical and tropical areas of China.
Eichleriella bambusicola is similar to
E. leveilleana (Berk. and M.A. Curtis) Burt and
E. schrenkii Burt because of its dimitic hyphal system with richly dichotomously branched and yellow-to-brown skeletal hyphae. This character was mentioned by Malysheva and Spirin [
15] when establishing the genus
Amphistereum for the latter two species. Notably,
E. bambusicola nested within the
Eichleriella s.s. lineage rather than the
Amphistereum lineage in our phylogenetic analyses, although the two lineages are sisters with relatively strong support values in MP and ML analyses (
Figure 2).
Eichleriella tenuicula (Lév.) Spirin and Malysheva also has a dimitic hyphal system but differs from
E. bambusicola by having a spiny hymenophore, unbranched skeletal hyphae, and two-celled basidia [
15,
16]. In the phylogenetic tree, samples of
E. bambusicola from Thailand and China formed a distinct linage (
Figure 3). The yellow, setae-like skeletal hyphae of
E. bambusicola resemble the dichohyphae of
Vararia P. Karst, which is a homobasidiomycetes in the Russulales [
34].
Eichleriella delicata (Bres.) S.H. He & Nakasone,
comb. nov. Figure 11A
MycoBank: MB847446
≡ Heterochaete delicata Bres., Hedwigia 53: 77, 1912.
≡ Hydnum delicatum Klotzch ex Berk., Ann. Nat. Hist., Mag. Zool. Bot. Geol. 3 no. 19: 395, 1839 (nom. illeg.); non H. delicatum Schwein., Trans. Am. Phil. Soc., New Series 4(2): 161. 1832.
Specimens studied—Eichleriella delicata: Australia, New South Wales, Sydney, Hyde Park, on hardwood, 15 October 2001, H.H. Burdsall, Jr., FP-140099 (CFMR); China, Yunnan Province, Ruili County, Moli Tropical Rain Forest Park, on fallen angiosperm branch, 2 December 2015, He 3469 (BJFC 021866, CFMR).
Eichleriella tenuicula: China, Hainan Province, Danzhou Tropical Botanical Garden, on fallen angiosperm branch, 7 May 2009, Cui 6306 (BJFC 004162); Taiwan, Nantou County, Xinyi Township, Xitou, on fallen angiosperm branch, 11 December 2016, He 4634 (BJFC 024077); Yunnan Province, Ruili County, Moli Tropical Rain Forest Park, on fallen angiosperm branch, 2 December 2015, He 3483 (BJFC 021880, CFMR); Thailand, Chiang Rai Province, Campus of Mae Fah Luang University, on dead but still attached branch of Cinnamomum porrectum, 21 July 2016, He 4053 (BJFC 023492).
Notes—Bodman [
16] treated
Heterochaete tenuicula (Lév.) Pat. as a synonym of
H. delicata, whereas Roberts [
35] preferred
H. tenuicula and treated
H. delicata as a synonym. Malysheva and Spirin [
15] transferred
H. tenuicula to
Eichleriella but did not mention
H. delicata.
Heterochaete delicata was described from India whereas
H. tenuicula was from Java, Indonesia. Morphologically, the two species are similar sharing effused-to-effuse-reflexed basidiomes, two-celled basidia, and long basidiospores [
16,
35]. However, the taxa are placed in distinct lineages in the phylogenetic tree (
Figure 3). Although inseparable in morphology, we accept
H. delicata and
H. tenuicula as distinct species and propose the transfer of
H. delicata to
Eichleriella. Significantly, the ITS sequence similarity between
H. delicata (He 3469) and
H. tenuicula (He 3483) is 97.5 % of 514 base pairs.
Eichleriella discolor (Berk. & Broome) S.H. He & Nakasone,
comb. nov. Figure 11B
MycoBank: MB847447
≡ Kneiffia discolor Berk. & Broome, Journal Linnean Society. Botany 14: 62, 1873 (1875).
≡ Kneiffiella discolor (Berk. & Broome) Henn., in Engler & Prantl, Nat. Pflanzenfam.: I, Teil. 1 (Leipzig), Abt.: Fungi (Eumycetes): 141, 1898.
≡ Heterochaete discolor (Berk. & Broome) Petch, Annals Royal Botanic Gardens Peradeniya 9: 137, 1924.
= Heterochaete tonkiniana Pat., Bulletin Herbier Boissier 1: 301, 1893.
= Heterochaete roseola Pat., Bulletin Société Mycologique France 29: 207, 1913.
= Heterochaete cheesmanii Wakef., Bulletin Miscellaneous Information, Kew 1915: 373.
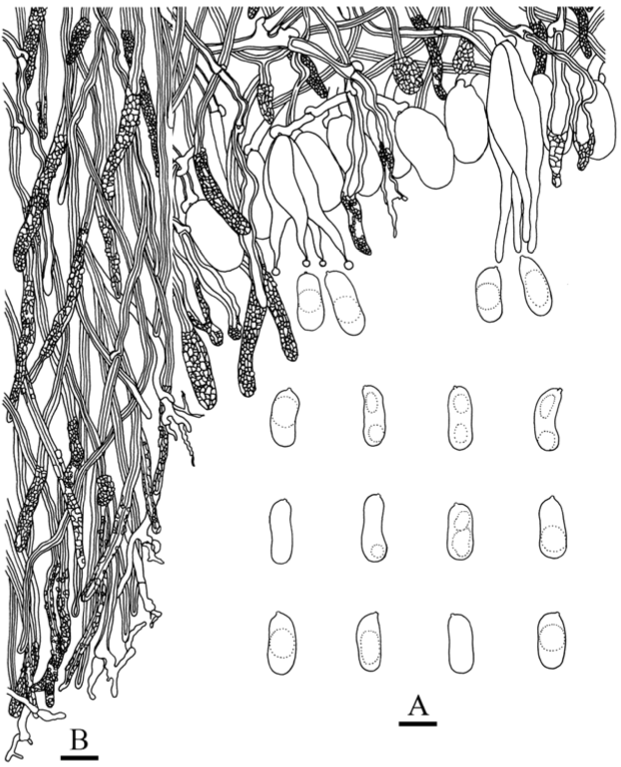
Fruiting body—Basidiomes widely effused, closely adnate; first as small circular-to-orbicular colonies, later confluent, up to 7 cm long, 4 cm wide, thin, 100–300 (–900) μm thick, soft, subceraceous to cretaceous. Hymenophore spiny-to-velutinous from hyphal pegs, (4–) 6–8 (–10) per mm, with smooth areas between pegs; pegs above hymenium (70–) 85–200 (–240) × (30–) 45–60 (–80) μm, single, terete, brittle, colorless to concolorous first then becoming dark yellow or light brown; overall surface first yellowish white (4A2), pale yellow (4A3), orange white (5A2), pale orange (5A3), light orange (5A4), then darkening to greyish orange (5B3), brownish orange (5C5), yellowish brown (5D4); margins variable sometimes within a single collection, adnate to somewhat detached, distinct, abrupt, thick, raised, felty-silky, brownish yellow, usually darker than hymenophore, or appressed, adnate, thinning out with edges white, fimbriate, silky to woolly.
Microscopic structures—Hyphal system monomitic but appearing dimitic from sclerified hyphae in subiculum and pegs. Hyphal pegs arising deep in subiculum as a compact, agglutinated fascicle composed of an inner core of brown pigmented, sclerified subicular hyphae, then above the hymenium hyphae differentiating into dikaryophyses along sides and at apices, often encrusted, sometimes terminal dikaryophyses colorless and thin-walled throughout or yellow to brownish yellow and walls thickening toward base. Subiculum up to 150 (–700) μm thick, a partially agglutinated tissue composed of a dense, dark yellow-to-brown tissue of sclerified hyphae arranged parallel to substrate, giving rise to hyphal pegs, hyphae 2–4 μm diam, clamps inconspicuous or degraded, sparsely branched, walls yellow to brownish yellow, thin to 1 μm thick, smooth. Subhymenium up to 80 μm thick, a moderately dense, colorless tissue of loosely interwoven hyphae turning into hymenium, composed of hyphae, 2.5–3.5 μm in diameter, clamped, walls thin, colorless, smooth. Hymenium 25–100 μm thick, a dense palisade of dikaryophyses enclosing cystidia and basidia. Dikaryophyses abundant, 35–60 × 3.5–5 μm, clamped at base, simple or with short nodulose or knobby branches, walls colorless, thin, smooth. Cystidia scattered to numerous, embedded to barely projecting, cylindrical, clavate, or subfusiform, often with a stalk, apices rounded, subacute or bulbous, 40–70 × 7–13 μm, clamped at base, walls colorless, thin, smooth, with homogenous contents. Basidia longitudinally septate, four-celled, embedded with only tips of sterigmata observed beyond hymenium, ellipsoid to ovoid, (11–) 13–23 × 8.5–11 μm, with a small, basal clamp, 4-sterigmate, rarely with 2 or 3 sterigmata, walls colorless, thin, smooth. Basidiospores cylindrical, often ventrally depressed, (10–) 11–13.5 (–15) × 5–6 μm, L = 12.5 μm, W= 5.5 μm, Q = 2.3–2.4 (60/2), colorless, thin-walled, smooth, IKI–, CB–, germination sometimes observed.
Distribution—Australia, Cambodia, China, Nepal, New Zealand, Philippines, Sri Lanka, Thailand, Vietnam.
Type specimens examined—Australia, New South Wales, Moruya, on bare wood, W.N. Cheesman, 1914 (K(M)4481, holotype of H. cheesmanii); Sri Lanka, Central Province, on wood December 1868, G.H.K.Thwaites 982 (K(M)132234, holotype of Kneiffia discolor); Vietnam, Hanoi, on bark, 22 June 1911, Dupont no. 676 (FH–HUH00940160, holotype of H. roseola); Vietnam, Tonkin, Thanh Hoa Ngoc Au (Than Hoa), on bark and wood, 29 January 1892, H.F. Bon 5062 (FH-HUH01093670, holotype of H. tonkiniana).
Representative specimens examined—Australia, Victoria, Gippsland Highland, Tarra Valley, on fallen branches of orangewood, 12 Sept 1955, K. Healy, N.W.M. Walters, & E. DaCosta (MEL-2313649, as H. discolor); Cambodia, Reserv foretrie de Campong Chhnang, on (bark of) dead branches, July 1921, P.A. Pételot 360 (FH-HUH00940161; BPI-719711, as H. roseola); China, Gansu Province, Tianshui County, Maijishan Forest Park, on fallen angiosperm branch, 8 August 2015, He 2488 (BJFC 020941); Guangxi Autonomous Region, Tianlin County, Cenwanglaoshan Nature Reserve, on dead angiosperm branch, 8 July 2017, He 4708 (BJFC 024227, CFMR); Huanjiang County, Mulun Nature Reserve, on rotten angiosperm trunk, 10 July 2017, He 4763 (BJFC 024281); Guizhou Province, Libo County, Maolan Nature Reserve, on dead angiosperm branch, 14 June 2016, He 3771 (BJFC 022270); 11 July 2017, He 4778 (BJFC 024295); Hainan Province, Wanning County, Xinglong Botanical Garden, on fallen angiosperm trunk, 19 March 2016, He 3610 (BJFC 022108); Wuzhishan County, Wuzhishan Nature Reserve, on dead angiosperm branch, 10 June 2016, He 3947 (BJFC 022449); Hunan Province, Zhangjiajie County, Zhangjiajie Nature Reserve, on fallen angiosperm tree, 7 July 2015, He 2297 (BJFC 020752); Yizhang County, Mangshan Nature Reserve, on dead angiosperm branch, 26 July 2016, He 4008a (BJFC 023446); Taiwan, Nantou County, Jenai Township, Entrance of Southern Tungyenshan, alt. 1700 m, on fallen angiosperm trunk, 7 December 2016, He 4584 (BJFC 024026); Xizang Autonomous Region, Linzhi County, Lulang, on dead angiosperm trunk, 17 September 2010, He 321 (BJFC 021980, CFMR); on dead Hippophae branch, 17 September 2010, He 331 (BJFC 021982); Yunnan Province, Jingdong County, Ailaoshan Nature Reserve, on dead angiosperm branch, 25 August 2015, He 2620 (BJFC 021066, CFMR); Baoshan County, Gaoligongshan Nature Reserve, Baihualing, on dead Quercus branch, He 3401 (BJFC 021797); Nepal, Chitwan National Park, Narayani River, Island Jungle Resort, on hardwood twig, 15 March 2008, T. Rödel, NP/13 (Rödel private herbarium, CFMR); New Zealand, Weraroa, Wellington, on fallen log, 30 October 1919, G.H. Cunningham 353 (BPI-719737, as H. tenuicula); Philippines, Luzon, Mt. Maquiling, on dead wood, 28 Oct 1920, F. Obias, ex Herb. O.A. Reinking no. 10516 (BPI-719764, FH-HUH00940152, as H. leveillei); Thaliand, Chiang Rai Province, Doi Mae Salong, on dead angiosperm branch, 22 July 2016, He4069 (BJFC 023510); Chiang Mai Province, Doi Saket, on dead angiosperm branch, 24 July 2016, He 4107 (BJFC 023549); Vietnam, Thac Mai Preservation Park, lowland rain forest Dipterocarp, on dead angiosperm branch, 14 October 2017, He 5211 (BJFC 024729); Tonkin, Cho-Ganh, sur branchettes de Flamboyant, 13 Mar 1914, L. Duport no. 14 (FH-HUH00940159, as H. roseola); sur “Dong” (bark of twigs), Janvier 1922, M. Duport no. 127 (FH-HUH00940157, as H. roseola) and M. Duport no. 132 pp (FH-HUH01093669, as H. tonkiniana).
Notes—
Eichleriella discolor is characterized by subceraceous-to-chalky or cretaceous basidiomes, small hyphal pegs arising from subiculum and composed of brownish yellow, sclerified hyphae and dikaryophyses, and 4-sterigmate basidia. The margin of specimens can be quite variable, and hyphal pegs are brittle, often breaking off to expose its dark brown interior. The size of the basidiospores is also variable and may reflect the number of sterigmata produced by the basidia. Its hyphal system was described as pseudodimitic [
36], because the hyphae in the subiculum and hyphal peg are brownish yellow with slightly thickened walls. On close inspection, however, rare clamp connections were observed on these sclerified hyphae.
Eichleriella discolor is widely distributed from East and Southeast Asia to New Zealand and has been described several times. The synonymy presented above was reached after studying type specimens and mostly supports previous reports except that the synonymy of
H. roseola with
K. discolor is new. Bodman [
16] considered
K. discolor and
H. tonkiniana to be synonyms but recognized
H. roseola as a distinct species. Earlier, Roberts and Spooner [
36] proposed the synonymy of
K. discolor and
H. cheesmanii. Eichleriella sinensis (Teng) S.H. He & Nakasone,
comb. nov. Figure 11C,D
MycoBank: MB847448
≡ Heterochaete sinensis Teng, Sinensia 7: 530, 1935.
= Eichleriella aculeobasidiata Hui Wang, Dong-Qiong Wang & C.L. Zhao, Kew Bull. 77: 326, 2022.
Fruiting body—Basidiomes widely effused, resupinate, first as small orbicular colonies, later confluent, adnate, moderately thick, (150–) 200–450 (–500) μm thick, firm, subceraceous to ceraceous. Hymenophore distinctly odontoid from hyphal pegs, (4–) 5–8 (–10) per mm, with smooth areas among pegs; pegs single, conical, terete, concolorous to brown, brittle, breaking off to reveal dark brown trama overall surface yellowish white (4A2), greyish yellow (4B3), pale orange (6A3), greyish orange (6B4), brownish orange (6C4), light brown [6D(4–5)] to dark brown (7F8), black in KOH then fading on drying; margins distinct, abrupt, detached, sometimes incurved, pale orange (5B3), light orange (5A4), greyish orange [5B(4-5)], often darker than hymenophore, with sterile edges up to 1 mm wide, even to scalloped, felty-woolly, rarely fimbriate.
Microscopic structures—Hyphal system dimitic with clamps on generative hyphae and thick-walled skeletal hyphae. Hyphal pegs arising from upper subiculum, 80–140 (–160) × (30–) 40–50 (–80) μm, a tight fascicle of non-agglutinated skeletal hyphae and coarse, colorless crystals, along sides and at apex skeletal hyphae differentiating into branched, thin-walled dikaryophyses. Subiculum up to 400 μm thick, composed of two distinct layers; (1) a non-agglutinated tissue, up to 200 μm thick, of dense, non-agglutinated, skeletal hyphae and degraded generative hyphae arranged parallel to substrate; (2) an interwoven tissue of non-agglutinated primarily skeletal hyphae and degraded subicular hyphae turning into hymenium, up to 250 μm thick, often with embedded, coarse colorless crystals; subicular hyphae mostly collapsed, 2–3 μm diam, clamped, walls colorless, thin; skeletal hyphae 2–4.5 μm diam, aseptate, sometimes with adventitious septa, rarely branched, walls pale yellow to brownish yellow, up to 2 μm thick, smooth. Subhymenium not observed. Hymenium up to 80 μm thick, a dense palisade of dikaryophyses enclosing cystidia and basidia. Dikaryophyses abundant, 30–60 × 3–4.5 μm, clamped at base, with short, knobby branches, walls colorless, thin, smooth. Cystidia numerous, embedded, cylindrical, clavate, or subfusiform, often stalked, apices rounded or subacute, 15–50 (–70) × 7.5–9 (–12.5) μm, clamped at base, walls colorless, thin, smooth, with homogenous contents. Basidia embedded, ovoid to ellipsoid, longitudinally septate, four-celled, 15–20 (–25) × 9–11 (–12.5) μm, with a small, basal clamp, 4-sterigmate, walls colorless, thin, smooth. Basidiospores scarce-to-numerous, cylindrical-to-narrowly cylindrical, sometimes ventrally depressed, (9.5–) 10.5–16 (–18) × (4.5–) 5.5–7 μm, L = 15 μm, W = 6.5 μm, Q = 2.2–2.4 (60/2), walls colorless, thin, smooth, IKI–, CB–, germination sometimes observed.
Distribution—Pantropical and subtropical: Australia, China, Indonesia, Japan, New Zealand, Philippines.
Specimens examined—Australia, Queensland, Cook, Innisfail, Joint Tropical Research Unit, on fallen, rotten twigs, 5 September 1972, N.E.M. Walters (MEL-231548, with E. discolor); China, Anhui Province, Chiu-hua-shan, (on small twigs), 18 September 1933, S.C.Teng 698 (paratype, BPI-719773); Fujian Province, Wuyishan County, Wuyishan Nature Reserve, Huanggangshan, on fallen angiosperm branch, 21 October 2005, Dai 7361 (BJFC 016640); Jian’ou County, Wanmulin Nature Reserve, on dead angiosperm branch, 19 August 2016, He 4547 (BJFC 023988); Guangxi Autonomous Region: Jinxiu County, Dayaoshan Nature Reserve, Shengtang Mountains, on rotten angiosperm trunk, 15 July 2017, He 4862 (BJFC 024381); Yinshan Park, on dead angiosperm branch, 16 July 2017, He 4900 (BJFC 024419); Guizhou Province, Suiyang County, Kuankuoshui Nature Reserve, on fallen angiosperm trunk, 26 November 2014, Dai 15044 (BJFC 018157); Libo County, Maolan Nature Reserve, on dead angiospermic branch, 14 June 2016, He 3761 (BJFC 022260, CFMR); Hainan Province, Baisha County, Yinggeling Nature Reserve, on dead angiosperm branch, 9 June 2016, He 3890 (BJFC 022392) and He 3892 (BJFC 022394); Henan Province, Xinyang County, Jigongshan Nature Reserve, on fallen Fagus branch, 27 October 2014, He 20141027-4 (BJFC 019266); Hubei Province, Wufeng County, on dead angiosperm branch, 14 August 2017, He 5025 (BJFC 024543); 15 August 2017, He 5057 (BJFC 024575); Hunan Province, Heng-shan, 22 September 1933, C.I. Shen 559 (paratype, BPI-719724); Dong’an County, Shunhangshan Nature Reserve, on dead angiospermic branch, 13 July 2015, He 2388 (BJFC 020842, CFMR); Liuyang County, Daweishan Forest Park, on dead angiosperm branch, 10 July 2015, He 2330 (BJFC 020784); Jilin Province: Jiaohe County, on dead angiosperm branch, 3 Sep. 2017, He 5145 (BJFC 024663). Jiangxi Province, Yifeng County, Guanshan Nature Reserve, on fallen angiosperm trunk, 9 August 2016, He 4185 (BJFC 023627); on dead angiosperm branch, 9 August 2016, He 4196 (BJFC 023638); Anyuan County, Sanbaishan Forest Park, on dead angiosperm branch, 15 August 2016, He 4425 (BJFC 023866); Sichuan Province, Ya’an, Bifengxia Forest Park, on bark of living Cunninghamia, 22 September 2012, He 20120922-4 (BJFC 014644); Qionglai County, Tiantaishan Forest Park, on dead angiosperm tree, 23 September 2012, He 20120923-18 (BJFC 014665); Taiwan, Nantou County, Jenai Township, Entrance of Southern Tungyenshan, alt. 1700 m, on dead angiosperm branch, 7 December 2016, He 4598 (BJFC 024040); Yunnan Province, Xichou County, Lotus pond, on fallen angiosperm branch, 25 July 2014, He 20140725-12 (BJFC 019206); Binchuan County, Jizhushan Forest Park, on dead angiosperm branch, 28 October 2017, He 5327 (BJFC 024845); Zhejiang Province, Tunglu, on dead twigs, 26 November 1933, S.Q. Deng 699 (paratype, BPI-719723; CUP-CH-000942); Indonesia, West Java, 1921, C. Hartley (BPI-719743, as H. tenuicula); Japan, no location data, K. Aoshima 10751 (NY-00461157, as H. tenuicula); New Zealand, Auckland, Mt. Albert Rd., on Tecoma sp., 6 November 1948, G.L.S. Chamberlain (PDD-24331, as H. tenuicula); Philippines, Quizon, Bataan, Lamae, on dead twigs, 30 November 1960, F.R. Vyenco (NY00461200, as H. delicata); Diliman, U.P. Campus, on bark, 7 August 1959, F.R. Vyenco (NY-00461151, as H. delicata); Los Banos, Luzon, Mt. Maquiling, on Cryptocarya dead branches, 30 July 1918, O.A. Reinking 7587 (BPI-719745, as H. tenuicula).
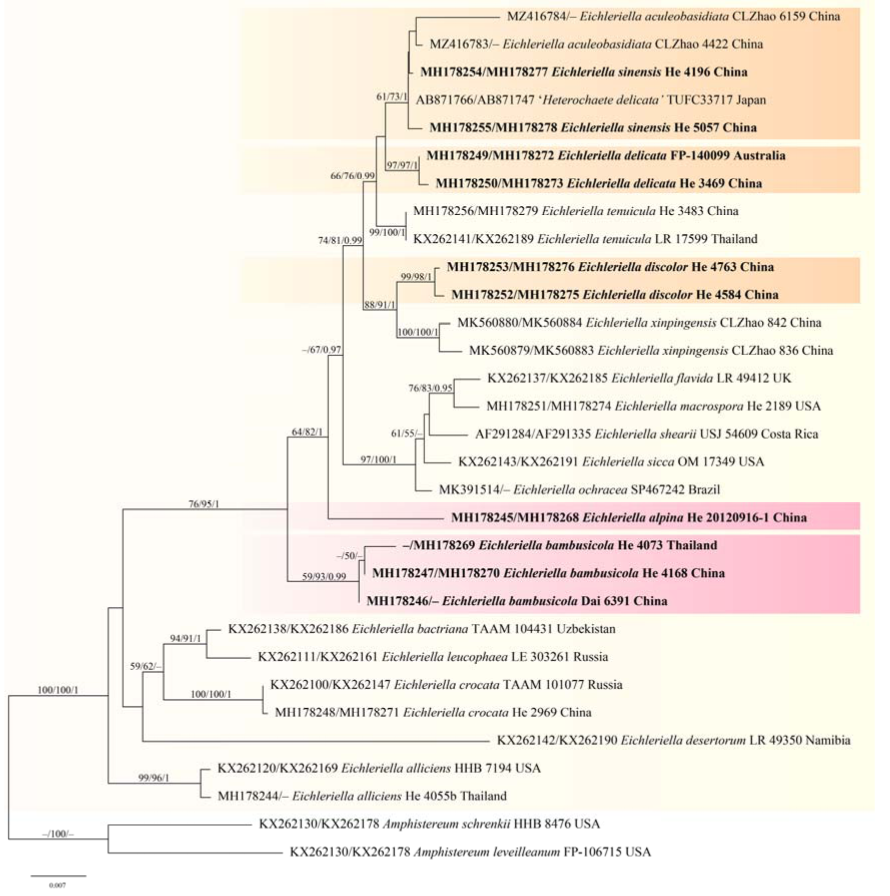
Notes—Eichleriella sinensis is similar to E. tenuicula except for the 4-sterigmate basidia and cylindrical basidiospores, average Q values 2.2–2.4, compared with allantoid basidiospores, average Q value 3.3 of the latter. The two taxa also overlap in distribution in Asia, Australia, New Zealand, and the Philippines but E. tenuicula has a pantropical distribution. Basidiospores were scarce in most of the specimens examined. Although the type specimen of H. sinensis was not examined, three paratypes were studied.
In the phylogenetic tree of
Eichleriella (
Figure 3),
E. sinensis,
E. tenuicula and
E. delicata are closely related but formed distinct lineages. The recently described species,
E. aculeobasidiata Hui Wang, Dong Qiong Wang & C.L. Zhao, has 2-sterigmate basidia but nested within the
E. sinensis lineage [
28]. Close inspection of basidia of the type specimens of
E. aculeobasidiata needs to be carried out, but at present we accept
E. aculeobasidiata as a synonym of
E. sinensis based on phylogenetic analyses and overall morphology. A sample from Japan identified as
Heterochaete delicata (TUFC33717) in GenBank also nested within the
E. sinensis lineage instead of the
E. delicata lineage in our phylogenetic tree (
Figure 3).
MycoBank: MB847449
Type—China, Guizhou Province, Libo County, Maolan Nature Reserve, on dead but still attached angiosperm branch, 11 July 2017, He 4773 (BJFC 024290, holotype, CFMR, isotype).
Etymology—refers to the type locality in Maolan Nature Reserve, Guizhou Province, southwestern China.
Fruiting body—Basidiomes annual, resupinate, effused, closely adnate, inseparable from substrate, coriaceous, first as small, scattered colonies, later confluent up to 10 cm long, 2.5 cm wide, up to 600 µm thick in section (hyphal pegs excluded). Hymenophore odontoid, irpicoid to subporoid from hyphal pegs, light orange [5A(4–5)], greyish orange [5B(4–6)] to brownish orange [5C(4–6)], not cracked; hyphal pegs densely arranged, single or fused, with blunt tips; margin abrupt, sterile, white (6A1) when juvenile, becoming concolorous with hymenophore with age.
Microscopic structures—Hyphal system monomitic-to-subdimitic, generative hyphae with clamp connections. Subiculum thick, pale yellow; hyphae colorless to pale yellow, slightly to distinctly thick-walled, rarely branched, septate, densely interwoven, smooth or encrusted with small granules, 2–3.5 µm in diam. Hyphal pegs originated from base of hymenium, pale yellow, up to 240 × 160 µm; hyphae similar to those of subiculum, agglutinated, vertically arranged, with tips heavily encrusted with crystals. Cystidia absent. Dikaryophyses numerous, colorless, thin-walled, encrusted, not branched or slightly branched at the tip. Basidia ovoid to subglobose, longitudinally septate, four-celled, without enucleate stalk, 18–25 × 10–15 µm. Basidiospores cylindrical with an apiculus, sometimes slightly curved, colorless, thin-walled, smooth, IKI–, CB–, containing one or two globules, capable of germinating by repetition, 12–15 × 6–7.5 µm, L = 16.5 µm, W = 7 µm, Q = 1.9–2 (n = 90/3).
Additional specimens examined—China, Guizhou Province, Libo County, Maolan Nature Reserve, on bark of dead angiosperm tree, 11 July 2017, He 4786 (BJFC 024303, CFMR); on dead but still attached angiosperm branch, 15 June 2016, He 3788 (BJFC 022287, CFMR).
Notes—
Heteroradulum maolanense is characterized by its odontioid, irpicoid-to-subporoid hymenophore with single or fused hyphal pegs, encrusted hyphae and cylindrical basidiospores. The species is similar to
H. mussooriense which differs in having wider ellipsoid-to-ovoid basidiospores (12–15 × 9–10 µm, [
16]).
Heteroradulum australiense L.W. Zhou, Q.Z. Li & S.L. Liu like
H. maolanense has an odontoid hymenophore but differs in having both simple- and nodose-septate generative hyphae and longer basidiospores (15–20 × 5–7 µm, [
20]). In the phylogenetic tree, the two samples of
H. maolanense formed a distinct, strongly supported (
Figure 4).
MycoBank: MB847450
≡ Heterochaete mussooriensis Bodman, Lloydia 15: 221, 1952.
= Heteroradulum semis Spirin & Malysheva, Fungal Biology 121: 712, 2017.
≡ Grammatus semis (Spirin & Malysheva) H.S. Yuan & Decock, MycoKeys 35: 35, 2018.
Specimens studied—China, Hubei Province, Shenlongjia Nature Reserve, on fallen angiosperm branch, 16 October 2016, Dai 17193 (BJFC 023291, CFMR); Yunnan Province, Binchuan County, Jizhushan Forest Park, on fallen angiosperm branch, 30 August 2015, He 2867 (BJFC 021301, CFMR); 26 November 2015, He 3168 (BJFC 021563, CFMR); 28 October 2017, He 5306 (BJFC 024824, CFMR) and He 5331 (BJFC 024849); Yongde County, Daxueshan Nature Reserve, on Quercus stump, 28 August 2015, He 2756 (BJFC 021194).
Notes—
Heterochaete mussooriensis was described from Mussoorie, northwestern India, and then also reported in China [
16,
37]. The species is characterized by its subporoid hymenophore formed by numerous densely arranged spines, large subclavate basidia and broadly ellipsoid-to-ovoid basidiospores [
16]. Malysheva and Spirin [
15] described
Heteroradulum semis from northeastern China and Japan. However, the protologue and our phylogenetic analyses show that
H. semis is conspecific with
Heteroradulum mussooriensis.
≡ Grammatus labyrinthinus H.S. Yuan & Decock, MycoKeys 35: 32, 2018.
Specimens examined—China, Guangxi Autonomous Region, Longzhou County, Nonggang Nature Reserve, on fallen angiosperm branch, 22 July 2012, He 20120722-1 (BJFC 014502, CFMR); Guizhou Province, Chishui County, Suoluo Nature Reserve, on fallen angiosperm branch, 7 July 2018, He 5431 (BJFC 026492) and He 5439 (BJFC026500).
Notes—Yuan et al. [
30] established the genus,
Grammatus H.S. Yuan & Decock, based on
G. labyrinthinus and proposed the combination
G. semis. The genus was later regarded as a synonym of
Heteroradulum [
20] and is also supported by our phylogenetic analyses (
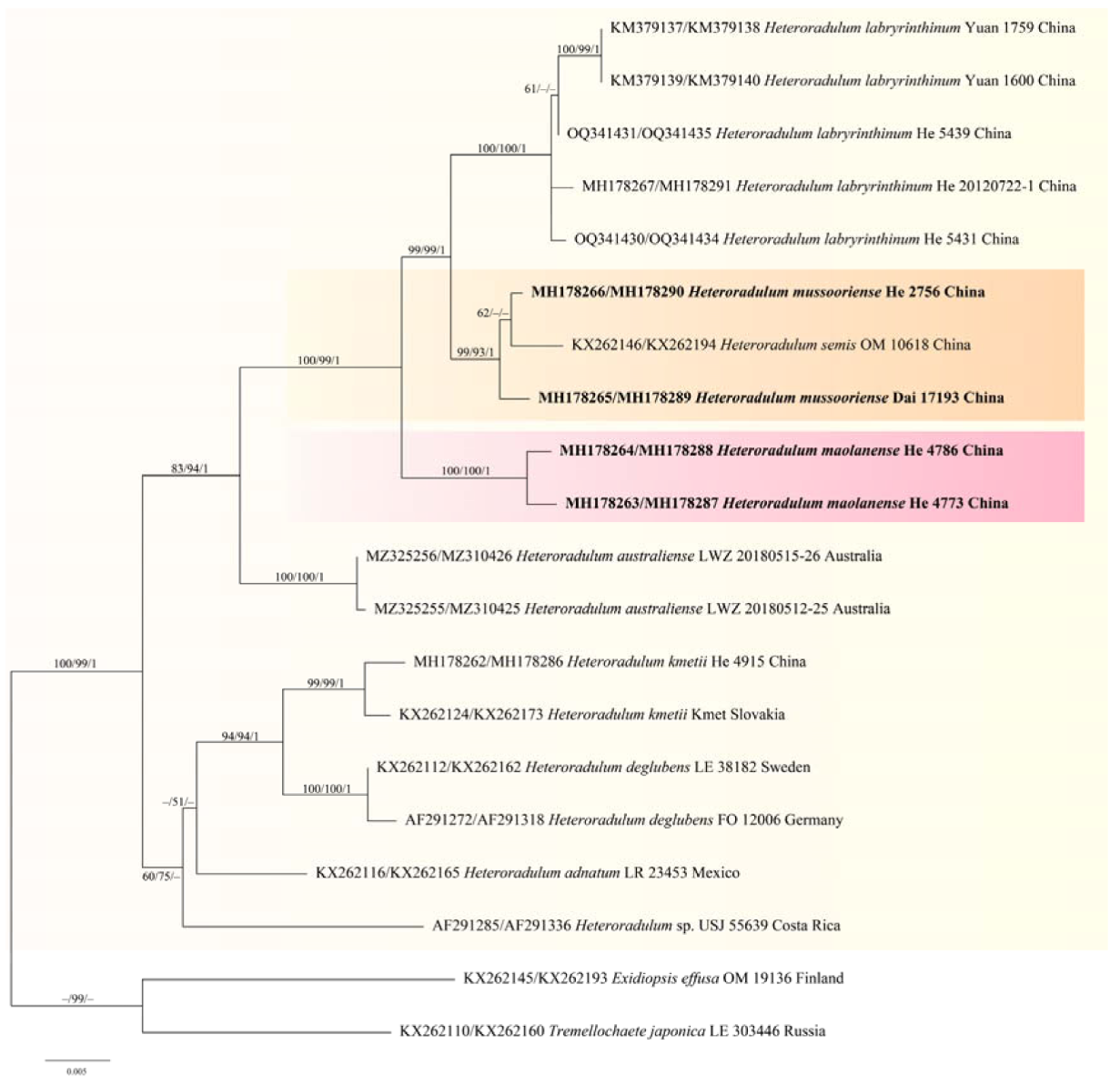
Figure 2). In the phylogenetic tree of
Heteroradulum (
Figure 4), three species,
H. labyrinthinum,
H. mussooriense and
H. maolanense, form a strongly supported clade. Morphologically, the three species have similar and varied hymenophore that can be odontioid, irpicoid to subporoid because of the confluence the hyphal pegs at different growth stages (
Figure 12).
Heteroradulum labyrinthinum was originally described from tropical areas of Yunnan Province; herein, we report it from Guanxi Autonomous Region and Guizhou Province.

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}